8. Нервная ткань 1-2 лекции.ppt
- Количество слайдов: 118



Морфофункциональные особенности НЕРВНАЯ ТКАНЬ

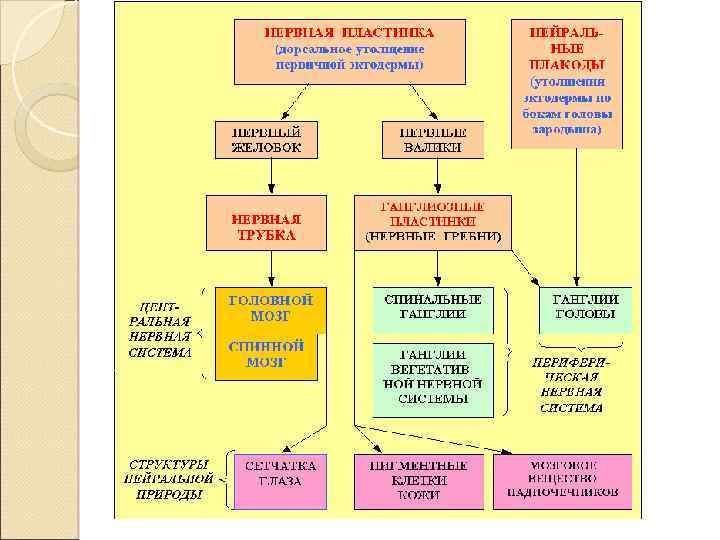
Развитие В срединной части эктодермы появляется – нервная пластинка. Под влиянием индукторов, выделяемых хордой, пластинка начинает впячиваться, что приводит к образованию нервного желобка и нервных валиков.

Развитие Затем желобок смыкается в нервную трубку , стенка которой представляет собой многорядный нейроэпителий, Нервные валики превращаются в парные нервные гребни, или ганглиозные пластинки – рыхлые скопления клеток между нервной трубкой и эктодермой.

Образование нервной трубки

Нервная ткань нейроны эфферентн ые афферентн ые нейроглия ассоциати вные макроглия микроглия астроциты эпендимоц иты олигоденд роциты

Строение нейрона Нейроны содержат: Ядро, преобладает эухроматин; Хорошо развитую гр- ЭПС, КГ - сетчатый аппарат Гольджи, Митохондрии, Лизосомы и т. д. Интенсивно синтезируются РНК и белки.
, Отростки. Длина отростков варьирует – от нескольких микрометров до 1–")
Строение нейрона Тело (перикарион), Отростки. Длина отростков варьирует – от нескольких микрометров до 1– 1, 5 м. По своей функциональной роли отростки подразделяются на 2 типа: дендриты – проводят импульсы к телу нейрона аксон, или нейрит - всегда один и проводит импульсы от тела нейрона.
 2 – аксональный холмик 3 –")
Строение нейрона 1 – хроматофильная субстанция (гр. ЭПС) 2 – аксональный холмик 3 – аксон 4 – ядро 5 - ядрышко
 поступающие сигналы, переходить в ответ на сигналы в состояние")
Функции нейрона Способны рецептировать (воспринимать) поступающие сигналы, переходить в ответ на сигналы в состояние возбуждения или торможения, проводить возбуждение или торможение (например, от конца дендрита – к перикариону и затем от последнего – к концу аксона), передавать сигнал другим объектам – очередному нейрону или эффекторному органу.
,")
Функциональная классификация нейронов По функции нейроциты делятся на 3 основных вида: чувствительные (или рецепторные), ассоциативные эффекторные. нейросекреторные нейроны.

Нейрофибриллы в теле и во всех отростках выявляются нейрофибриллы. в теле они образуют плотную сеть, в отростках идут вдоль оси. Нейрофибриллы представлены пучками нейротрубочек и нейрофиламентов. они формируют трёхмерную сеть – важнейшую структуру цитоскелета.

Строение нейрона

Нейроны коры головного мозга

Морфологическая классификация По общему количеству отростков нейроны и их предшественники делятся на 4 типа: Униполярные (нейробласты), Псевдоуниполярные, Биполярные Мультиполярные нейроны.

Классификация нейронов

Способность нейронов к возбуждению и торможению связана с наличием в их плазмолемме систем транспорта ионов: Na+, K+-насосов, К+-каналов Na+-каналов.
: снаружи клеток имеется некоторый")
в покоящейся клетке создаётся трансмембранная разность потенциалов (трансмембранный потенциал): снаружи клеток имеется некоторый избыток положительных зарядов, а внутри – избыток отрицательных зарядов.

При возбуждении открываются Na+-каналы: ионы Na+ устремляются в клетку по градиенту концентрации; Поэтому трансмембранная разность потенциалов снижается – происходит деполяризация плазмолеммы. По окончании возбуждения Na+-каналы закрываются и восстанавливается исходное значение потенциала (реполяризация).

При торможении ситуация обратная: открытие Na+-каналов затрудняется, это понижает чувствительность клетки к возбуждающим сигналам.

Направление транспорта веществ Отростки нейронов служат не только для проведения сигналов. По ним постоянно происходит транспорт веществ. Различают два направления транспорта: прямое (антероградное) – перемещение веществ от перикариона к периферии отростка ретроградное – перемещение в обратном направлении, к перикариону.
 по аксонам в прямом направлении - со скоростью 1")
Виды транспорта медленный ток (транспорт) по аксонам в прямом направлении - со скоростью 1 -3 мм/сутки; быстрый ток по аксонам в прямом направлении - 100 -1000 мм/сутки; ток по дендритам в прямом направлении - 75 мм/сутки; ретроградный ток по аксонам и дендритам.
; Кислород")
Транспорт веществ от тела клетки Метаболиты (образование медиаторов и энергетическое обеспечение данного процесса); Кислород (для окисления в митохондриях); Белки (в т. ч. ферменты), Нейрогормоны (в аксонах нейросекреторных клеток) и др. вещества;

Транспорт веществ к телу клетки - конечные продукты обмена. Многие перечисленные вещества переносятся в растворённой форме, другие же вещества (например, гормоны и медиаторы) - в составе пузырьков или гранул.
")
Механизм транспорта Быстрый транспорт растворённых веществ осуществляется путём тока жидкости (под действием гидродинамического давления) через межтубулярное пространство. Пузырьки же и гранулы транспортируются с помощью двух белков, использующих энергию АТФ: кинезин обеспечивает транспорт в прямом, а динеин – в ретроградном направлении.

Классификация Нейроглия Макроглия Микроглия

Функции нейроглии Обеспечивают деятельность нейронов, играя вспомогательную роль – Опорную, Трофическую, Электроизоляционную, Барьерную Защитную. Секреторная (ликвор),

Макроглия и микроглия Макроглия ЦНС - происходит из глиобластов: олигодендроглия, астроглия, эпендимная глия; микроглия ЦНС- происходит из промоноцитов.
, нейролеммоциты (шванновские клетки).")
Макроглия Глия ПНС: мантийные глиоциты (клетки-сателлиты, или глиоциты ганглиев), нейролеммоциты (шванновские клетки).

Астроциты Имеют многочисленные отростки, идущие во все стороны, что придаёт звёздчатую форму, чем и обусловлено их название (от греч. astron – звезда). На концах многие отростки астроцитов имеют пластинчатые расширения. Толщина и длина отростков зависит от типа астроглии. По этому признаку последнюю подразделяют на 2 вида: протоплазматическую волокнистую.

Функции астроцитов Опорная и барьерная – образуют в мозгу поддерживающую сеть и глиальные пограничные мембраны между капиллярами и нейронами Элемент гематоэнцефалического барьера. Транспортная и трофическая - содержат системы транспорта определённых веществ – в нейроны и, видимо, из нейронов;

Функции астроцитов Регуляторная - выделяют факторы роста нейроцитов – в период развития мозга и при регенерации нервной ткани Обменная - участвуют в обмене медиаторов.
 образует эпендиму – ткань, которая выстилает спинномозговой канал и желудочки мозга, покрывает")
Эпендимоциты (эпендимоглиоциты) образует эпендиму – ткань, которая выстилает спинномозговой канал и желудочки мозга, покрывает сосудистые сплетения желудочков.

Функции эпендимоцитов Продукция и перемещение ликвора – особенно, эпендима, покрывающая сосудистые сплетения желудочков. С помощью же ресничек приводят ликвор в движение, препятствуя его застою в какомлибо желудочке или канале. Барьерные свойства - эпендимоциты, покрывающие сосудистые сплетения желудочков, соединены плотными контактами, создаёт гематоэнцефалический барьер (гематоликворный барьер).

Функции эпендимоцитов Транспортная функция - передача одних веществ из гипоталамуса в гипофиз и других веществ – в обратном направлении.

Эпендимоциты Астроциты
![Олигодендроциты Небольшие глиальные клетки, у которых отростки - немногочисленные (от корня oligo ["мало"] происходит](https://present5.com/presentation/3/-72216801_303138682.pdf-img/-72216801_303138682.pdf-37.jpg "Олигодендроциты Небольшие глиальные клетки, у которых отростки - немногочисленные (от корня oligo [\"мало\"] происходит")
Олигодендроциты Небольшие глиальные клетки, у которых отростки - немногочисленные (от корня oligo ["мало"] происходит название клеток), короткие и слабоветвящиеся. Два вида: Клетки-сателлиты – окружают тела нейронов (мантийные глиоциты) Глиоциты нервных волокон – окружают отростки (леммоциты или шванновские клетки)

Функции олигодендроцитов Трофическую, Барьерную Электроизоляционную. Глиоциты контролируют обмен веществ между нейронами и окружающей средой.

Олигодендроглиоциты

Микроглия Мелкие клетки с продолговатым ядром и с небольшим числом отростков. Встречаются и в сером, и в белом веществе ЦНС. Образуются из промоноцитов, Способны к фагоцитозу и выполняют роль глиальных макрофагов. Считают, что у больных СПИДом микроглиоциты (благодаря своей высокой подвижности) разносят вирус по ЦНС.

Три типа микроглии Амёбоидную - способны к амёбоидным движениям, активно фагоцитируют – например, фрагменты разрушающихся клеток, превращаются в следующий тип микроглии. Ветвистую (покоящуюся) - содержатся в сформированном мозгу, имеют ветвящиеся отростки, не фагоцитируют. Реактивную - образуется из покоящейся микроглии после травмы мозга, отличается высокой фагоцитарной активностью.

Микроглия

Нервные волокна Отросток нейрона вместе с оболочкой называется нервным волокном. Отросток нейрона, находящийся в составе волокна, называется осевым цилиндром. В качестве осевых цилиндров могут выступать дендриты чувствительных нейронов, аксоны эффекторных, а в случае вегетативной нервной системы - и ассоциативных нейронов.

Классификация нервных волокон По своему строению нервные волокна подразделяются на 2 типа – Безмиелиновые (безмякотные) Миелиновые (мякотные).

Локализация безмиелиновых волокон Безмиелиновые волокна находятся: преимущественно - в составе ВНС, содержат аксоны эффекторных нейронов этой системы; в меньшей степени - в ЦНС.
. По периферии в цитоплазму погружено")
Строение безмиелинового волокна В центре располагается ядро олигодендроцита (леммоцита). По периферии в цитоплазму погружено обычно несколько (10 -20) осевых цилиндров. Причём плазмолемма леммоцита смыкается почти над каждым цилиндром, так что образуются дупликатуры плазмолеммы - мезаксоны. С поверхности нервное волокно покрыто базальной мембраной.
 -")
Резюме Таким образом, оболочка осевых цилиндров в безмиелиновых волокнах включает: плазмолемму олигодендроцита (леммоцита) - в составе мезаксона, узкий слой цитоплазмы той же клетки, базальную мембрану.

Схема строения Безмиелиновые волокна

Безмиелиновые нервные волокна

Миелиновые волокна Миелиновые нервные волокна образуют: белое вещество ЦНС, афферентные и эфферентные пути соматической части периферической нервной системы, часть путей вегетативной нервной системы. Миелиновые волокна содержат как аксоны, так и дендриты нервных клеток.

Строение миелиновых волокон Осевой цилиндр в волокне - всего один, располагается в центре и гораздо больше по диаметру, чем в безмиелиновом волокне. Оболочка волокна имеет два слоя: внутренний - миелиновый слой наружный - нейролемму (или неврилемму).
, концентрически закрученными вокруг")
Строение миелинового волокна Миелиновый слой представлен несколькими слоями мембраны олигодендроцита (леммоцита), концентрически закрученными вокруг осевого цилиндра. Фактически это очень удлинённый мезаксон, образующийся при погружении осевого цилиндра в цитоплазму глиоцита и последующем многократном вращении цилиндра вокруг своей оси.
 цитоплазма")
Нейролемма - это оттеснённые к периферии (т. е. кнаружи от миелинового слоя) цитоплазма и ядро глиоцита. Снаружи волокно в периферическом нерве покрыто базальной мембраной.

Миелиновые волокна Главная особенность миелиновых волокон состоит в том, что вокруг осевого цилиндра находится много слоёв плазмолеммы, плотно прижатых друг к другу. Химический состав плазмолеммы – очень специфичен: в них значительно преобладают липиды ( 80% массы). Миелиновые оболочки являются электроизоляторами.

Миелиновый слой волокна регулярно прерывается : Это места стыка соседних леммоцитов: здесь у волокна остаётся только истончённая нейролемма. Такие участки называются узловыми перехватами Ранвье.

Схема строения Миелиновые волокна

Миелиновые нервные волокна

В этих перехватах сосредоточены Na+- каналы осевого цилиндра; а в тех участках цилиндра, которые покрыты миелиновой оболочкой, каналов нет. Такое расположение Na+-каналов значительно увеличивает скорость проведения возбуждения

Передача импульса Импульс передаётся не путём открытия-закрытия Na+-каналов, а путём распространения изменений электрического поля (возникающих в области перехватов). Эти изменения распространяются в хорошо изолированном проводнике (каковым является осевой цилиндр под миелиновым слоем) почти мгновенно.
 механизм передачи нервного")
Механизм передачи нервного импульса В итоге реализуется т. н. сальтаторный (скачкообразный) механизм передачи нервного импульса Он включает 2 чередующихся процесса: сравнительно медленное проведение возбуждения (в виде волны деполяризации) в очередном перехвате Ранвье и очень быструю передачу сигнала в миелинизированном фрагменте волокна до следующего перехвата.

Насечки миелина Помимо перехватов Ранвье, в миелиновом слое обнаруживаются также узкие, косо расположенные, просветления – т. н. насечки миелина. В этих местах концентрические листки мезаксона не так плотно прилегают друг к другу, отчего между ними сохраняются прослойки цитоплазмы. В миелиновых волокнах ЦНС таких насечек нет.

Миелиновые и безмиелиновые нервные волокна

Безмиелиновые Несколько осевых цилиндров, располагающихся по периферии волокна. Осевые цилиндры - аксоны эффекторных нейронов вегетативной нервной системы. Ядра леммоцитов (олигодендроцитов) находятся в центре волокон. Мезаксоны осевых цилиндров – короткие. Na+-каналы располагаются по всей длине осевого цилиндра. Скорость передачи сигнала 1 - 2 м/с Миелиновые Один осевой цилиндр, находящийся в центре волокна. Это может быть и аксон, и дендрит нейроцита. Ядра и цитоплазма леммоцитов (олигодендроцитов) оттеснены к периферии волокна, образуя неврилемму. Мезаксон многократно закручивается вокруг осевого цилиндра, образуя миелиновый слой. Na+-каналы – только в перехватах Ранвье. 5 - 120 м/c

Нервная ткань НЕРВНЫЕ ОКОНЧАНИЯ

Нервные окончания Двигательные – моторная бляшка Рецепторные. Экстерорецепторы Инкапсулированные Неинкапсулированны е Синапсы Электрические Химические (медиаторы) Аксо-аксональные Аксо-соматоческие Аксо-дендрические
 нейронов. Ими заканчиваются эфферентные пути. Вместе с мембраной")
Эффекторные Это окончания аксонов эффекторных (двигательных) нейронов. Ими заканчиваются эфферентные пути. Вместе с мембраной эффекторных клеток (или волокон) они образуют нейроэффекторные синапсы (моторные бляшки).
 - Это окончания дендритов чувствительных нервов. Ими начинаются")
Рецепторные окончания Рецепторные (чувствительные, или афферентные) - Это окончания дендритов чувствительных нервов. Ими начинаются афферентные нервные пути.

Синапсы Основные типы межнейронных синапсы таковы: аксодендритические; аксосоматические; аксональные. Обнаружены также соматодендритические синапсы (между телом одного и дендритом другого нейрона).

Синапсы Аксовазальные синапсы - это окончания аксонов нейросекреторных нейронов на капиллярах.

Рецепторные По происхождению воспринимаемых сигналов: Экстерорецепторы - воспринимают сигналы из внешней среды, интерорецепторы - настроены на сигналы из внутренней среды.
, механо-, баро-, хемо-, термопроприо- (мышечно-суставное")
Рецепторные нервные окончания По природе воспринимаемых сигналов: ноци- (болевые), механо-, баро-, хемо-, термопроприо- (мышечно-суставное чувство) и пр. рецепторы.
; Несвободные")
Классификация По строению рецепторов: Свободные нервные окончания (конечные ветвления осевого цилиндра лишены оболочки); Несвободные нервные окончания (вокруг осевого цилиндра сохраняются клетки глии) неинкапсулированные, инкапсулированные (заключены в соединительнотканную капсулу).

Рецепторы температуры и боли Рецепторы, воспринимающие температуру и боль, просто проникают между клетками эпителия. Механизм действия: при действии раздражителей в рецепторном окончании меняется состояние ионных каналов (причём, не столько натриевых, сколько кальциевых), и это инициирует волну возбуждения в дендрите.

Тактильные рецепторы эпидермиса воспринимают очень слабое прикосновение. Окончания дендрита контактируют с основаниями осязательных эпителиоцитов – клеток Меркеля , которые находятся в базальном слое эпидермиса и часто объединяются в т. н. диски Меркеля. Данный рецептор отнесён к свободным рецепторным окончаниям

Несвободные неинкапсулированные Содержатся в дерме, в больших количествах. Это ветвления дендритов чувствительных нейронов, сопровождаемые обычно изменёнными леммоцитами. Данные рецепторы (как и свободные рецепторы эпидермиса) воспринимают боль и температуру.

Несвободные инкапсулированные окончания Содержат 3 элемента: терминали дендрита, видоизменённые глиальные клетки, окружающие эти терминали; наружную соединительнотканную оболочку.

Виды рецепторов Существует несколько разновидностей инкапсулированных рецепторов: осязательные тельца Мейснера (не путать с осязательными дисками Меркеля!), пластинчатые тельца Фатера. Пачини, тельца Руффини, колбы Краузе и др.

Локализация Все перечисленные рецепторы встречаются в дерме, а тельца Фатера-Пачини, кроме того, содержатся в строме внутренних органов.
 слое дермы. Тельце")
Осязательные тельца Мейснера располагаются сразу под эпидермисом - в поверхностном (сосочковом) слое дермы. Тельце включает 3 компонента: окончания дендрита, окружающие их глиоциты, тонкую капсулу из волокнистой соединительной ткани.

Тельца Мейснера Глиоциты тельца Мейснера называются тактильными клетками. Они передают деформационные сдвиги капсулы тельца на окончания дендритов. Благодаря своему поверхностному положению, тельца Мейснера воспринимают слабое давление, и обеспечивают осязание.

Осязательное тельце Мейснера

Пластинчатые тельца Фатера-Пачини Находятся в глубоком - сетчатом - слое дермы, в строме внутренних органов и в брыжейке. Имеются три компонента: Терминали дендрита (всегда одного), Глиальные клетки образуют внутреннюю колбу (или внутреннюю луковицу) Капсулу – имеет пластинчатую структуру, которая повышает чувствительность рецептора к механическим воздействиям, причём, не только к статическому давлению и вибрации.

Тельце Фатер-Пачини

Тельца Руффини Располагаются в сетчатом слое дермы. В этих тельцах окончания дендритов связаны с коллагеновыми волокнами. Тельца реагируют на растяжение кожи. Некоторые авторы приписывают им также температурную чувствительность.

Колбы Краузе Встречаются, в основном, в коже наружных половых органов и являются механорецепторами. Название обусловлено тем, что окончания дендритов очень сильно ветвятся, образуя сферообразную гроздь, отчего окружающая эту “гроздь” капсула имеет вид колбы.

Нервно-мышечные и нервносухожильные веретена Афферентная импульсация, идущая в ЦНС от мышц и сухожилий, рассматривается как проприоцептивная чувствительность. Она обеспечивается специальными рецепторными структурами, которые тоже являются инкапсулированными. Это нервно -мышечные веретёна, реагирующие на растяжение мышцы, а также нервно-сухожильные веретёна, реагирующие на сокращение мышцы.
 содержит 4 компонента:")
Нервно-мышечные веретена располагаются в толще скелетных мышц. Каждое веретено (fusus) содержит 4 компонента:

Нервно-мышечное веретено 1 - от 1 до 12 мышечных волокон, 2 -соединит. капсулу вокруг веретена, 3 -афферентные нервные волокна и их окончания, 4 -эфферентные нервные волокна - с окончаниями

Различают 2 вида интрафузальных мышечных волокон. Волокна с ядерной сумкой Таких волокон - 1 -3 в веретене; ядра сосредоточены в центральной расширенной части (ядерной сумке) такого волокнa
, ядра")
Волокна с ядерной цепочкой Этих волокон - больше (3 -7 в веретене), ядра расположены цепочкой по длине волокнa, сами волoкна тоньше и короче, чем предыдущие.
, образуют окончания в виде спирали -")
Первичные волокна - относительно толстые (17 мкм), образуют окончания в виде спирали - т. н. кольцеспиральные окончания, оплетают оба вида интрафузальных мышечных волокон, и при растяжении их центральной части реагируют - на скорость и - на общую величину растяжения.
, образуют гроздьевидные окончания, оплетают только волокна")
Вторичные волокна – относительно тонкие (8 мкм), образуют гроздьевидные окончания, оплетают только волокна с ядерной цепочкой и реагируют лишь на величину растяжения центральной части этих волокон.
 растяжении мышцы (например, при резком движении) афферентная ипмульсация")
Таким образом, при внезапном (быстром) растяжении мышцы (например, при резком движении) афферентная ипмульсация возникает только в первичных нервных волокнах, а при медленном и длительном растяжении (например, при стоянии в неудобном положении) - в нервных волокнах обоих видов.

Синапсы Синапс - структура, предназначенная для передачи сигнала с нервной клетки на другую нервную клетку или на эффекторный орган. В синапсе различают 3 основных компонента: пресинаптическую часть синаптическую щель постсинаптическую часть.

Схема строения Классификация Синапсы

Способ передачи импульса У человека и высших животных синапсы относятся к химическому типу: сигнал передаётся с помощью химического вещества - медиатора, который диффундирует в синаптической щели от пресинаптической части к постсинаптической. При этом сигнал может передавать только в одном направлении,
,")
Пресинаптическая часть пресинаптическое часть – это окончание аксона нейрона. содержит пресинаптические пузырьки (с медиатором), В плазмолемме пресинаптического окончания находятся Са 2+-каналы, закрытые в состоянии покоя.

Когда происходит возбуждение, Са 2+- каналы открываются, внутри окончания повышается концентрация ионов Са 2+. Пузырьки выделяются в щель путем экзоцитоза.

Синаптическая щель содержит филаменты, скрепляющие пре- и постсинаптические клетки. Ширина щели – 20 -30 нм. Это расстояние медиатор преодолевает путём диффузии.
, которая находится под")
Постсинаптическая мембрана Это та часть плазмолеммы постсинаптической клетки (или мышечного волокна), которая находится под пресинаптическим окончанием. Содержатся 3 группы специфических белков: рецепторы к медиатору, белки эффекторного или трансмиттерного устройства ферменты, разрушающие медиатор (медиаторы).
 устройства – это ионные каналы, которые открываются при связывании медиатора")
Белки эффекторного (трансмиссионного) устройства – это ионные каналы, которые открываются при связывании медиатора с белками-рецепторами. В результате каналы открываются, что приводит к либо к возбуждению постсинаптической клетки, либо к её торможению.

Холинергические синапсы Медиатором в холинергических синапсах является ацетилхолин. Данные синапсы по природе белкарецептора делятся на две группы: с м-холинорецепторами и с н-холинорецепторами.
 реализуется ионотропный механизм: в них рецепторы связаны с")
В синапсах второй группы (с нхолинорецепторами) реализуется ионотропный механизм: в них рецепторы связаны с катионными каналами, которые открываются под действием ацетилхолина. После открытия катионных каналов внутрь постсинаптической клетки поступают (по градиенту своей концентрации) ионы Na+. Поэтому развивается деполяризация, а значит возбуждение постсинаптической мембраны.
. ГАМК-рецепторы подразделяются на несколько типов: ГАМК-рецепторы типа")
ГАМК-ергические синапсы Медиатором служит ГАМК (гаммааминомасляная кислота). ГАМК-рецепторы подразделяются на несколько типов: ГАМК-рецепторы типа А - ионотропные: но связаны они не с катионными, а с анионными каналами (точнее, каналами для ионов Сl-), которые открываются под действием ГАМК - по градиенту концентрации. Это вызывает гиперполяризацию постсинаптической мембраны и означает торможение соответствующей клетки.

В результате открываются анионные каналы, Открытие каналов приводит к току внутрь постсинаптической клетки отрицательных ионов Это вызывает гиперполяризацию постсинаптической мембраны и торможение соответствующей клетки.

Глицинергические синапсы В них медиатором служит аминокислота глицин, рецептор к нему тоже связан с анионными каналами, и результатом действия глицина на эти рецепторы является торможение постсинаптической клетки.

Возбуждающие и тормозные синапсы Аксодендритические и аксосоматические синапсы могут быть как возбуждающего, так и тормозного типа, Аксоаксональные синапсы бывают только тормозного типа. Спектр медиаторов весьма широк: ацетилхолин, серотонин, норадреналин, ГАМК, дофамин, глицин и многие другие.
 норадреналин, – как правило, тормозные")
ГАМК, дофамин, глицин и (в межнейронных синапсах ЦНС) норадреналин, – как правило, тормозные медиаторы, Ацетилхолин и серотонин – возбуждающие. Нейроны, в зависимости от вида синапса, образуемого их аксоном, тоже делятся на возбуждающие и тормозные.
 передаются на эффекторные органы - мышцы и")
Эффекторные нервные окончания Сигналы (приходящие по аксону) передаются на эффекторные органы - мышцы и железы. Эффекторные нервные окончания в скелетных мышцах называются нервно -мышечными окончаниями - моторными пластинками, или моторными бляшками.

В данных окончаниях медиатор - ацетилхолин, рецепторы - нхолинорецепторы. Являются н-холинергическими.
")
Моторная бляшка (двигательное нервное окончание)

Пресинаптическое часть Являются тела эффекторных нейронов, иннервирующих скелетные мышцы, расположены в двигательных ядрах спинного и головного мозга.

Строение моторной бляшки Подходя к мышечному волокну, аксон теряет оболочку, образуемую леммоцитами, и разветвляется на несколько терминальных ветвей, которые погружаются в мышечное волокно вместе с прогибающейся сарколеммой и образуют пресинаптические окончания, которые отделены от сарколеммы синаптической щелью. Плазмолемма такого окончания служит пресинаптической мембраной синапса.

Строение моторной бляшки В пресинаптических окончаниях содержится много митохондрий и пузырьков с ацетилхолином. Для синтеза медиатора используется фермент ацетилхолинсинтетаза. В плазмолемме окончаний имеются, как уже отмечалось, Са 2+-каналы, которые открываются при возбуждении

Механизм передачи импульса В последующие события, приводящие к экзоцитозу, вовлечён ещё целый ряд специфических белков: протеинкиназа и синаптогамин, активируемые ионами Са 2+; синапсин – в покое связывает пузырьки с цитоскелетом, а под действием указанной протеинкиназы теряет такую способность; синаптопорин – под влиянием синаптогамина связывает пузырьки с пресинаптической мембраной, формируя в них общие поры и инициируя слияние мембран, т. е. экзоцитоз.

Постсинаптическая мембрана В качестве постсинаптической мембраны выступают участки сарколеммы, которые прогнулись вглубь мышечного волокна, окружая пресинаптические окончания аксона. Постсинаптическая мембрана имеет многочисленные инвагинации, значительно увеличивающие её площадь.
, соединённые с ними")
В мембране содержатся три ключевых белка: рецепторы к ацетилхолину (нхолинорецепторы), соединённые с ними катионные каналы, открывающиеся при связывании ацетилхолина с холинорецепторами, и фермент холинэстераза, разрушающий ацетилхолин.

СПАСИБО ЗА ВНИМАНИЕ!
8. Нервная ткань 1-2 лекции.ppt