

Молекул основы наследств.ppt
- Количество слайдов: 48
 Молекулярные основы наследственности
Молекулярные основы наследственности
 1. Классическая генетика Ø 1865 г. - Г. Мендель - закономерности наследования признаков В основе лежит представление о дискретности – генетический материал состоит из дискретных единиц наследственности – генов. Ø 1930 г. - Т. Морганом и его сотрудниками была доказана физическая связь между геном и хромосомой – хромосомная теория наследственности. Ø 1950 г. – концепция гена – оставалась лишенной материального содержания § В 1950 г. Г. Меллер писал: «…. истинная сущность генетической теории все еще покоится в глубинах неизвестного» . § К началу 40 -ых годов в результате принципиально новых подходов (физических и химических методов) создались реальные условия для раскрытия материальной основы гена, ранее абстрактного и неделимого.
1. Классическая генетика Ø 1865 г. - Г. Мендель - закономерности наследования признаков В основе лежит представление о дискретности – генетический материал состоит из дискретных единиц наследственности – генов. Ø 1930 г. - Т. Морганом и его сотрудниками была доказана физическая связь между геном и хромосомой – хромосомная теория наследственности. Ø 1950 г. – концепция гена – оставалась лишенной материального содержания § В 1950 г. Г. Меллер писал: «…. истинная сущность генетической теории все еще покоится в глубинах неизвестного» . § К началу 40 -ых годов в результате принципиально новых подходов (физических и химических методов) создались реальные условия для раскрытия материальной основы гена, ранее абстрактного и неделимого.
 2. Нуклеиновые кислоты – носители наследственной информации Ø 1869 г. - швейцарский биохимик Фридрих Мишер, выделил из клеток гноя вещество и назвал его «нуклеином» . Ø Два типа нуклеиновых кислот: § § дезоксирибонуклеиновая кислота (ДНК) рибонуклеиновая кислота (РНК). С помощью химического анализа и гистохимических методов окрашивания установлено: Ø ДНК содержится в ядре клетки и, по-видимому, локализуется в хромосомах. ü В клеточном ядре ДНК обнаруживается в виде нуклеопротеида. Ø РНК обнаруживается повсеместно – и в ядре, и в цитоплазме.
2. Нуклеиновые кислоты – носители наследственной информации Ø 1869 г. - швейцарский биохимик Фридрих Мишер, выделил из клеток гноя вещество и назвал его «нуклеином» . Ø Два типа нуклеиновых кислот: § § дезоксирибонуклеиновая кислота (ДНК) рибонуклеиновая кислота (РНК). С помощью химического анализа и гистохимических методов окрашивания установлено: Ø ДНК содержится в ядре клетки и, по-видимому, локализуется в хромосомах. ü В клеточном ядре ДНК обнаруживается в виде нуклеопротеида. Ø РНК обнаруживается повсеместно – и в ядре, и в цитоплазме.
 Доказательства генетической роли ДНК q До 1944 г. о химическом составе и структуре хромосом и генов было известно крайне мало. q Генетические функции хромосом, такие как способность определять развитие признаков, способность к самоудвоению до начала 40 -х годов XX в. большинство ученых связывали с белками, так как белок может принимать множество уникальных конфигураций.
Доказательства генетической роли ДНК q До 1944 г. о химическом составе и структуре хромосом и генов было известно крайне мало. q Генетические функции хромосом, такие как способность определять развитие признаков, способность к самоудвоению до начала 40 -х годов XX в. большинство ученых связывали с белками, так как белок может принимать множество уникальных конфигураций.
 Доказательства генетической роли ДНК q Косвенные доказательства Ø ДНК присутствует во всех клетках – растительных, животных и бактериальных и содержится фактически только в ядрах этих клеток. Ø Все соматические клетки одного вида содержат строго постоянное количество ДНК независимо от их функциональной дифференцировки и метаболического состояния в противоположность РНК. Ø В гаплоидных зародышевых клетках количество ДНК составляет половину количества ДНК в диплоидных соматических клетках, то есть количество ДНК изменяется прямопропорционально плоидности соматических клеток. Ø 4. Обнаружена прямая зависимость между поглощением УФ лучей определенной волны и количеством наследственных изменений у микроорганизмов. Максимальным мутагенным действием обладают УФ лучи той части спектра, (2537 А), в которой специфически поглощают нуклеиновые кислоты.
Доказательства генетической роли ДНК q Косвенные доказательства Ø ДНК присутствует во всех клетках – растительных, животных и бактериальных и содержится фактически только в ядрах этих клеток. Ø Все соматические клетки одного вида содержат строго постоянное количество ДНК независимо от их функциональной дифференцировки и метаболического состояния в противоположность РНК. Ø В гаплоидных зародышевых клетках количество ДНК составляет половину количества ДНК в диплоидных соматических клетках, то есть количество ДНК изменяется прямопропорционально плоидности соматических клеток. Ø 4. Обнаружена прямая зависимость между поглощением УФ лучей определенной волны и количеством наследственных изменений у микроорганизмов. Максимальным мутагенным действием обладают УФ лучи той части спектра, (2537 А), в которой специфически поглощают нуклеиновые кислоты.
 Прямые доказательства Объекты исследования наследственности и природы генетического материала - бактерии, вирусы. Ø Преимущества микроорганизмов как генетических объектов: 1. Они обычно гаплоидны, нет перекрытия рецессивных генов доминантными, что позволяет сразу проявляться мутантному гену. 2. При размножении создают точные копии самих себя. 3. Скорость размножения. В питательной среде за один день плотность популяции, возникшей из одной клетки Escherichia coli, может достигнуть 2 -3 109 бактерий в миллилитре. ü Наиболее интенсивно исследуемый вид бактерий Escherichia coli – кишечная палочка.
Прямые доказательства Объекты исследования наследственности и природы генетического материала - бактерии, вирусы. Ø Преимущества микроорганизмов как генетических объектов: 1. Они обычно гаплоидны, нет перекрытия рецессивных генов доминантными, что позволяет сразу проявляться мутантному гену. 2. При размножении создают точные копии самих себя. 3. Скорость размножения. В питательной среде за один день плотность популяции, возникшей из одной клетки Escherichia coli, может достигнуть 2 -3 109 бактерий в миллилитре. ü Наиболее интенсивно исследуемый вид бактерий Escherichia coli – кишечная палочка.
 Прямые доказательства v В период с 1944 по 1953 гг. в проведенных исследованиях получены доказательства того, что генетическую основу жизненных процессов составляет молекула ДНК. 2. 1 ДНК – трансформирующий (Diplococcus pneumoniae). фактор пневмококка Фредерик Гриффит в 1928 г. при изучении пневмококковой инфекции у мышей - обнаружил явление трансформации. S - вирулентный штамм - гладкие блестящие колонии: тип I, тип III. R - невирулентный штамм на твердой питательной среде образуют шероховатые колонии. Мутация S R идет исключительно в одном направлении, при обратных мутациях R S : IS I R, II S I I R, III S III R и т. д
Прямые доказательства v В период с 1944 по 1953 гг. в проведенных исследованиях получены доказательства того, что генетическую основу жизненных процессов составляет молекула ДНК. 2. 1 ДНК – трансформирующий (Diplococcus pneumoniae). фактор пневмококка Фредерик Гриффит в 1928 г. при изучении пневмококковой инфекции у мышей - обнаружил явление трансформации. S - вирулентный штамм - гладкие блестящие колонии: тип I, тип III. R - невирулентный штамм на твердой питательной среде образуют шероховатые колонии. Мутация S R идет исключительно в одном направлении, при обратных мутациях R S : IS I R, II S I I R, III S III R и т. д
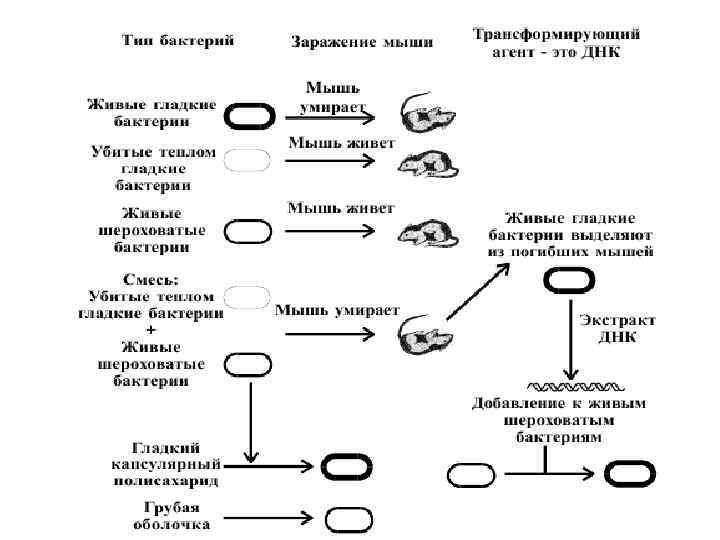
 v Вывод: какой-то компонент мертвых бактерий типа III S может трансформировать живых бактерий типа II R так, что они начинают синтезировать капсульный полисахарид типа III S. q Метод in vitro (в пробирке) позволил исследовать природу трансформирующего фактора убитых нагреванием клеток III S непосредственно, не вводя их мышам и не дожидаясь гибели последних. Ø В 1944 г. О. Эвери, К. Мак-Леод и М. Мак-Карти показали трансформирующий фактор – это ДНК. ü Добавление в растущую культуру клеток II R очищенной ДНК пнемококка III S оказалось достаточным для возникновения способности синтезировать капсульный полисахарид типа III S. ü Добавление фермента дезоксирибонуклеазы, расщепляющего ДНК, необратимо инактивирует трансформирующий фактор.
v Вывод: какой-то компонент мертвых бактерий типа III S может трансформировать живых бактерий типа II R так, что они начинают синтезировать капсульный полисахарид типа III S. q Метод in vitro (в пробирке) позволил исследовать природу трансформирующего фактора убитых нагреванием клеток III S непосредственно, не вводя их мышам и не дожидаясь гибели последних. Ø В 1944 г. О. Эвери, К. Мак-Леод и М. Мак-Карти показали трансформирующий фактор – это ДНК. ü Добавление в растущую культуру клеток II R очищенной ДНК пнемококка III S оказалось достаточным для возникновения способности синтезировать капсульный полисахарид типа III S. ü Добавление фермента дезоксирибонуклеазы, расщепляющего ДНК, необратимо инактивирует трансформирующий фактор.
 Ø О. Эвери, Мак-Леод и Мак-Карти впервые показали: наследственная способность клетки осуществлять определенную биосинтетическую функцию может передаваться другой клетке вместе с очищенной ДНК. q Механизм трансформации: фрагменты ДНК, в том числе содержащие ген, ответственный за синтез полисахарида, из убитых нагреванием клеток S попадают в R-клетки и посредством рекомбинации включаются в их ДНК.
Ø О. Эвери, Мак-Леод и Мак-Карти впервые показали: наследственная способность клетки осуществлять определенную биосинтетическую функцию может передаваться другой клетке вместе с очищенной ДНК. q Механизм трансформации: фрагменты ДНК, в том числе содержащие ген, ответственный за синтез полисахарида, из убитых нагреванием клеток S попадают в R-клетки и посредством рекомбинации включаются в их ДНК.
 q Фундаментальное значение этого открытия не было оценено по следующим причинам: 1. Знания о химической структуре ДНК были неполными и неправильно интерпретировались. Считалось, что ДНК недостаточно сложноорганизованное химическое вещество для содержания огромного количества информации. 2. Многие ученые полагали, что химической основой генов служат белки, относительно которых было известно, что они устроены сложно, тогда как ДНК была оставлена роль структурного вещества. 3. Изучение наследственных основ бактерий в 1944 г. только начиналось, и не было установлено, что бактериальные гены во всех отношениях аналогичны генам высших организмов.
q Фундаментальное значение этого открытия не было оценено по следующим причинам: 1. Знания о химической структуре ДНК были неполными и неправильно интерпретировались. Считалось, что ДНК недостаточно сложноорганизованное химическое вещество для содержания огромного количества информации. 2. Многие ученые полагали, что химической основой генов служат белки, относительно которых было известно, что они устроены сложно, тогда как ДНК была оставлена роль структурного вещества. 3. Изучение наследственных основ бактерий в 1944 г. только начиналось, и не было установлено, что бактериальные гены во всех отношениях аналогичны генам высших организмов.
 2. 2 Нуклеиновые кислоты – наследственный материал вирусов
2. 2 Нуклеиновые кислоты – наследственный материал вирусов
 Структура бактериофага Т 4 Во время сборки к головке добавляются части хвоста. А затем хвостовые фибриллы.
Структура бактериофага Т 4 Во время сборки к головке добавляются части хвоста. А затем хвостовые фибриллы.
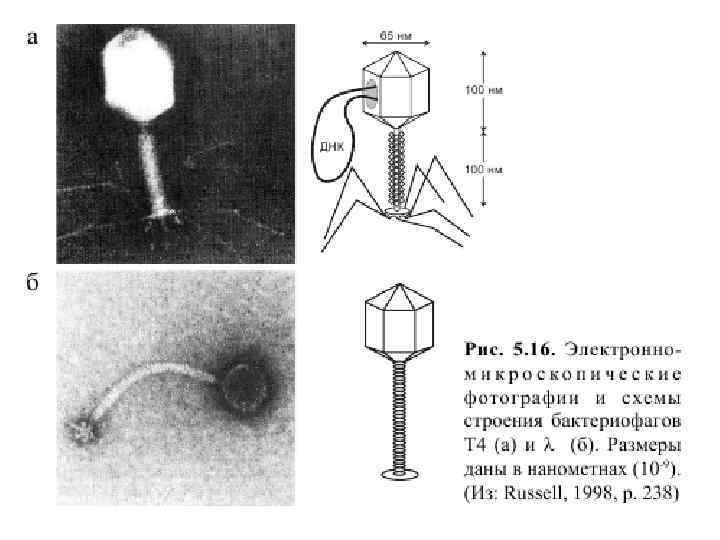
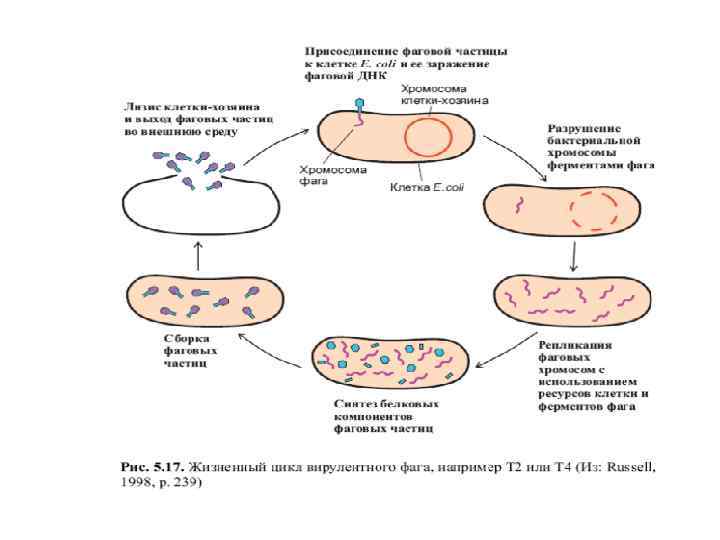
 2. 2 Нуклеиновые кислоты – наследственный материал вирусов q В 1952 г. Альфред Херши и Марта Чейз - доказали роль ДНК для синтеза новых вирусных частиц. Ø Белок фага был помечен радиоактивным изотопом серы 35 S, лишь белковая составляющая содержит серу в составе аминокислот метионина и цистеина. ДНК пометили радиоактивным изотопом фосфора 32 Р, около 99 % 32 Р в фаге приходится на ДНК. Ø В потомстве фага, образованном после инфицирования было найдено примерно 30 % исходной метки 32 Р, а от исходного белка менее 1 %.
2. 2 Нуклеиновые кислоты – наследственный материал вирусов q В 1952 г. Альфред Херши и Марта Чейз - доказали роль ДНК для синтеза новых вирусных частиц. Ø Белок фага был помечен радиоактивным изотопом серы 35 S, лишь белковая составляющая содержит серу в составе аминокислот метионина и цистеина. ДНК пометили радиоактивным изотопом фосфора 32 Р, около 99 % 32 Р в фаге приходится на ДНК. Ø В потомстве фага, образованном после инфицирования было найдено примерно 30 % исходной метки 32 Р, а от исходного белка менее 1 %.
 v Вывод для образования копий фага в зараженной бактериальной клетке существенна лишь ДНК родительского фага, тогда как новые фаговые частицы содержат как ДНК, так и белок. Ø На примере бактериофага Т 2 еще раз был подтвержден вывод о генетической роли ДНК. общий Ø Эксперимент был признан в качестве решающего доказательства генетической роли ДНК. Ø Было известно, что по характеру наследования признаков бактериофаг аналогичен высшим организмам. Его признаки точно воспроизводятся и подчиняются тем же правилам, что делает возможным линейное картирование мутаций. Ø К 1952 г. было показано, что ДНК обладает достаточной химической сложностью для носителя наследственной информации.
v Вывод для образования копий фага в зараженной бактериальной клетке существенна лишь ДНК родительского фага, тогда как новые фаговые частицы содержат как ДНК, так и белок. Ø На примере бактериофага Т 2 еще раз был подтвержден вывод о генетической роли ДНК. общий Ø Эксперимент был признан в качестве решающего доказательства генетической роли ДНК. Ø Было известно, что по характеру наследования признаков бактериофаг аналогичен высшим организмам. Его признаки точно воспроизводятся и подчиняются тем же правилам, что делает возможным линейное картирование мутаций. Ø К 1952 г. было показано, что ДНК обладает достаточной химической сложностью для носителя наследственной информации.
 С помощью химических методов можно разделить") 2. 3 Опыты на вирусе табачной мозаики (ВТМ) С помощью химических методов можно разделить РНК и белок вируса. стандартный штамм ВТМ (тип S) = белок S + S РНК (отсутствуют метионин и гистидин) штамм Н R = белок Н R + Н R РНК (имеются метионин и гистидин) сконструирован гибридный вирус = очищенный белок НR + очищенная S РНК v. Вывод Ø Потомство гибридных вирусов имело тип белковой соответствующий типу РНК в сконструированном вирусе. Ø Состав белковой исключительно РНК. оболочки не наследовался. Он оболочки, определялся
2. 3 Опыты на вирусе табачной мозаики (ВТМ) С помощью химических методов можно разделить РНК и белок вируса. стандартный штамм ВТМ (тип S) = белок S + S РНК (отсутствуют метионин и гистидин) штамм Н R = белок Н R + Н R РНК (имеются метионин и гистидин) сконструирован гибридный вирус = очищенный белок НR + очищенная S РНК v. Вывод Ø Потомство гибридных вирусов имело тип белковой соответствующий типу РНК в сконструированном вирусе. Ø Состав белковой исключительно РНК. оболочки не наследовался. Он оболочки, определялся
 Вывод Результаты проведенных исследований доказали: нуклеиновые кислоты являются носителем наследственной информации во всех организмах (генетический материал всех организмов, исключая вирусов, представлен ДНК).
Вывод Результаты проведенных исследований доказали: нуклеиновые кислоты являются носителем наследственной информации во всех организмах (генетический материал всех организмов, исключая вирусов, представлен ДНК).
 3. Структурный анализ ДНК Величайшим открытием в истории биологии было открытие структуры ДНК. В 1953 г. Джеймс Уотсон и Френсис Крик (работали в Кавендишской лаборатории в Кембримдже) на основании результатов: Ø состава оснований в молекулах ДНК после гидролиза Ø дифракционного рентгеновских лучей анализа v Предположили молекула ДНК двуспиральная. структуры ДНК с помощью
3. Структурный анализ ДНК Величайшим открытием в истории биологии было открытие структуры ДНК. В 1953 г. Джеймс Уотсон и Френсис Крик (работали в Кавендишской лаборатории в Кембримдже) на основании результатов: Ø состава оснований в молекулах ДНК после гидролиза Ø дифракционного рентгеновских лучей анализа v Предположили молекула ДНК двуспиральная. структуры ДНК с помощью
 и английский физик и Френсис Крик (1916") Американский биохимик Джеймс Уотсон (1928 г. ) и английский физик и Френсис Крик (1916 г. )
Американский биохимик Джеймс Уотсон (1928 г. ) и английский физик и Френсис Крик (1916 г. )
 Химический состав и строение нуклеиновых кислот ДНК – это полимер, состоящий из последовательности химически связанных между собой нуклеотидов. Ø Нуклеотид состоит из трех химически различных компонентов, соединенных ковалентными связями: содержит гетероциклическое кольцо из атомов углерода и азота (азотистое основание), пятиуглеродное сахарное кольцо (пентозу – С 5 Н 10 О 4) и фосфатную группу, сообщающей ДНК кислотные свойства. Два типа азотистых оснований: § пуриновые - основания, имеющие два конденсированных кольца: одно пятичленное и второе шестичленное, относятся аденин (А) и гуанин (Г). § пиримидиновые - в молекуле которых имеется только одно шестичленное кольцо, относятся тимин (Т) и цитозин (Ц). Ø Нуклеозид - соединение азотистого основания с каждым остатком 2 дезоксирибозы гликозидной связью между С-1 пентозного кольца и N-3 пиримидина и N-9 пурина (гликозид –дезоксиаденозин, дезоксицитидин, дезоксигуанозин, дезокситимидин). • Соседние нуклеозиды связаны остатками фосфорной кислоты через С 3 одного остатка дезоксирибозы и С-5 соседнего остатка (связь 3 -5 -3 -5). • Нуклеозид с одной или несколькими фосфатными группами называется нуклеотидом
Химический состав и строение нуклеиновых кислот ДНК – это полимер, состоящий из последовательности химически связанных между собой нуклеотидов. Ø Нуклеотид состоит из трех химически различных компонентов, соединенных ковалентными связями: содержит гетероциклическое кольцо из атомов углерода и азота (азотистое основание), пятиуглеродное сахарное кольцо (пентозу – С 5 Н 10 О 4) и фосфатную группу, сообщающей ДНК кислотные свойства. Два типа азотистых оснований: § пуриновые - основания, имеющие два конденсированных кольца: одно пятичленное и второе шестичленное, относятся аденин (А) и гуанин (Г). § пиримидиновые - в молекуле которых имеется только одно шестичленное кольцо, относятся тимин (Т) и цитозин (Ц). Ø Нуклеозид - соединение азотистого основания с каждым остатком 2 дезоксирибозы гликозидной связью между С-1 пентозного кольца и N-3 пиримидина и N-9 пурина (гликозид –дезоксиаденозин, дезоксицитидин, дезоксигуанозин, дезокситимидин). • Соседние нуклеозиды связаны остатками фосфорной кислоты через С 3 одного остатка дезоксирибозы и С-5 соседнего остатка (связь 3 -5 -3 -5). • Нуклеозид с одной или несколькими фосфатными группами называется нуклеотидом
 Азотистые основания
Азотистые основания
 Нуклеозид и нуклеотид
Нуклеозид и нуклеотид
 Фосфодиэфирная связь между двумя нуклеотидами
Фосфодиэфирная связь между двумя нуклеотидами
 – фосфодиэфирных связей,") v Таким образом, полинуклеотидная цепь ДНК состоит из ряда (5 -3) – фосфодиэфирных связей, когда остаток фосфорной кислоты соединяется эфирными связями с двумя гидроксильными группами сахаров в позиции С-3 – С-5, образующих остов молекулы, к которому присоединяются азотистые основания. Ø Короткие цепочки из одного-двух десятков нуклеотидов называются олигонуклеотидами, а более длинные – полинуклеотидами. Ø Интактная молекула ДНК в зависимости вида организмов содержит от нескольких тысяч до многих миллионов нуклеотидов. ДНК хромосомы 1 человека равна 263 м. п. н. Ø Интактная молекула РНК содержит от 100 до 100 000 нуклеотидов. Ø Любая молекула нуклеиновой кислоты записывается как последовательность нуклеотидов с различными основаниями, например – АТАЦГАГААААААЦГЦА. Эта последовательность нуклеотидов является первичной структурой молекулы нуклеиновой кислоты. Ø Вторичная структура – двойная цепочка ДНК.
v Таким образом, полинуклеотидная цепь ДНК состоит из ряда (5 -3) – фосфодиэфирных связей, когда остаток фосфорной кислоты соединяется эфирными связями с двумя гидроксильными группами сахаров в позиции С-3 – С-5, образующих остов молекулы, к которому присоединяются азотистые основания. Ø Короткие цепочки из одного-двух десятков нуклеотидов называются олигонуклеотидами, а более длинные – полинуклеотидами. Ø Интактная молекула ДНК в зависимости вида организмов содержит от нескольких тысяч до многих миллионов нуклеотидов. ДНК хромосомы 1 человека равна 263 м. п. н. Ø Интактная молекула РНК содержит от 100 до 100 000 нуклеотидов. Ø Любая молекула нуклеиновой кислоты записывается как последовательность нуклеотидов с различными основаниями, например – АТАЦГАГААААААЦГЦА. Эта последовательность нуклеотидов является первичной структурой молекулы нуклеиновой кислоты. Ø Вторичная структура – двойная цепочка ДНК.
 В 1950 г. Эдвином Чаргаффом с помощью хромотографии были получены следующие важные данные о химическом составе молекулы ДНК. 1. Молярное содержание аденина равно содержанию тимина, а молярное содержание гуанина равно содержанию цитозина [А] = [Т], [Г] = [Ц]. 2. Независимо от источника получения, ДНК содержит равное количество пуриновых и пиримидиновых оснований. ( [А] +[ Г ] ) = ( [ Т] + [Ц] ). v Эти равенства называются правилами Чаргаффа. Согласно правилам Чаргаффа, в молекуле ДНК пары оснований связаны между собой в определенном порядке.
В 1950 г. Эдвином Чаргаффом с помощью хромотографии были получены следующие важные данные о химическом составе молекулы ДНК. 1. Молярное содержание аденина равно содержанию тимина, а молярное содержание гуанина равно содержанию цитозина [А] = [Т], [Г] = [Ц]. 2. Независимо от источника получения, ДНК содержит равное количество пуриновых и пиримидиновых оснований. ( [А] +[ Г ] ) = ( [ Т] + [Ц] ). v Эти равенства называются правилами Чаргаффа. Согласно правилам Чаргаффа, в молекуле ДНК пары оснований связаны между собой в определенном порядке.
 q Ливен предполагал – ДНК представляет тетрануклеотидную последовательность, например р. Ар. Цр. Гр. Т, многократно повторенную так, что она образует полимер вида (р. Ар. Цр. Гр. Т)n. q Исследования Чаргаффа по составу ДНК многих различных организмов показали, что состав оснований в ДНК различен у различных видов. Ø В зависимости от видовой принадлежности меняется отношение (А + Т)/ (Г + Ц) - так называемое отношение оснований. Для любого данного вида эта величина остается постоянной. Ø У разных видов бактерий величина отношения оснований колеблется в пределах 0, 35 – 2, 7. v Вывод ü Исключить возможность строения ДНК из одинаковых тетрануклеотидов. ü ДНК обладает сложностью, необходимой для передачи наследственной информации.
q Ливен предполагал – ДНК представляет тетрануклеотидную последовательность, например р. Ар. Цр. Гр. Т, многократно повторенную так, что она образует полимер вида (р. Ар. Цр. Гр. Т)n. q Исследования Чаргаффа по составу ДНК многих различных организмов показали, что состав оснований в ДНК различен у различных видов. Ø В зависимости от видовой принадлежности меняется отношение (А + Т)/ (Г + Ц) - так называемое отношение оснований. Для любого данного вида эта величина остается постоянной. Ø У разных видов бактерий величина отношения оснований колеблется в пределах 0, 35 – 2, 7. v Вывод ü Исключить возможность строения ДНК из одинаковых тетрануклеотидов. ü ДНК обладает сложностью, необходимой для передачи наследственной информации.
, проведенный Р. Франклин и Уилкинсоном в 1950–") Рентгеноструктурный дифракционный анализ (анализ с помощью Х-лучей), проведенный Р. Франклин и Уилкинсоном в 1950– 1953 гг. ,
Рентгеноструктурный дифракционный анализ (анализ с помощью Х-лучей), проведенный Р. Франклин и Уилкинсоном в 1950– 1953 гг. ,
 Выявлены следующие особенности молекулярной структуры ДНК: 1. Пуриновые и пиримидиновые основания представляют собой плоские структуры, расположенные в плоскостях, перпендикулярных длинной оси полинуклеотидной цепи, друг над другом, расстояние между соседними основаниями составляет 3, 4 А (10 А = 1 нм). 2. Полинуклеотидная цепь закручена вокруг некой воображаемой оси, шаг этой спирали равен 34 А, диаметр 20 А. 3. Плотность свидетельствовала, что ДНК состоит более чем из одной спирально закрученной полинуклеотидной цепи. v Р. Франклин предположила, что молекула ДНК имеет спиральную структуру с шагом между витками 3, 4, но точной модели ДНК построено не было. v На основании этих и других данных Л. Полинг ошибочно предположил, что ДНК представляет собой тройную спираль
Выявлены следующие особенности молекулярной структуры ДНК: 1. Пуриновые и пиримидиновые основания представляют собой плоские структуры, расположенные в плоскостях, перпендикулярных длинной оси полинуклеотидной цепи, друг над другом, расстояние между соседними основаниями составляет 3, 4 А (10 А = 1 нм). 2. Полинуклеотидная цепь закручена вокруг некой воображаемой оси, шаг этой спирали равен 34 А, диаметр 20 А. 3. Плотность свидетельствовала, что ДНК состоит более чем из одной спирально закрученной полинуклеотидной цепи. v Р. Франклин предположила, что молекула ДНК имеет спиральную структуру с шагом между витками 3, 4, но точной модели ДНК построено не было. v На основании этих и других данных Л. Полинг ошибочно предположил, что ДНК представляет собой тройную спираль
 Модель молекулы ДНК
Модель молекулы ДНК
 ДНК показывает, что ее структура стабилизируется водородными связями. Титрование и") Кислотно-щелочное титрирование нативной (природной) ДНК показывает, что ее структура стабилизируется водородными связями. Титрование и нагревание нативной ДНК вызывает заметные изменения ее физических свойств – вязкости, переводя ее в денатурированную форму, причем, ковалентные связи не разрушаются.
Кислотно-щелочное титрирование нативной (природной) ДНК показывает, что ее структура стабилизируется водородными связями. Титрование и нагревание нативной ДНК вызывает заметные изменения ее физических свойств – вязкости, переводя ее в денатурированную форму, причем, ковалентные связи не разрушаются.
 Модель структуры ДНК Уотсона–Крика Носитель генетической информации должна удовлетворять двум основным требованиям: § воспроизводиться (реплицироваться) с высокой точностью § детерминировать (кодировать) синтез белковых молекул. v 1953 г. американский биохимик Дж. Уотсон (1928 г. ) английский физик Ф. Крик (1916 г. ), синтезировав данные химического и рентгеноструктурного анализов, предложили модель структуры ДНК – двойная спираль – состоит из двух попарно соединенных полинуклеотидных цепей, закрученных в двойную спираль. Была построена молекулярная модель, на которой точно выдерживались все межатомные расстояния.
Модель структуры ДНК Уотсона–Крика Носитель генетической информации должна удовлетворять двум основным требованиям: § воспроизводиться (реплицироваться) с высокой точностью § детерминировать (кодировать) синтез белковых молекул. v 1953 г. американский биохимик Дж. Уотсон (1928 г. ) английский физик Ф. Крик (1916 г. ), синтезировав данные химического и рентгеноструктурного анализов, предложили модель структуры ДНК – двойная спираль – состоит из двух попарно соединенных полинуклеотидных цепей, закрученных в двойную спираль. Была построена молекулярная модель, на которой точно выдерживались все межатомные расстояния.
 Принципы строения ДНК Ø Нерегулярность. Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
Принципы строения ДНК Ø Нерегулярность. Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
.") Ø Две цепи антипараллельные в отношении расположения связей 3 -5 (ориентированы в противоположных направлениях). 3`-конец одной расположен напротив 5`-конца другой. Ø Удерживаются посредством водородных связей между азотистыми основаниями аденином и тимином, гуанином и цитозином, которые обращены во внутрь спирали и направлены навстречу другу. Специфичность спаривания (ключевое свойство модели) определяет только комплементарность оснований в двух полинуклеотидных цепях, но не накладывает никакого ограничения на саму последовательность оснований. Спаривание оснований занимает центральное место во всех процессах, протекающих с участием нуклеиновых кислот.
Ø Две цепи антипараллельные в отношении расположения связей 3 -5 (ориентированы в противоположных направлениях). 3`-конец одной расположен напротив 5`-конца другой. Ø Удерживаются посредством водородных связей между азотистыми основаниями аденином и тимином, гуанином и цитозином, которые обращены во внутрь спирали и направлены навстречу другу. Специфичность спаривания (ключевое свойство модели) определяет только комплементарность оснований в двух полинуклеотидных цепях, но не накладывает никакого ограничения на саму последовательность оснований. Спаривание оснований занимает центральное место во всех процессах, протекающих с участием нуклеиновых кислот.
 Ø Наличие регулярной вторичной структуры. Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью. • В пределах любой последовательность оснований. цепи допускается любая В полинуклеотидной цепочке ДНК может храниться разнообразная генетическая информация. Например, если последовательность ДНК содержит 1000 нуклеотидов, то ее исходя из четырех разных нуклеотидов эту последовательность можно записать различными способами 41000 раз.
Ø Наличие регулярной вторичной структуры. Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью. • В пределах любой последовательность оснований. цепи допускается любая В полинуклеотидной цепочке ДНК может храниться разнообразная генетическая информация. Например, если последовательность ДНК содержит 1000 нуклеотидов, то ее исходя из четырех разных нуклеотидов эту последовательность можно записать различными способами 41000 раз.
 Модель структуры ДНК Уотсона – Крика объясняет как способность генов к самоудвоению, так и их информационные свойства. Поскольку две цепи ДНК комплементарны, каждая из них при расплетании двойной спирали может служить матрицей для синтеза новой комплементарной цепи, образуя при этом две одинаковые дочерние молекулы (полуконсервативная модель репликации).
Модель структуры ДНК Уотсона – Крика объясняет как способность генов к самоудвоению, так и их информационные свойства. Поскольку две цепи ДНК комплементарны, каждая из них при расплетании двойной спирали может служить матрицей для синтеза новой комплементарной цепи, образуя при этом две одинаковые дочерние молекулы (полуконсервативная модель репликации).
 Альтернативные формы ДНК
Альтернативные формы ДНК
 Альтернативные формы ДНК А-, В- и С- формы, D- и Е-формы, Р-ДНК, Z-форма Отличаются числом нуклеотидов на виток и расстоянием между нуклеотидами, шагом спирали и углом наклона между плоскостью оснований и осью спирали. Конфигурация зависит от содержания солей в препарате, нуклеотидного состава ДНК и относительной влажности препарата. В-форма Характеризуется плоскопараллельным расположением пар нуклеотидных оснований внутри двойной спирали. Считают, что именно в такой форме ДНК обычно находится в живой клетке. А-форма Основания наклонены по отношению к оси спирали – угол наклона 20 , число оснований на виток больше, чем в В-форме – 11, диаметр 23. А-форма близка к структуре гибридов ДНК-РНК и двуспиральных участков РНК, этим она интересна с биологической точки зрения.
Альтернативные формы ДНК А-, В- и С- формы, D- и Е-формы, Р-ДНК, Z-форма Отличаются числом нуклеотидов на виток и расстоянием между нуклеотидами, шагом спирали и углом наклона между плоскостью оснований и осью спирали. Конфигурация зависит от содержания солей в препарате, нуклеотидного состава ДНК и относительной влажности препарата. В-форма Характеризуется плоскопараллельным расположением пар нуклеотидных оснований внутри двойной спирали. Считают, что именно в такой форме ДНК обычно находится в живой клетке. А-форма Основания наклонены по отношению к оси спирали – угол наклона 20 , число оснований на виток больше, чем в В-форме – 11, диаметр 23. А-форма близка к структуре гибридов ДНК-РНК и двуспиральных участков РНК, этим она интересна с биологической точки зрения.
 Z-форма представляет собой наиболее резкий контраст с выше рассмотренными формами (открыта в 1979 г. Эндрю Ванг и Александром Рич). Ø Левосторонняя, тогда как все остальные – правосторонние. Ø Число пар оснований на виток – 12, то есть она менее скрученная и более тонкая (d = 18). Ø Сахарофосфатный остов образует вдоль спирали зигзагообразную линию. Ø Найдена Z-форма двойной спирали в полимерах, имеющих многократно повторяющиеся динуклеотидные пары: polu-d (GC ) n и polu-d ( AC)n – 26 -32 дуплета. CG TG Ø Белки оказывают значительное влияние на возможность перехода из В-формы в Z-форму. Ø Присутствие Z-ДНК in vivo пока не доказано.
Z-форма представляет собой наиболее резкий контраст с выше рассмотренными формами (открыта в 1979 г. Эндрю Ванг и Александром Рич). Ø Левосторонняя, тогда как все остальные – правосторонние. Ø Число пар оснований на виток – 12, то есть она менее скрученная и более тонкая (d = 18). Ø Сахарофосфатный остов образует вдоль спирали зигзагообразную линию. Ø Найдена Z-форма двойной спирали в полимерах, имеющих многократно повторяющиеся динуклеотидные пары: polu-d (GC ) n и polu-d ( AC)n – 26 -32 дуплета. CG TG Ø Белки оказывают значительное влияние на возможность перехода из В-формы в Z-форму. Ø Присутствие Z-ДНК in vivo пока не доказано.
 Водородные связи и анализ структуры нуклеиновых кислот Природа водородных связей уникальна. При слабом нагревании двойная спираль денатурирует и раскручивается. При медленном охлаждении смеси комплементарных одноцепочечных молекул ДНК они реассоциируют, вновь образуя двойную спираль. У молекул с более высоким содержанием пар Г-Ц (три водородных связи) по сравнению с А-Т температура плавления выше. Методы молекулярной гибридизации В основе методов молекулярной гибридизации лежит способность нуклеиновых кислот к обратимой денатурации. При соответствующей температуре между ДНК разных видов, между ДНК и РНК образуются гибридные молекулы, что говорит о сходстве их последовательностей.
Водородные связи и анализ структуры нуклеиновых кислот Природа водородных связей уникальна. При слабом нагревании двойная спираль денатурирует и раскручивается. При медленном охлаждении смеси комплементарных одноцепочечных молекул ДНК они реассоциируют, вновь образуя двойную спираль. У молекул с более высоким содержанием пар Г-Ц (три водородных связи) по сравнению с А-Т температура плавления выше. Методы молекулярной гибридизации В основе методов молекулярной гибридизации лежит способность нуклеиновых кислот к обратимой денатурации. При соответствующей температуре между ДНК разных видов, между ДНК и РНК образуются гибридные молекулы, что говорит о сходстве их последовательностей.
 Генетический код Первичная структура каждого белка в организме кодируется определенным геном. Ø В состав белков входит 20 основных различных аминокислот ØВ ДНК содержатся нуклеотиды только четырех типов Соответствие между нуклеотидными и аминокислотными последовательностями не может быть построено по принципу «один к одному» . Каждой данной аминокислоте соответствовала бы комбинация из нескольких нуклеотидов v Последовательность нуклеотидов в молекуле ДНК, опеределяющая последовательность аминокислот в белке, называется генетическим кодом (соответствие меду нуклеотидными триплетами и кодируемыми ими аминокислотами).
Генетический код Первичная структура каждого белка в организме кодируется определенным геном. Ø В состав белков входит 20 основных различных аминокислот ØВ ДНК содержатся нуклеотиды только четырех типов Соответствие между нуклеотидными и аминокислотными последовательностями не может быть построено по принципу «один к одному» . Каждой данной аминокислоте соответствовала бы комбинация из нескольких нуклеотидов v Последовательность нуклеотидов в молекуле ДНК, опеределяющая последовательность аминокислот в белке, называется генетическим кодом (соответствие меду нуклеотидными триплетами и кодируемыми ими аминокислотами).
 Особенности генетического кода 1. Триплетный – одна аминокислота кодируется последовательностью из трех нуклеотидов, называемой кодоном. Всего 64 кодона = 43, из них 61 используется для кодирования аминокислот (смысловые кодоны). Один из них стартовый – метиониновый (АUG). Триплет или кодон - последовательность из трех нуклеотидов, кодирующая одну аминокислоту. 2. Недвусмысленный – каждый триплет кодирует только одну аминокислоту. Исключение составляет кодон AUG. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин.
Особенности генетического кода 1. Триплетный – одна аминокислота кодируется последовательностью из трех нуклеотидов, называемой кодоном. Всего 64 кодона = 43, из них 61 используется для кодирования аминокислот (смысловые кодоны). Один из них стартовый – метиониновый (АUG). Триплет или кодон - последовательность из трех нуклеотидов, кодирующая одну аминокислоту. 2. Недвусмысленный – каждый триплет кодирует только одну аминокислоту. Исключение составляет кодон AUG. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин.
 3. Вырожденный – одной аминокислоте соответствует более чем один кодон. Все аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом. § Лейцин, например, кодируется шестью кодонами. Кодоны «синонимы» , различаются друг от друга только по последнему нуклеотиду. v Синонимичные кодоны используются различными организмами с разной частотой. ü Из четырех кодонов для глицина GGA используется в структурных генах человека в 26% случаев, а в Escherichia coli — в 9%. ü Такая же ситуация наблюдается и для стоп-кодонов. Так, у человека частота использования кодонов UAA, UAG и UGA составляет 0, 22, 0, 17 и 0, 61 соответственно, а у E. coli — 0, 62, 0, 09 и 0, 30. Несмотря на все эти различия, генетический код у всех организмов, за редким исключением, одинаков.
3. Вырожденный – одной аминокислоте соответствует более чем один кодон. Все аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом. § Лейцин, например, кодируется шестью кодонами. Кодоны «синонимы» , различаются друг от друга только по последнему нуклеотиду. v Синонимичные кодоны используются различными организмами с разной частотой. ü Из четырех кодонов для глицина GGA используется в структурных генах человека в 26% случаев, а в Escherichia coli — в 9%. ü Такая же ситуация наблюдается и для стоп-кодонов. Так, у человека частота использования кодонов UAA, UAG и UGA составляет 0, 22, 0, 17 и 0, 61 соответственно, а у E. coli — 0, 62, 0, 09 и 0, 30. Несмотря на все эти различия, генетический код у всех организмов, за редким исключением, одинаков.
,") 3. Код содержит три бессмысленных кодона ( нонсес кодоны, стопкодоны – УАА. УАГ, УГА), не кодирующие ни какую аминокислоту. 4. Непрерывный С началом транскрипции кодоны м. РНК считываются один за другим без перерывов. 5. Наличие межгенных знаков препинания. ü Определение: ген- это участок ДНК, кодирующий одну полипептидную цепь или одну молекулу t. РНК, r. РНК или s. РНК. ü Гены t. РНК, r. РНК, s. РНК белки не кодируют. ü В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3 -х терминирующих кодонов, или стоп-сигналов: UAA, UAG, UGA. Они терминируют трансляцию. ü Условно к знакам препинания относится и кодон AUG - первый после лидерной последовательности. Он выполняет функцию заглавной буквы. В этой позиции он кодирует формилметионин (у прокариот). 6. Неперекрывающийся ü Каждое основание может принадлежать только одному триплету и не является частью другого триплета. ü Кодоны ни чем не отделены друг от друга. ü Код считывается направлении. с фиксированной точки в пределах гена в одном
3. Код содержит три бессмысленных кодона ( нонсес кодоны, стопкодоны – УАА. УАГ, УГА), не кодирующие ни какую аминокислоту. 4. Непрерывный С началом транскрипции кодоны м. РНК считываются один за другим без перерывов. 5. Наличие межгенных знаков препинания. ü Определение: ген- это участок ДНК, кодирующий одну полипептидную цепь или одну молекулу t. РНК, r. РНК или s. РНК. ü Гены t. РНК, r. РНК, s. РНК белки не кодируют. ü В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3 -х терминирующих кодонов, или стоп-сигналов: UAA, UAG, UGA. Они терминируют трансляцию. ü Условно к знакам препинания относится и кодон AUG - первый после лидерной последовательности. Он выполняет функцию заглавной буквы. В этой позиции он кодирует формилметионин (у прокариот). 6. Неперекрывающийся ü Каждое основание может принадлежать только одному триплету и не является частью другого триплета. ü Кодоны ни чем не отделены друг от друга. ü Код считывается направлении. с фиксированной точки в пределах гена в одном
 Неперекрываемость В 1956 г. Георгий Гамов предложил вариант перекрываемого кода. Согласно Гамовскому коду, каждый нуклеотид, начиная с третьего в гене, входит в состав 3 -х кодонов. Когда генетический код был расшифрован, оказалось, что он неперекрываем, т. е. каждый нуклеотид входит в состав лишь одного кодона.
Неперекрываемость В 1956 г. Георгий Гамов предложил вариант перекрываемого кода. Согласно Гамовскому коду, каждый нуклеотид, начиная с третьего в гене, входит в состав 3 -х кодонов. Когда генетический код был расшифрован, оказалось, что он неперекрываем, т. е. каждый нуклеотид входит в состав лишь одного кодона.
 6. Универсален Он одинаков у всех организмов: одни и те же триплеты кодируют одну и ту же аминокислоту у всех организмов. Отличие митохондриальный генетический код имеет отличия от универсального кода. Кодон Универсальный код Митохондриальные коды Позвоночны е Беспозвоночные Дрожжи Растения UGA STOP Trp Trp STOP AUA Ile Met Met Ile CUA Leu Leu Thr Leu AGA Arg STOP Ser Arg AGG Arg STOP Ser Arg
6. Универсален Он одинаков у всех организмов: одни и те же триплеты кодируют одну и ту же аминокислоту у всех организмов. Отличие митохондриальный генетический код имеет отличия от универсального кода. Кодон Универсальный код Митохондриальные коды Позвоночны е Беспозвоночные Дрожжи Растения UGA STOP Trp Trp STOP AUA Ile Met Met Ile CUA Leu Leu Thr Leu AGA Arg STOP Ser Arg AGG Arg STOP Ser Arg
 Расшифровка кода Ø В 1961 г. М. Ниренберг и Г. Маттеи впервые расшифровали триплет УУУ, являющийся кодоном для фенилаланина. Синтезировали РНК-гомополимеры СССССС……. , UUUUUU(полиуридиловая GGGGGG…… – ААААААА……, кислота)…… или Прямой метод анализа, основанный на синтезе полипептидной цепочке in vitro на матрице синтетической РНК с известным соотношением случайно расположенных нуклеотидов. Ø Полностью расшифровка генетического кода была завершена в 1966 г. Гобинд Корана (химик-органик) осуществил синтез всех 64 триплетов и оказалось возможным определить значение всех триплетов генетического кода.
Расшифровка кода Ø В 1961 г. М. Ниренберг и Г. Маттеи впервые расшифровали триплет УУУ, являющийся кодоном для фенилаланина. Синтезировали РНК-гомополимеры СССССС……. , UUUUUU(полиуридиловая GGGGGG…… – ААААААА……, кислота)…… или Прямой метод анализа, основанный на синтезе полипептидной цепочке in vitro на матрице синтетической РНК с известным соотношением случайно расположенных нуклеотидов. Ø Полностью расшифровка генетического кода была завершена в 1966 г. Гобинд Корана (химик-органик) осуществил синтез всех 64 триплетов и оказалось возможным определить значение всех триплетов генетического кода.