К.У.Р.С..pptx
- Количество слайдов: 52



Молекулярные основы биологии развития Экспрессия генов

Дифференциальная экспрессия генов Две клетки дифференцированы поразному, если, обладая одинаковым геномом, они синтезируют разные белки. Φ. ЖАКОБ и Ж. МОНО (1963)

Лекции Р. П. Костюченко Дифференциальная экспрессия генов 1960 -е 1. Каждое ядро соматической клетки содержит полный геном, возникающий при оплодотворении яйцеклетки. Это означает, что ДНК во всех дифференцированных клетках идентична. 2. Неиспользуемые гены в дифференцированных клетках не подвергаются разрушению или мутациям, они сохраняют способность к экспрессии. 3. Только небольшой процент генома экспрессируется в каждой клетке, часть РНК, синтезируемой в клетке, специфична для данного типа клеток. Экспрессия гена - реализация генетической информации, закодированной в гене

Механизмы дифференциальной экспрессии генов. Эпигенетический ландшафт Уоддингтона. Шарик на вершине изображает клетку, а долины под ним - различные пути развития, по которым она может пойти.

Регуляция экспрессии генов ядро цитоплазма Контроль деградации м. РНК ДНК Первичный транскрипт м. РНК Контроль транскрипции м. РНК Процессинг РНК м. РНК Контроль транспорта м. РНК Контроль деградации белка Деградация белка Контроль трансляции м. РНК Деградация м. РНК Неактив ный белок Контроль ферментативной активности белка Активный белок

Нуклеосома - базовая единица хроматиновой структуры гистоны (две молекулы каждого из гистонов H 2 A-H 2 B и гистонов H 3 -H 4), обернутых двумя витками ДНК. Динамичная структура, сворачивается/разворачивается около 4 раз в секунду. Модификации гистонов: «гистоновый код» ацетилирование гистонов - активирует транскрипцию ( «разрыхляя» хроматин) деацетилирование гистонов – Инактивирует метилирование гистонов – при метилировании по «хвостам» H 3, Н 4 – уплотнение хроматина, умолкание генов, гетерохроматинизация. хроматин - комплекс ДНК с белком

Считывание гистонового кода. Комплекс считывания кода связывается специфически свяжется только с областью хроматина, содержащей распознаваемые им метки, так что только определенная комбинация меток вызовет связывание комплекса с хроматином и привлечет дополнительные белковые комплексы, которые катализируют одну или несколько биологических функций.

• Мозаичный эффект положения, проявляющийся в окраске глаз дрозофилы: заглушение гетерохроматином прилежащих генов.

Строение гена эукариот и процесс реализации генетической информации

Факторы транскрипции — белки, контролирующие процесс синтеза м. РНК на матрице ДНК путём связывания со специфичными участками ДНК. Транскрипционные факторы выполняют свою функцию либо самостоятельно, либо в комплексе с другими белками. Они обеспечивают снижение (репрессоры) или повышение (активаторы) константы связывания РНК-полимеразы с регуляторными последовательностями регулируемого гена. Определяющая черта факторов транскрипции — наличие в их составе одного или более ДНК-связывающих доменов, которые взаимодействуют с характерными участками ДНК, расположенными в регуляторных областях генов.

• Конститутивные ТФ — присутствуют всегда во всех клетках — главные факторы транскрипции. • Активируемые ТФ (активны в определенных условиях) – Участвующие в развитии организма (клеткоспецифичные) — экспрессия строго контролируется, но, начав экспрессироваться, не требуют дополнительной активации —, Myo. D, Myf 5, Hox. • Сигнал-зависимые — требуют внешнего сигнала для активации - внеклеточные сигнал-зависимые - внутриклеточные сигнал-зависимые - мембраносвязанные рецептор-зависимые — фосфорилируются киназами сигнального каскада ДНК-связывающий домен типа «лейциновая молния» в комплексе с ДНК. • резидентные ядерные факторы — находятся в ядре независимо от активации — CREB, AP-1, Mef 2 • латентные цитоплазматические факторы — в неактивном состоянии локализованы в цитоплазме, после активации транспортируются в ядро — STAT, R-SMAD, NF-k. B, Notch,

• Сборка комплекса инициации транскрипции у эукариот на последовательности ТАТА. Структура комплекса TATAсвязывающего белка/транскрипционного фактора TF(II)B из археи Pyrococcus woesei с ДНК
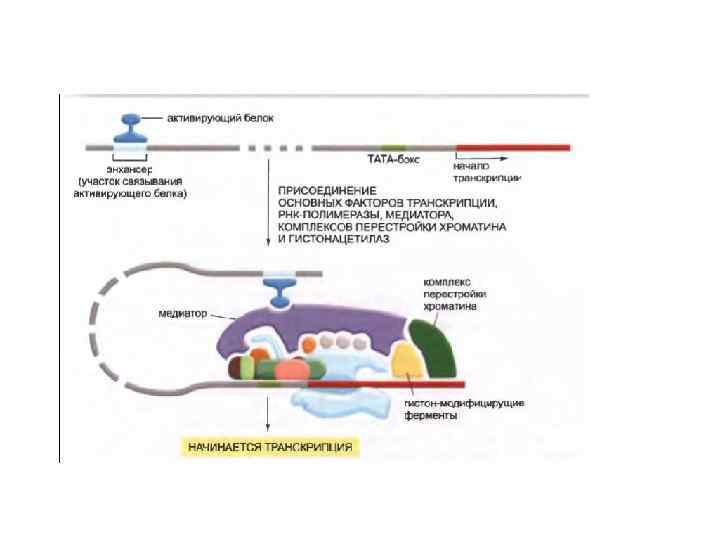

Регуляторные белки эукариот собираются в комплексы на ДНК. Природа и функция такого комплекса зависит от специфической последовательности ДНК, которая служит затравкой для их сборки. Белки, которые не связываются самостоятельно с ДНК, но собираются на других связывающихся с ДНК регуляторных белках, часто называются коактиваторами или корепрессорами (кофакторами) транскрипции. Под этим термином могут пониматься комплексы перестройки хроматина (напр. гистонацетилазы), белки, усиливающие сродство полимеразного комплекса к ДНК, или же просто белки-”строительные леса”, служащие основой для прикрепления обладающих специфической активностью белков.

Объединение множества входящих сигналов на промоторе. Чтобы воздействовать на инициацию транскрипции на промоторе, многочисленные белковые комплексы работают сообща. Конечная транскрипционная активность гена является результатом конкуренции между активаторами и репрессорами.

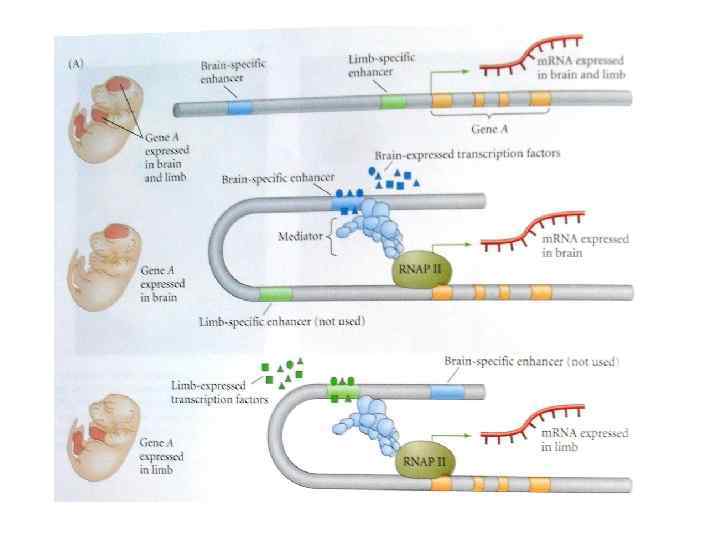
Энхансеры ДНК-последовательности, которая селективно повышают активность промотора, контролируя частоту осуществляющейся с него инициации транскрипции. Связывают транскрипционные (ко)факторы, которые способны увеличивать уровень транскрипции.

Экпрессия Pax-6 Энхансеры могут контролировать временную и тканеспецифическую экспрессию любого дифференциально регулируемого гена, так что различные типы генов имеют как правило различные энхансеры

Лекции Р. П. Костюченко Сайленсеры Районы ДНК, которые отвечают за репрессию транскрипции какого-либо гена. (Посредством привлечения белков с соответствующей активностью) Нейроспецифический сайленсерный элемент (neural restrictive silencer element) — NRSE; найден в регуляторных районах нескольких мышиных генов, экспрессия которых ограничена нервной системой Нейроспецифический сайленсерный фактор (neural restrictive silencer factor) — NRSF , по- видимому, синтезируется в каждой клетке организма, не являющейся зрелым нейроном по Гилберт, 2010
 на соседние гены")
Лекции Р. П. Костюченко Инсуляторы Чтобы предотвратить распространение влияние энхансера (сайленсера) на соседние гены существуют определенные участки ДНК, которые связывают белки, блокирующие действие регуляторного элемента на соседний промотор. по Гилберт, 2010
 Связывание активатора с лигандом стимулирует сборку")
Возможные варианты регуляции инициации транскрипции у прокариот (а) Связывание активатора с лигандом стимулирует сборку комплекса и транскрипцию (б) Активатор стимулирует транскрипцию, при связывании с лигандом дезактивируется. (в)Репрессор запрещает транскрипцию. Взаимодействие с лигандом инактивирует репрессор и позволяет транскрипцию. (г) В отсутствие лиганда репрессор не способен взаимодействов ать с ДНК, репрессия происходит только в присутствие лиганда 21

Лекция вторая. Возникновение упорядоченности. Материнские факторы. Сигнальные пути клетки.

Запасение материнских белков.

Запасение материнских м. РНК

Cell signalling

Клеточные сигнальные пути Каскады межмолекулярных взаимодействий, обеспечивающие такую коммуникацию между клеточной мембраной и внутриклеточной точкой приложения, что способна привести к некоторым изменениям в клетке.
 Изменение цитоскелета (изменение")
Точки приложения сигнальных каскадов Регуляция экспрессии генов (пролиферация, дифференцировка, выполнение функций) Изменение цитоскелета (изменение формы клетки, миграция, установление/разборка клеточных контактов) Влияние на метаболические пути (секреция метаболитов, регуляция активности ферментов) Не обязательно вовлекаются ДНК/РНК.

Способы передачи сигналов от клетки к клетке: Через воздействие паракринных факторов, взаимодействие клеток с внеклеточным матриксом, через межклеточные контакты Диффузия растворимых сигнальных факторов Внеклеточный матрикс, секретируемый одной клеткой, вызывает изменения в другой Контакт между индуцирующей и отвечающей клетками

Источник сигнала Клетка-источник сигнала секретирует определённый тип сигнальной молекулы. Эта молекула детектируется клеткой-мишенью с помощью белка -рецептора, распознающего её и специфически с ней взаимодействующего. Каждая клетка способна отзываться на ограниченный набор сигнальных молекул. Реакция клетки на сигнал зависит от её состояния и типа дифференцировки.

Интеграция сигнала Сигналы из разных источников могут сходиться на The signals from several different sources may be integrated though a single shared protein (A) or protein complex (B) общем белке или белковом комплексе.

Амплификация сигнала 1 рецептор активирует множество G-белков 1 ligand-receptor 500 G-protein 500 enzymes Each enzyme Y produces many second messangers, each messanger activates 1 enzyme Y 105 (2 nd messanger) 250 (ion channels) 105 -107 (ions)

Ветвление сигнальных путей

Сигнальные модули не консервативны в широких эволюционных масштабах.

Независимая эволюция ядерных рецепторов в разных группах эукариот
 С другой стороны, структура белковучастников зачастую высококонсерватина.")
Ле Суперсемейство ростовых факторов TGF-β (TGF-β -суперсемейство) С другой стороны, структура белковучастников зачастую высококонсерватина. Зачастую белки, экспрессируемые в неродственном организме, способны функционально замещать гомологичные им белки хозяина. по Гилберт, 2010

Адапторные белки: доменная организация
 ◦ Ковалентные модификации:")
Молекулярные взаимодействия Белок-белковые взаимодействия: ◦ Присоединение/диссоциация (Создание или разрушение белковых комплексов) ◦ Ковалентные модификации: фосфорилирование (tyr, thr, ser) ◦ Конформационные изменения ◦ Перемещение в другую функциональную область клетки ◦ Убиквитинирование и деградация Взаимодействие белков с малыми молекулами ◦ Присоединение/диссоциация, ведущая к изменению конформации, энергетического состояния ◦ Распространение вторичных мессенджеров ( Ca 2+, ц. АМФ)
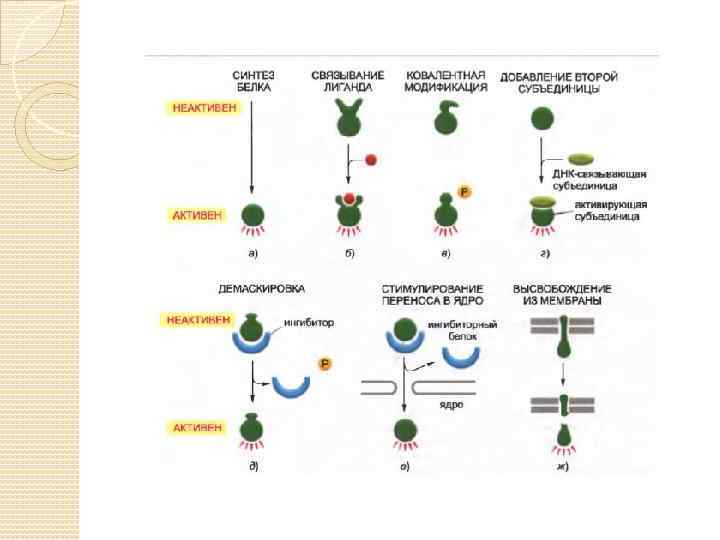

Мерой силы связывания служит константа равновесия

Фосфорилирование белков • Привнесение двух отрицательных фосфатных зарядов может вызвать значительное конформационное изменение в белке за счёт, например, притяжения группы положительно заряженных боковых цепей аминокислот. Это может, в свою очередь, повлиять на связывание лигандов и тем самым заметно изменить активность фосфорилированного белка по сравнению с исходным.

Фосфорилирование белков • Привнесённая фосфатная группа может образовать часть структуры, которую опознают участки связывания других белков.

Распознавание p-Tyr SH 2 -доменом

Убиквитинирование и деградация в протеасомах.

Убиквитинирование и деградация в протеасомах.

Принципиальная схема влияния полученного сигнала на дифференциальную экспрессию генов Внешняя среда ядро цитоплазма Активный белок (1) Коактиваторы транскрипции Р Р Факторы транскрипции Перенос Активный белок (2) Лиганд Белок (2) Белковый комплекс Корепрессоры транскрипции Белок(2) n раз Деградация белка Белокрецептор

Сигнальный путь Wnt
 семейство гликопротеинов,")
Лекции Р. П. Костюченко ПАРАКРИННЫЕ ФАКТОРЫ: сигнальный путь Wnt Семейство Wingless (Wnt-семейство) семейство гликопротеинов, богатых цистеином индуцируют дорсальные клетки сомитов становиться мышечными. участвуют в спецификации клеток среднего мозга Белки Wnt важны для становления полярности конечностей насекомых и позвоночных; они также участвуют в развитии (на различных этапах) мочеполовой системы Б — мочеполовой зачаток новорожденной самки мыши дикого типа. В — Мочеполовой зачаток самки мыши, нокаутированной по гену Wnt 4, с дефектом развития почки. Кроме того, яичник начинает синтезировать тестостерон и окружается системой протоков мужского типа. Фото J. Perasaari, S. Vainio
 сигналинг: Сигнальный путь Notch. мембраносвязанные лиганды и рецепторы")
Лекции Р. П. Костюченко Юкстакринный (контактный) сигналинг: Сигнальный путь Notch. мембраносвязанные лиганды и рецепторы по Гилберт, 2010

Лекции Р. П. Костюченко Юкстакринный сигналинг • первоначальные различия между клетками возникают случайно • эти различия закрепляются по принципу обратной связи

Лекции Р. П. Костюченко Модель создания пространственной структуры нейробластов из исходно равноценных клеток нейрогенной эктодермы. Нейрогенные клетки производят сигнал в виде белка Delta (темная штриховка), а клетки, не становящиеся нейрогенными, продуцируют рецепторный белок Notch (белые)
- и Hh (hedgehog) - путей сигналинга у Drosophila")
Взаимообразные обмены сигналами Wnt (wingless)- и Hh (hedgehog) - путей сигналинга у Drosophila

Major themes in ST The “internal complexity” of each interaction The combinatorial nature of each component molecule (may receive and send multiple signals) The integration of pathways and networks
К.У.Р.С..pptx