

Механизмы мутагенеза репарации.ppt
- Количество слайдов: 12
 Молекулярные механизмы мутагенеза и репарации А. К. Гришанин
Молекулярные механизмы мутагенеза и репарации А. К. Гришанин
. Мутации- это спонтанные или индуцированные изменения генетического материала. Основные типы точковых мутаций:") Mutatio (изменение). Мутации- это спонтанные или индуцированные изменения генетического материала. Основные типы точковых мутаций: а) замена оснований и б) мутации со сдвигом рамки считывания. Транзиции: Замены пурина на пурин A→G; G→A; Пиримидина на пиримидин С→ Т; Т→С Азотистая к-та + аденин = урацил (C-G) C→U→ U-A (T-A) Трансверсии – замена пурина на пиримидин или пиримидина на пурин A→T; A→C; G→T; С→ A; Т→A; С→ G; Т→G Замены оснований ведут к появлению миссенс-мутаций ( с измененным смыслом), что приводит к изменению триплета и замены аминокислоты (так код вырожденный, то иногда замены аминокислоты не происходит- нейтральные мутации); и нонсенс ( бессмысленных) мутаций, когда замена оснований превращает кодон в нонсенс-кодон, останавливающий трансляцию.
Mutatio (изменение). Мутации- это спонтанные или индуцированные изменения генетического материала. Основные типы точковых мутаций: а) замена оснований и б) мутации со сдвигом рамки считывания. Транзиции: Замены пурина на пурин A→G; G→A; Пиримидина на пиримидин С→ Т; Т→С Азотистая к-та + аденин = урацил (C-G) C→U→ U-A (T-A) Трансверсии – замена пурина на пиримидин или пиримидина на пурин A→T; A→C; G→T; С→ A; Т→A; С→ G; Т→G Замены оснований ведут к появлению миссенс-мутаций ( с измененным смыслом), что приводит к изменению триплета и замены аминокислоты (так код вырожденный, то иногда замены аминокислоты не происходит- нейтральные мутации); и нонсенс ( бессмысленных) мутаций, когда замена оснований превращает кодон в нонсенс-кодон, останавливающий трансляцию.
 Мутации со сдвигом рамки считывания- frame shift-обусловлены выпадением или вставкой одного или нескольких нуклеотидов. Частота мутаций в «Горячих точках» (Hotspots) в 10 -100 раз может превышать частоту мутаций в других сайтах. Мобильные элементы перемещаясь приводят к инсерциям или делециям. Экспансия тринуклеотидных повторовв интронах (CGG) и в экзонах (CAG). Мутации гена дикого типа называют прямыми. Мутации восстанавливающие дикий фенотип- обратные мутации или реверсии. Супрессорная мутация- компенсаторная мутация: а) замена мутантного кодона внутри гена, б) вне гена, изменение в структуре т-РНК. Причины мутирования. Спонтанные мутации: ошибки репликации, изменения в химических составляющих молекулы ДНК (кето и аминоформы пуринов и пиримидинов приводят АС, G-T и далее к транзициям и трансверсиям). Гены – мутаторы увеличивают мутабильность в 100 -1000 раз. Ионизирующее излучение. УФ. Интеркалирующие агенты. Сайт-пецифический мутагенез.
Мутации со сдвигом рамки считывания- frame shift-обусловлены выпадением или вставкой одного или нескольких нуклеотидов. Частота мутаций в «Горячих точках» (Hotspots) в 10 -100 раз может превышать частоту мутаций в других сайтах. Мобильные элементы перемещаясь приводят к инсерциям или делециям. Экспансия тринуклеотидных повторовв интронах (CGG) и в экзонах (CAG). Мутации гена дикого типа называют прямыми. Мутации восстанавливающие дикий фенотип- обратные мутации или реверсии. Супрессорная мутация- компенсаторная мутация: а) замена мутантного кодона внутри гена, б) вне гена, изменение в структуре т-РНК. Причины мутирования. Спонтанные мутации: ошибки репликации, изменения в химических составляющих молекулы ДНК (кето и аминоформы пуринов и пиримидинов приводят АС, G-T и далее к транзициям и трансверсиям). Гены – мутаторы увеличивают мутабильность в 100 -1000 раз. Ионизирующее излучение. УФ. Интеркалирующие агенты. Сайт-пецифический мутагенез.
 вот лес вяз дуб ивы тут был пал дым шел три дня (вставка а ) вот лес авя зду бив ыту тбы лпа лды мше лтр идн я (вторичная мутация в результате делеции з) вот лес авя дуб ивы тут был пал дым шел три дня
вот лес вяз дуб ивы тут был пал дым шел три дня (вставка а ) вот лес авя зду бив ыту тбы лпа лды мше лтр идн я (вторичная мутация в результате делеции з) вот лес авя дуб ивы тут был пал дым шел три дня
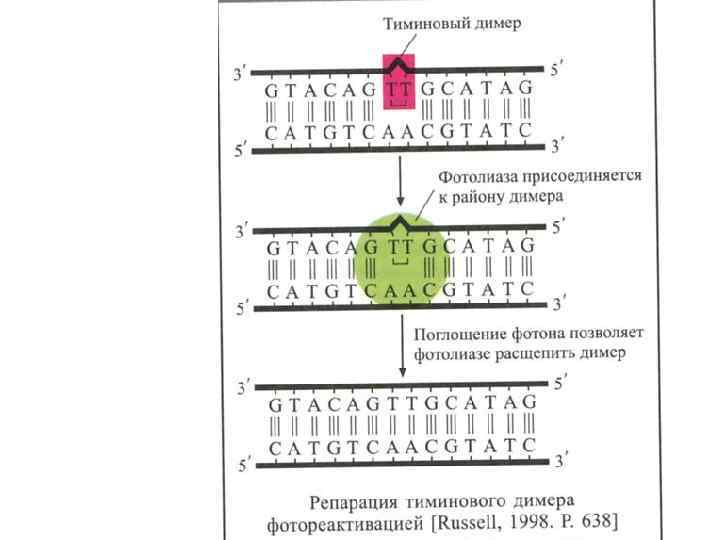
 Механизмы репарации ДНК Прямая коррекция в бактериальных геномах: ДНК-полимераза, которая осуществляет синтез цепи 5′-3′, проверяет цепь 3′-5′. Фотореактивация была обнаружена в 1949 г. Кельнером, Дюльбеко и Ковалевым, в ходе Которой облучение видимым светом позволяет фотолиазе расщеплять тиминовые димеры. Репарация алкилирующих повреждений заключается в удалении алкильных или метильных групп нуклеотида. Сам нуклеотид не удаляется. ДНК-полинуклеотидлигаза воссоединяет однонитевые концы ДНК вследствие разрывов ДНК.
Механизмы репарации ДНК Прямая коррекция в бактериальных геномах: ДНК-полимераза, которая осуществляет синтез цепи 5′-3′, проверяет цепь 3′-5′. Фотореактивация была обнаружена в 1949 г. Кельнером, Дюльбеко и Ковалевым, в ходе Которой облучение видимым светом позволяет фотолиазе расщеплять тиминовые димеры. Репарация алкилирующих повреждений заключается в удалении алкильных или метильных групп нуклеотида. Сам нуклеотид не удаляется. ДНК-полинуклеотидлигаза воссоединяет однонитевые концы ДНК вследствие разрывов ДНК.
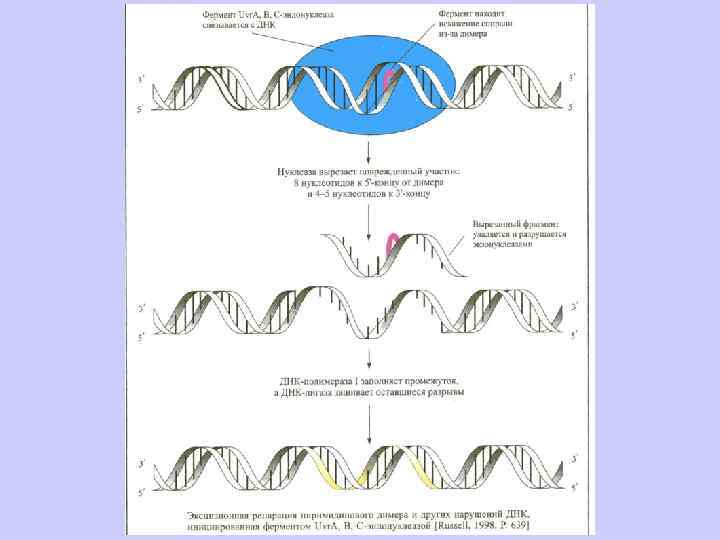
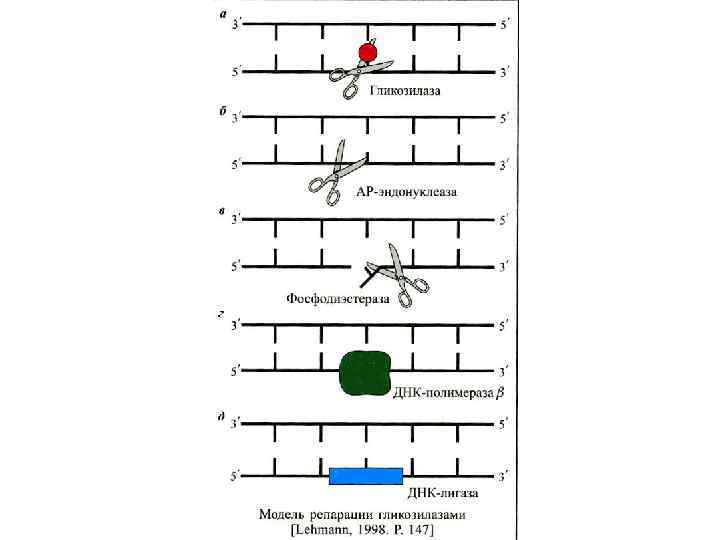
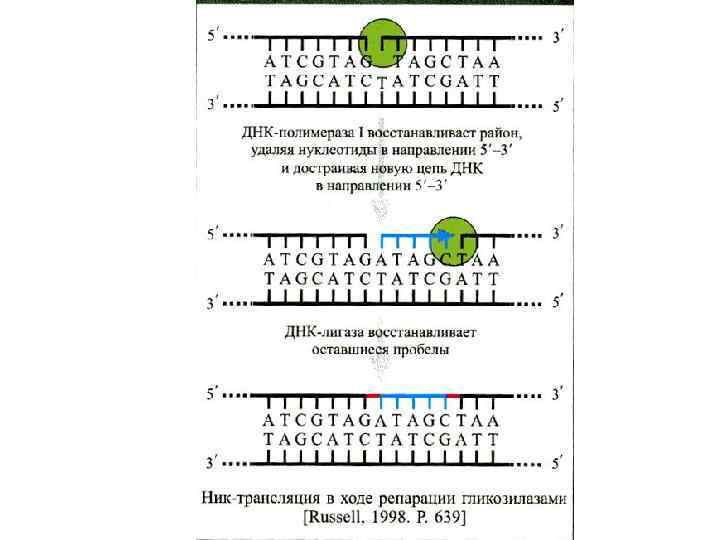
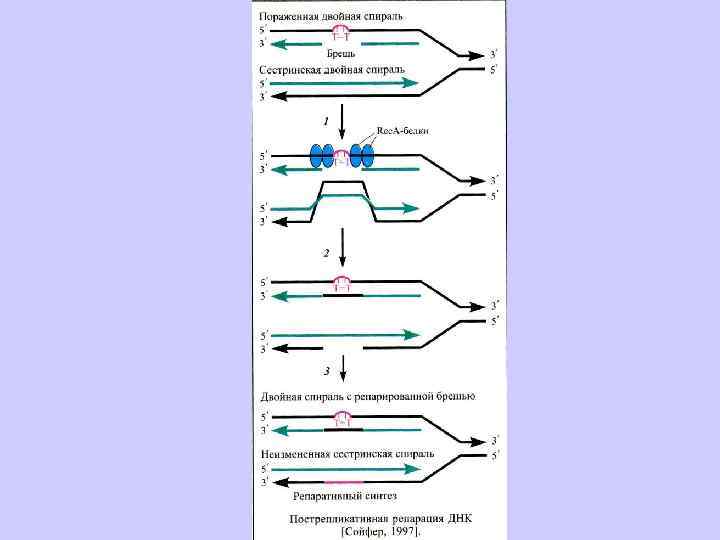
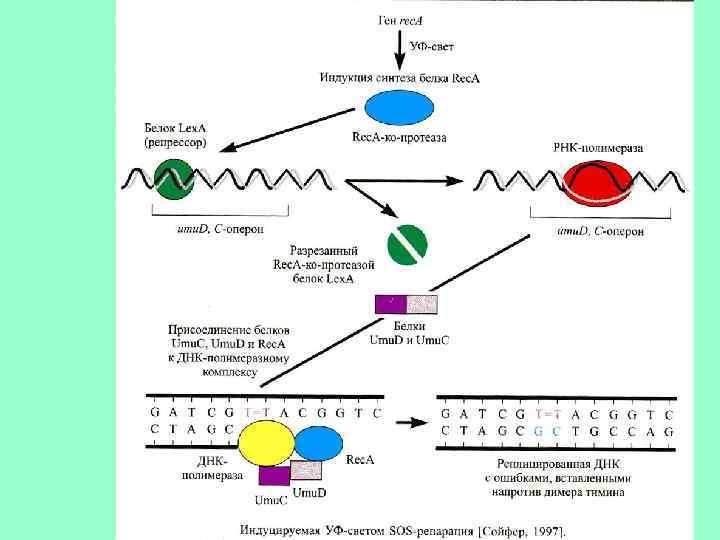
 Эксцизионная репарация была открыта в 1964 г. группами Р. Бойса, П. Говарда-Флендерса, Р. Сетлоу и Кэрриера и наблюдается у большинства изученных организмов. Поврежденные азотистые основания (дезаминированные, окисленные) репарируются гликозилазами. НИК-трансляция – это активность ДНК-полимеразы-I, в ходе которой фермент удаляет нуклеотиды впереди растущей цепи ДНК. Пострепликативная или рекомбинационная репараци(ППР) осуществляется путем рекомбинационных обменов между двумя сестринскими молекулами ДНК, и позволяет репарировать пробелы в дочерней цепи ДНК напротив димеров пиримидина. ППР контролируется не менее, чем 17 генами. SOS- репарация включается в случае, если другие системы репарации не устранили повреждение. ДНК-полимеразный комплекс и специально синтезируемые белки достраивают нить ДНК напротив дефектных
Эксцизионная репарация была открыта в 1964 г. группами Р. Бойса, П. Говарда-Флендерса, Р. Сетлоу и Кэрриера и наблюдается у большинства изученных организмов. Поврежденные азотистые основания (дезаминированные, окисленные) репарируются гликозилазами. НИК-трансляция – это активность ДНК-полимеразы-I, в ходе которой фермент удаляет нуклеотиды впереди растущей цепи ДНК. Пострепликативная или рекомбинационная репараци(ППР) осуществляется путем рекомбинационных обменов между двумя сестринскими молекулами ДНК, и позволяет репарировать пробелы в дочерней цепи ДНК напротив димеров пиримидина. ППР контролируется не менее, чем 17 генами. SOS- репарация включается в случае, если другие системы репарации не устранили повреждение. ДНК-полимеразный комплекс и специально синтезируемые белки достраивают нить ДНК напротив дефектных