

ОМФ 11 Гормоны.ppt
- Количество слайдов: 39
 Молекулярные аспекты эндокринной регуляции Дружинин В. Ю. , ассистент кафедры нормальной физиологии.
Молекулярные аспекты эндокринной регуляции Дружинин В. Ю. , ассистент кафедры нормальной физиологии.
 Глюкагон • 29 АК остатков. • Секретируется α-клетками поджелудочной • • железы. Рецептор глюкагона (РГ) клонирован в 1993 году, располагается 17 q 25. РГ – член GPCR. Запускает ц. АМФ-зависимый путь и фосфолипазный механизм; ПК А катализирует фосфорилирование сериновых и треониновых остатков на регуляторных участках ключевых ферментов;
Глюкагон • 29 АК остатков. • Секретируется α-клетками поджелудочной • • железы. Рецептор глюкагона (РГ) клонирован в 1993 году, располагается 17 q 25. РГ – член GPCR. Запускает ц. АМФ-зависимый путь и фосфолипазный механизм; ПК А катализирует фосфорилирование сериновых и треониновых остатков на регуляторных участках ключевых ферментов;
 • В дополнение к быстрым эффектам, ПК А фосфорилирует CREB – ядерный фактор транскрипции, связывающийся непосредственно с участком ДНК (c. AMPresponse element), изменяя экспрессию гена. • Множество печёночных эффектов глюкагона могут быть также стимулированы активацией β-АР→ц. АМФ.
• В дополнение к быстрым эффектам, ПК А фосфорилирует CREB – ядерный фактор транскрипции, связывающийся непосредственно с участком ДНК (c. AMPresponse element), изменяя экспрессию гена. • Множество печёночных эффектов глюкагона могут быть также стимулированы активацией β-АР→ц. АМФ.
 • РГ → Gq-белок → ДАГ → ПК С. • Фосфолипазный и ц. АМФ-зависимый путь синергичны.
• РГ → Gq-белок → ДАГ → ПК С. • Фосфолипазный и ц. АМФ-зависимый путь синергичны.
 Печёночные эффекты Печень – главная мишень глюкагона! Глюкагон повышает образование глюкозы и выброс её в кровь. Глюкагон стимулирует распад гликогена и тормозит его синтез, взаимодействуя через посредников с ключевыми ферментами, контролирующими этот процесс.
Печёночные эффекты Печень – главная мишень глюкагона! Глюкагон повышает образование глюкозы и выброс её в кровь. Глюкагон стимулирует распад гликогена и тормозит его синтез, взаимодействуя через посредников с ключевыми ферментами, контролирующими этот процесс.
 активирует её. • Это обеспечивается") Гликогенолиз • Фосфорилирование серин-14 -участка гликоген-фосфорилазы (100 к. Да) активирует её. • Это обеспечивается работой киназы фосфорилазы (1300 к. Да), которая фосфорилируется по δ-СЕ ПК С или по α- или β-СЕ посредством ПК А. • Действие глюкагона лимитируется работой фосфатаз (PP-1 Gs), которые переводят гликоген-фосфорилазу в неактивную форму
Гликогенолиз • Фосфорилирование серин-14 -участка гликоген-фосфорилазы (100 к. Да) активирует её. • Это обеспечивается работой киназы фосфорилазы (1300 к. Да), которая фосфорилируется по δ-СЕ ПК С или по α- или β-СЕ посредством ПК А. • Действие глюкагона лимитируется работой фосфатаз (PP-1 Gs), которые переводят гликоген-фосфорилазу в неактивную форму
,") Гликогенолиз • Торможение синтеза гликогена происходит вследствие фосфорилирования сериновых остатков гликоген-синтазы (85 к. Да), опосредованное ПК А.
Гликогенолиз • Торможение синтеза гликогена происходит вследствие фосфорилирования сериновых остатков гликоген-синтазы (85 к. Да), опосредованное ПК А.
 Гликолиз/Глюконеогенез • Глюкагон тормозит гликолиз и стимулирует глюконеогенез. • Метаболический баланс между распадом и образованием глюкозы зависит от баланса между активностью фосфофрукто-1 киназой (PFK-1) и фруктозо-1, 6 бисфосфатазой. • Через ПК А глюкагон стимулирует фосфорилирование PFK-2 (фосфофрукто 2 -киназа), снижая гликолиз. • Ингибируя PFK-1, глюкагон стимулирует глюконеогенез.
Гликолиз/Глюконеогенез • Глюкагон тормозит гликолиз и стимулирует глюконеогенез. • Метаболический баланс между распадом и образованием глюкозы зависит от баланса между активностью фосфофрукто-1 киназой (PFK-1) и фруктозо-1, 6 бисфосфатазой. • Через ПК А глюкагон стимулирует фосфорилирование PFK-2 (фосфофрукто 2 -киназа), снижая гликолиз. • Ингибируя PFK-1, глюкагон стимулирует глюконеогенез.
 Глюкагон также изменяется экспрессию генов посредством CREB, повышая уровень ферментов глюконеогенеза – фосфоенол-пируват-карбоксикиназы, фруктозо-1, 6 -бисфосфатазы, глюкозо-6 -фосфатазы. Глюкагон уменьшает уровень GLUT-2 и глюкокиназы, уменьшая захват глюкозы печенью.
Глюкагон также изменяется экспрессию генов посредством CREB, повышая уровень ферментов глюконеогенеза – фосфоенол-пируват-карбоксикиназы, фруктозо-1, 6 -бисфосфатазы, глюкозо-6 -фосфатазы. Глюкагон уменьшает уровень GLUT-2 и глюкокиназы, уменьшая захват глюкозы печенью.
 Глюкагон индуцирует образование кетонов и тормозит синтез жиров в печени. Синтез липидов ингибируется посредством фосфорилирования ацетил-ко. Акарбоксилазы. Этот эффект усиливается снижением гликолиза, что приводит к уменьшению ацетил-ко. А
Глюкагон индуцирует образование кетонов и тормозит синтез жиров в печени. Синтез липидов ингибируется посредством фосфорилирования ацетил-ко. Акарбоксилазы. Этот эффект усиливается снижением гликолиза, что приводит к уменьшению ацетил-ко. А
 Роль глюкагона • В сверхвысоких дозах глюкагон оказывает липолитическое действие на жировую ткань. • Глюкагон потенциирует секрецию инсулина β-клетками, действуя в паракринной манере, хотя и в меньшей степени, чем глюкагон-подобный пептид -1.
Роль глюкагона • В сверхвысоких дозах глюкагон оказывает липолитическое действие на жировую ткань. • Глюкагон потенциирует секрецию инсулина β-клетками, действуя в паракринной манере, хотя и в меньшей степени, чем глюкагон-подобный пептид -1.
 Роль глюкагона • Оказывает положительное инотропное и хронотропное действие на сердце, является вазодилататором. • В почках глюкагон увеличивает почечный кровоток, СКФ, выведение электролитов. • Несмотря на большую роль почек в глюконеогенезе, глюкагон не оказывает влияния на продукцию глюкозы в почках.
Роль глюкагона • Оказывает положительное инотропное и хронотропное действие на сердце, является вазодилататором. • В почках глюкагон увеличивает почечный кровоток, СКФ, выведение электролитов. • Несмотря на большую роль почек в глюконеогенезе, глюкагон не оказывает влияния на продукцию глюкозы в почках.
 При СД 1 типа повышается уровень циркулирующего глюкагона, его") Аномальное действие глюкагона • 1) При СД 1 типа повышается уровень циркулирующего глюкагона, его секреция не подавляется приёмом пищи. Это наводит на мысль, что СД 1 – бигормональная болезнь, а гиперглюкагонемия вносит вклад в гипергликемию. • С другой стороны может страдать выброс глюкагона в ответ на гипогликемию, что значительно повышает риск тяжёлых эпизодов гипогликемии.
Аномальное действие глюкагона • 1) При СД 1 типа повышается уровень циркулирующего глюкагона, его секреция не подавляется приёмом пищи. Это наводит на мысль, что СД 1 – бигормональная болезнь, а гиперглюкагонемия вносит вклад в гипергликемию. • С другой стороны может страдать выброс глюкагона в ответ на гипогликемию, что значительно повышает риск тяжёлых эпизодов гипогликемии.
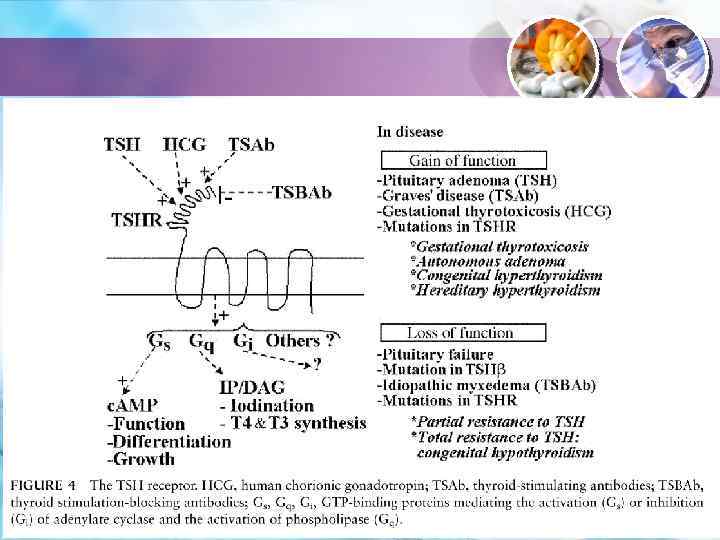
 ТТГ • • ТТГ оказывает два типа эффектов на ЩЖ. Когда назначается остро, в больших дозах – в течение минут и часов стимулирует синтез и секрецию гормонов. При постоянном высоком уровне ТТГ контролирует рост и пролиферацию тироцитов. Тиротропин (МВ 7000) – гетеродимер α-СЕ, общей для ФСГ и ЛГ + β-СЕ, специфичной для ТТГ. Синтезируется и секретируется гипофизарными тиротрофами. Распознается особым рецептором, находящимся в тироцитах.
ТТГ • • ТТГ оказывает два типа эффектов на ЩЖ. Когда назначается остро, в больших дозах – в течение минут и часов стимулирует синтез и секрецию гормонов. При постоянном высоком уровне ТТГ контролирует рост и пролиферацию тироцитов. Тиротропин (МВ 7000) – гетеродимер α-СЕ, общей для ФСГ и ЛГ + β-СЕ, специфичной для ТТГ. Синтезируется и секретируется гипофизарными тиротрофами. Распознается особым рецептором, находящимся в тироцитах.
 ТТГ • Рецептор ТТГ – классический GPCR. Он активирует три G-белка: Gs, Gq и Gi, что приводит к образованию/снижению. ц. АМФ, ИТФ и ДАГ. У человека преимущественно активирует ц. АМФ-зависимый путь и фосфолипазный механизм. • ц. АМФ активирует два эффекторных протеина: • ПК А и EPAC-протеин (обменник гуаниновых нуклеотидов, активирующий G-белки Rap 1 и Rap 2).
ТТГ • Рецептор ТТГ – классический GPCR. Он активирует три G-белка: Gs, Gq и Gi, что приводит к образованию/снижению. ц. АМФ, ИТФ и ДАГ. У человека преимущественно активирует ц. АМФ-зависимый путь и фосфолипазный механизм. • ц. АМФ активирует два эффекторных протеина: • ПК А и EPAC-протеин (обменник гуаниновых нуклеотидов, активирующий G-белки Rap 1 и Rap 2).
 ТТГ активирует ФЛ С → ДАГ + ИТФ → ПК С. Этот путь требует больших доз ТТГ, чем стимуляция АЦ. Активация ПК С приводит к: 1) Стимуляция образования Н 2 О 2; 2) Йодирование протеина; 3) Синтез гормонов ЩЖ. Са-кальмодулин вызывает те же эффекты.
ТТГ активирует ФЛ С → ДАГ + ИТФ → ПК С. Этот путь требует больших доз ТТГ, чем стимуляция АЦ. Активация ПК С приводит к: 1) Стимуляция образования Н 2 О 2; 2) Йодирование протеина; 3) Синтез гормонов ЩЖ. Са-кальмодулин вызывает те же эффекты.
 • ТТГ стимулирует экспрессию генов, связанных со специфическими тироидными белками, посредством фосфорилирования ПК-й А различных факторов транскрипции (CREB, CREM и др. )
• ТТГ стимулирует экспрессию генов, связанных со специфическими тироидными белками, посредством фосфорилирования ПК-й А различных факторов транскрипции (CREB, CREM и др. )
 Захвата йода") Синтез гормонов ЩЖ • Синтез гормонов на верхушке тироцита требует: • 1) Захвата йода из внеклеточной жидкости посредством Na/I-симпорта (NIS) на основании клетки. • 2) На верхушке клетки – транспорт йодида из клетки в полость фолликула посредством специализированных каналов. • 3) Синтез тироглобулина (МВ 680 000), который является матрицей для синтеза тироидных гормонов.
Синтез гормонов ЩЖ • Синтез гормонов на верхушке тироцита требует: • 1) Захвата йода из внеклеточной жидкости посредством Na/I-симпорта (NIS) на основании клетки. • 2) На верхушке клетки – транспорт йодида из клетки в полость фолликула посредством специализированных каналов. • 3) Синтез тироглобулина (МВ 680 000), который является матрицей для синтеза тироидных гормонов.
 Транспорт тироглобулина в полость фолликула посредством экзоцитоза. 5) Образование Н") Синтез гормонов ЩЖ 4) Транспорт тироглобулина в полость фолликула посредством экзоцитоза. 5) Образование Н 2 О 2 посредством ТНОХ (тироидной Н 2 О 2 -генерирующей системы). 6) Окисление йодида и его ковалентное связывание с тирозиновыми остатками тироглобулина посредством тиропероксидазы, при участии Н 2 О 2. 7) Спаривание йодотиронинов посредством тиропероксидазы.
Синтез гормонов ЩЖ 4) Транспорт тироглобулина в полость фолликула посредством экзоцитоза. 5) Образование Н 2 О 2 посредством ТНОХ (тироидной Н 2 О 2 -генерирующей системы). 6) Окисление йодида и его ковалентное связывание с тирозиновыми остатками тироглобулина посредством тиропероксидазы, при участии Н 2 О 2. 7) Спаривание йодотиронинов посредством тиропероксидазы.
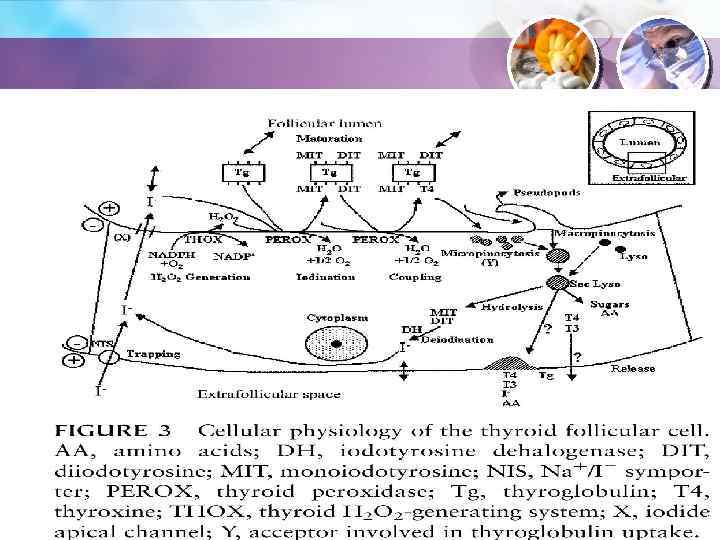
 Перенос йодида в полость фолликула; 2) Генерация Н 2 О") Действие ТТГ стимулирует: 1) Перенос йодида в полость фолликула; 2) Генерация Н 2 О 2; 3) Йодирование тироглобулина; 4) Спаривание йодотирозинов; 5) Захват и секрецию тироглобулина. 6) Синтез NIS посредством изменения транскрипции гена.
Действие ТТГ стимулирует: 1) Перенос йодида в полость фолликула; 2) Генерация Н 2 О 2; 3) Йодирование тироглобулина; 4) Спаривание йодотирозинов; 5) Захват и секрецию тироглобулина. 6) Синтез NIS посредством изменения транскрипции гена.
 Количество лактотрофов") Пролактин • Протеин 23 к. Да, секретируемый питуитарными лактотрофами (15 -20% популяции) Количество лактотрофов стремительно возрастает при высоком уровне эстрогенов, особенно во время беременности. • Одноцепочечный протеин из 199 АКостатков.
Пролактин • Протеин 23 к. Да, секретируемый питуитарными лактотрофами (15 -20% популяции) Количество лактотрофов стремительно возрастает при высоком уровне эстрогенов, особенно во время беременности. • Одноцепочечный протеин из 199 АКостатков.
 • Пролактин тонически ингибируется дофамином гипоталамуса, а также ГАМК, соматостатином, гонадолиберинассоциированным фактором, а стимулируется многими нейропептидами (серотонинергические и опиоидные пути), соматолиберином, стероидными гормонами и факторами роста.
• Пролактин тонически ингибируется дофамином гипоталамуса, а также ГАМК, соматостатином, гонадолиберинассоциированным фактором, а стимулируется многими нейропептидами (серотонинергические и опиоидные пути), соматолиберином, стероидными гормонами и факторами роста.
 → ↓ АЦ + активация") • Дофаминергическое ингибирование опосредуется Д 2 -рецепторами (Gi) → ↓ АЦ + активация К-тока → гиперполяризация мембраны → ↓ Сатока.
• Дофаминергическое ингибирование опосредуется Д 2 -рецепторами (Gi) → ↓ АЦ + активация К-тока → гиперполяризация мембраны → ↓ Сатока.
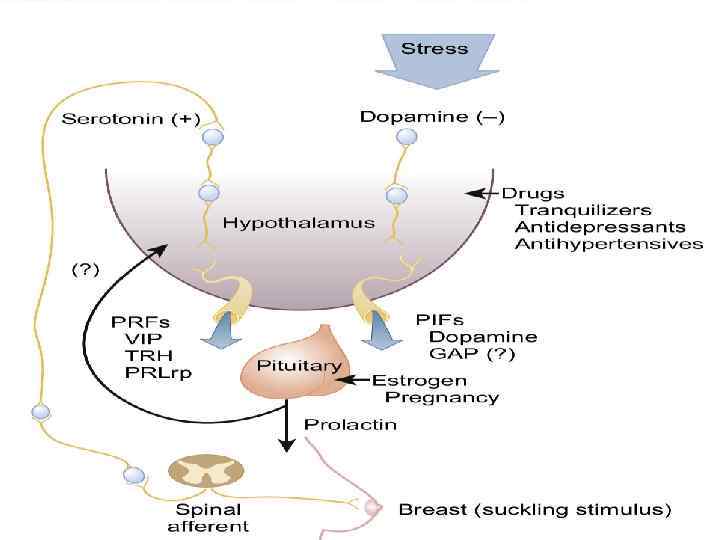
 Рефлекс стимул-секреция при раздражении рецепторов сосков; • 2) Повышение уровня") Стимулы выделения • 1) Рефлекс стимул-секреция при раздражении рецепторов сосков; • 2) Повышение уровня эстрогенов; • 3) Стресс. • Эстрогены стимулируют рост лактотрофов, а также экспрессию гена пролактина и его освобождение.
Стимулы выделения • 1) Рефлекс стимул-секреция при раздражении рецепторов сосков; • 2) Повышение уровня эстрогенов; • 3) Стресс. • Эстрогены стимулируют рост лактотрофов, а также экспрессию гена пролактина и его освобождение.
 ВИП; 2) Нейротензин; 3) Окситоцин; 4) Тиреолиберин; 5) Пролактин-релизинг-пептид;") Стимулы • • • 1) ВИП; 2) Нейротензин; 3) Окситоцин; 4) Тиреолиберин; 5) Пролактин-релизинг-пептид; Пролактин ингибирует сам себя, связываясь со своими рецепторами на дофаминергических нейронах, активируя их.
Стимулы • • • 1) ВИП; 2) Нейротензин; 3) Окситоцин; 4) Тиреолиберин; 5) Пролактин-релизинг-пептид; Пролактин ингибирует сам себя, связываясь со своими рецепторами на дофаминергических нейронах, активируя их.
 • Рецепторы пролактина относятся к группе тирозинкиназных рецепторов.
• Рецепторы пролактина относятся к группе тирозинкиназных рецепторов.
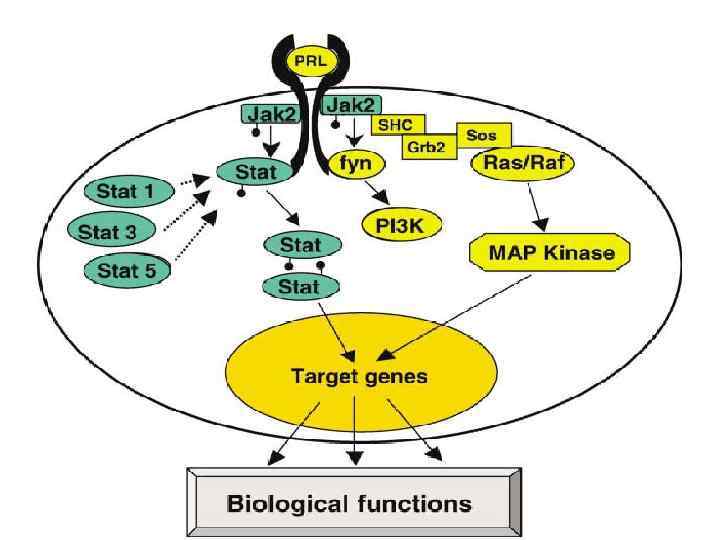
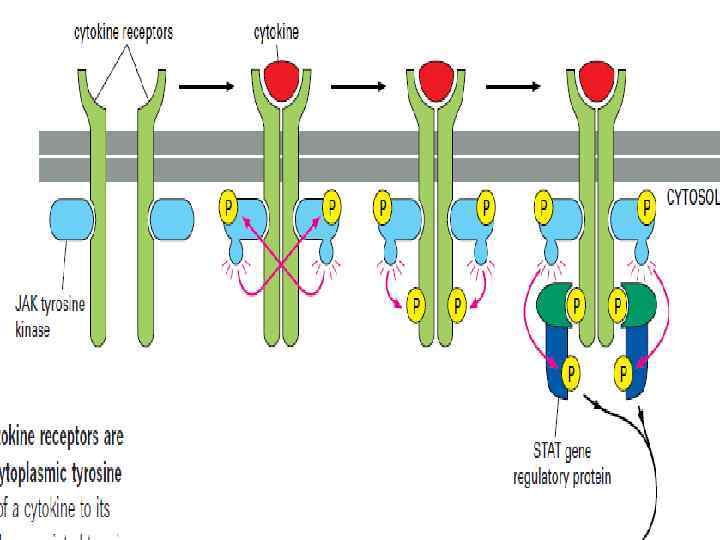
 Связывание вещества с рецептором приводит ассоциированные тирозинкиназы (JAK-киназы) к кроссфосфорилизации и") Jak/STAT • 1) Связывание вещества с рецептором приводит ассоциированные тирозинкиназы (JAK-киназы) к кроссфосфорилизации и активации друга. • 2) Активированные киназы затем фосфорилируют рецепторные протеины на тирозине.
Jak/STAT • 1) Связывание вещества с рецептором приводит ассоциированные тирозинкиназы (JAK-киназы) к кроссфосфорилизации и активации друга. • 2) Активированные киназы затем фосфорилируют рецепторные протеины на тирозине.
 Протеины, регулирующие генный аппарат, называемые STAT (передатчики сигнала и активаторы транскрипции),") • 3) Протеины, регулирующие генный аппарат, называемые STAT (передатчики сигнала и активаторы транскрипции), находятся в цитозоле, а затем прикрепляются к фосфорилированным тирозинам на рецепторах, и киназы активируют эти белки.
• 3) Протеины, регулирующие генный аппарат, называемые STAT (передатчики сигнала и активаторы транскрипции), находятся в цитозоле, а затем прикрепляются к фосфорилированным тирозинам на рецепторах, и киназы активируют эти белки.
 STAT затем отделяются от рецепторных белков, димеризуются и активируют транскрипцию определённых") • 4) STAT затем отделяются от рецепторных белков, димеризуются и активируют транскрипцию определённых генов-мишеней.
• 4) STAT затем отделяются от рецепторных белков, димеризуются и активируют транскрипцию определённых генов-мишеней.
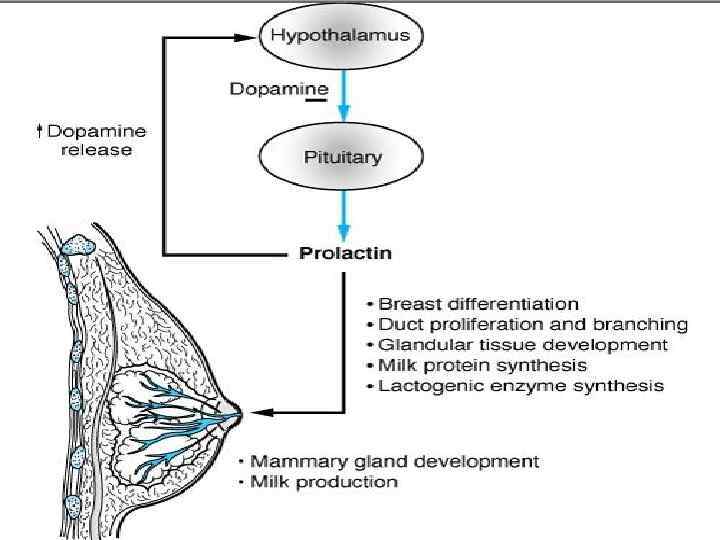
 Функции пролактина Около 100 функций. Репродукция, рост и развитие, осморегуляция, метаболизм, иммунорегуляция, когнитивные функции, поведение. Главная мишень пролактина – молочная железа. Пролактин стимулирует рост и дифференцировку эпителия молочных желёз, что является необходимым для начала и поддержания лактации.
Функции пролактина Около 100 функций. Репродукция, рост и развитие, осморегуляция, метаболизм, иммунорегуляция, когнитивные функции, поведение. Главная мишень пролактина – молочная железа. Пролактин стимулирует рост и дифференцировку эпителия молочных желёз, что является необходимым для начала и поддержания лактации.
 Стимуляция роста и развития молочных желез; • 2) Синтез молока; •") • 1) Стимуляция роста и развития молочных желез; • 2) Синтез молока; • 3) Поддержание секреции молока; • Пролактин стимулирует захват глюкозы и АК, синтез б-казеина и алактальбумина, лактозы и молочных жиров эпителиальными клетками железы.
• 1) Стимуляция роста и развития молочных желез; • 2) Синтез молока; • 3) Поддержание секреции молока; • Пролактин стимулирует захват глюкозы и АК, синтез б-казеина и алактальбумина, лактозы и молочных жиров эпителиальными клетками железы.
 • Во время беременности пролактин подготавливает грудь к лактации. • Пролактин модулирует репродуктивное и родительское поведение. • Пролактин – компонент иммунонейроэндокринной системы в воспалительном ответе. • Пролактин может действовать как цитокин, освобождаясь из клеток иммунной системы.
• Во время беременности пролактин подготавливает грудь к лактации. • Пролактин модулирует репродуктивное и родительское поведение. • Пролактин – компонент иммунонейроэндокринной системы в воспалительном ответе. • Пролактин может действовать как цитокин, освобождаясь из клеток иммунной системы.
 Спасибо за внимание!
Спасибо за внимание!