068d0a5e58b323fdcd28bd8e4d96a0f5.ppt
- Количество слайдов: 41
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА Репликация ДНК: процесс у прокариот Лекция 6
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА Репликация ДНК: процесс у прокариот Лекция 6
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Словарик
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Словарик
 Словарик Клеточный цикл – период жизни клетки от одного деления до другого или от деления до смерти Репликативная вилка – Y-образная структура, перемещающаяся вдоль родительской спирали ДНК и характеризующаяся местным расхождением двух её цепей, в пределах которой происходит активная репликация ДНК Ориджин (ori)– участок репликона, с которого начинается процесс репликации
Словарик Клеточный цикл – период жизни клетки от одного деления до другого или от деления до смерти Репликативная вилка – Y-образная структура, перемещающаяся вдоль родительской спирали ДНК и характеризующаяся местным расхождением двух её цепей, в пределах которой происходит активная репликация ДНК Ориджин (ori)– участок репликона, с которого начинается процесс репликации
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Общая характеристика процесса
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Общая характеристика процесса
 Репликация – полимеразная реакция, обеспечивающая процесс матричного синтеза молекулы ДНК и приводящая к ее самовоспроизведению (удвоению).
Репликация – полимеразная реакция, обеспечивающая процесс матричного синтеза молекулы ДНК и приводящая к ее самовоспроизведению (удвоению).
 Основные структурные элементы ЭЛЕМЕНТЫ РЕПЛИКОН ОРИДЖИН РЕПЛИКАТИВНАЯ ВИЛКА
Основные структурные элементы ЭЛЕМЕНТЫ РЕПЛИКОН ОРИДЖИН РЕПЛИКАТИВНАЯ ВИЛКА
 Репликонные системы
Репликонные системы
 Ориджины репликации Сборка белковых комплексов и формированние репликативной вилки начинается в определенной участке ДНК – Origin (Ori) Схема ori вируса SV 40 ORE (origin recognition element) – участок связывания белка Т-антигена (Т-аg) DUE (DNA unwinding element) – участок для связывания белка, расплетающего ДНК ТА – элемент, обогащенный АТ-нуклеотидами OBR (origin bidirectional replication) – начало двунаправленной репликации Аux-1 – вспомогательный элемент, связывает димеры Т-антигена Аuх-2 – связывает фактор транскрипции Sp 1
Ориджины репликации Сборка белковых комплексов и формированние репликативной вилки начинается в определенной участке ДНК – Origin (Ori) Схема ori вируса SV 40 ORE (origin recognition element) – участок связывания белка Т-антигена (Т-аg) DUE (DNA unwinding element) – участок для связывания белка, расплетающего ДНК ТА – элемент, обогащенный АТ-нуклеотидами OBR (origin bidirectional replication) – начало двунаправленной репликации Аux-1 – вспомогательный элемент, связывает димеры Т-антигена Аuх-2 – связывает фактор транскрипции Sp 1
 Формы движения репликативной вилки
Формы движения репликативной вилки
 Одноправленная репликативная вилка
Одноправленная репликативная вилка
 Двунаправленная репликативная вилка
Двунаправленная репликативная вилка
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Основные стадии репликации
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Основные стадии репликации
 Инициация • Образование пререпликативного комплекса • Формирование репликативной вилки Элонгация • Продолжение процесса репликации, рост дочерней цепи ДНК Терминация • Завершение процесса репликации
Инициация • Образование пререпликативного комплекса • Формирование репликативной вилки Элонгация • Продолжение процесса репликации, рост дочерней цепи ДНК Терминация • Завершение процесса репликации
 – один репликон") Хромосома E. coli (4, 2 млн. п. н. ) – один репликон
Хромосома E. coli (4, 2 млн. п. н. ) – один репликон
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Репликация ДНК у прокариот
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Репликация ДНК у прокариот
") ДНК-полимеразы прокариот (E. coli)
ДНК-полимеразы прокариот (E. coli)
 ДНК-полимеразы прокариот ДНК-полимераза III является репликазой, синтезирует обе цепи ДНК, обладает активностью 5’ → 3’ полимеразной и 3’ → 5’ экзонуклеазной ДНК-полимеразы I действует на запаздывающей цепи для удаления РНК-праймеров (5’ → 3’ экзонуклеазы) и дорепликации очищенных мест ДНК, выполняет репаративную функцию, обладает активностью 5’ → 3’ полимеразной и 3’ → 5’ экзонуклеазной ДНК-полимераза II имеет отношение лишь к репарации, обладает активностью 5’ → 3’ полимеразной и 3’ → 5’ экзонуклеазной
ДНК-полимеразы прокариот ДНК-полимераза III является репликазой, синтезирует обе цепи ДНК, обладает активностью 5’ → 3’ полимеразной и 3’ → 5’ экзонуклеазной ДНК-полимеразы I действует на запаздывающей цепи для удаления РНК-праймеров (5’ → 3’ экзонуклеазы) и дорепликации очищенных мест ДНК, выполняет репаративную функцию, обладает активностью 5’ → 3’ полимеразной и 3’ → 5’ экзонуклеазной ДНК-полимераза II имеет отношение лишь к репарации, обладает активностью 5’ → 3’ полимеразной и 3’ → 5’ экзонуклеазной
 Общий принцип строения ДНК-полимераз По форме ДНКполимеразы можно уподобить полураскрытой кисти правой руки, в которой ладонь, большой палец и остальные пальцы представляют три основных пространственных домена и формируют полость, удерживающую ДНКматрицу и затравку в ходе синтеза. « Пальцы» удерживают однонитевую матрицу, а « большой палец» прижимает праймер – матричный двунитевой участок.
Общий принцип строения ДНК-полимераз По форме ДНКполимеразы можно уподобить полураскрытой кисти правой руки, в которой ладонь, большой палец и остальные пальцы представляют три основных пространственных домена и формируют полость, удерживающую ДНКматрицу и затравку в ходе синтеза. « Пальцы» удерживают однонитевую матрицу, а « большой палец» прижимает праймер – матричный двунитевой участок.
 Скользящий зажим и погрузчик зажима ДНК-полимеразы работают совместно с различными белковыми комплексами, удерживающими их в вилке репликации. Их называют «зажим» и «загрузчик зажима» ( «sliding clamp» , «clamp loader» ).
Скользящий зажим и погрузчик зажима ДНК-полимеразы работают совместно с различными белковыми комплексами, удерживающими их в вилке репликации. Их называют «зажим» и «загрузчик зажима» ( «sliding clamp» , «clamp loader» ).
 Скользящий зажим и погрузчик зажима После объединения ДНК-полимеразы с зажимом, «загрузчик зажима» отходит от места реакции, но держится поближе к отстающей нити, чтобы провести загрузку на новом месте объединения праймерматрица, как только ДНК-полимераза диссоциирует при завершении синтеза предыдущего фрагмента Оказаки.
Скользящий зажим и погрузчик зажима После объединения ДНК-полимеразы с зажимом, «загрузчик зажима» отходит от места реакции, но держится поближе к отстающей нити, чтобы провести загрузку на новом месте объединения праймерматрица, как только ДНК-полимераза диссоциирует при завершении синтеза предыдущего фрагмента Оказаки.
 ДНК полимеразы прокариот: ДНК полимераза III Осуществляет синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе отстающей цепи. Holo-фермент состоит из 10 субъединиц, каждая из которых обладает своей функцией.
ДНК полимеразы прокариот: ДНК полимераза III Осуществляет синтез ведущей цепи ДНК и фрагментов Оказаки при синтезе отстающей цепи. Holo-фермент состоит из 10 субъединиц, каждая из которых обладает своей функцией.
 длиной 928 остатков, которую") ДНК полимераза I состоит из одной субъединицы (109 к. ДА) длиной 928 остатков, которую можно разделить на три домена : N-концевую треть молекулы занимает (5’→ 3’)-экзонуклеазный домен, в центре расположен самый короткий (3’→ 5’)-экзонуклеазный домен, а самый большой полимеразный домен находится на С-конце.
ДНК полимераза I состоит из одной субъединицы (109 к. ДА) длиной 928 остатков, которую можно разделить на три домена : N-концевую треть молекулы занимает (5’→ 3’)-экзонуклеазный домен, в центре расположен самый короткий (3’→ 5’)-экзонуклеазный домен, а самый большой полимеразный домен находится на С-конце.
 ДНК полимеразы прокариот: ДНК полимераза I
ДНК полимеразы прокариот: ДНК полимераза I
экзонуклеазного домена") ДНК полимеразы прокариот: ДНК полимераза II ДНК-полимераза II (состоит из N-концевого (3’→ 5’)экзонуклеазного домена и более длинного Сконцевого полимеразного домена, не обладает 5’экзонуклеазной активностью, лигируют повреждённую цепь ДНК
ДНК полимеразы прокариот: ДНК полимераза II ДНК-полимераза II (состоит из N-концевого (3’→ 5’)экзонуклеазного домена и более длинного Сконцевого полимеразного домена, не обладает 5’экзонуклеазной активностью, лигируют повреждённую цепь ДНК
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Стадия инициации у прокариот
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Стадия инициации у прокариот
, IHF (integration host factor), FIS") Общая схема репликация E. coli ГИСТОНОПОДОБНЫЕ БЕЛКИ: HU (…), IHF (integration host factor), FIS (factor for inversion stimulation) НЕ – HOLE-фермент
Общая схема репликация E. coli ГИСТОНОПОДОБНЫЕ БЕЛКИ: HU (…), IHF (integration host factor), FIS (factor for inversion stimulation) НЕ – HOLE-фермент
 Формирование пререпликативного комплекса и репликативной вилки Репликация в оri. С начинается с формирования комплекса, в состав которого входят шесть белков: Dna. А, Dna. В, Dna. С, НU, Girase и SSВ Структура (ori С) E. coli Особенности структуры ДНК Ori-участков: 1. негативная суперспирализация ДНК 2. присутствием «легкоплавких» А-Т участков ДНК 3. Места для связывания со вспомогательными белками
Формирование пререпликативного комплекса и репликативной вилки Репликация в оri. С начинается с формирования комплекса, в состав которого входят шесть белков: Dna. А, Dna. В, Dna. С, НU, Girase и SSВ Структура (ori С) E. coli Особенности структуры ДНК Ori-участков: 1. негативная суперспирализация ДНК 2. присутствием «легкоплавких» А-Т участков ДНК 3. Места для связывания со вспомогательными белками
 узнает область начала репликации ori. C, последовательно связываясь с") Белок Dna. A Функции: 1) узнает область начала репликации ori. C, последовательно связываясь с нонамерными повторами в ДНК – блоками Dna. A 2) способствует расплетанию легкоплавких АТ-богатых участков ДНК ori. C 3) вербует на расплетенные участки ori. C Dna. B (хеликазу) Структура белка Dna. A E. coli. I-IV - области гомологии бактериальных белков Dna. A 1 - домен олигомеризации 2 - области связывания с хеликазой Dna. В, 3 - сайт связывания нуклеотидов 4 - домен АТФазной активности 5 - участок взаимодействия с мембранными фосфолипидами 6 - домен связывания с ДНК
Белок Dna. A Функции: 1) узнает область начала репликации ori. C, последовательно связываясь с нонамерными повторами в ДНК – блоками Dna. A 2) способствует расплетанию легкоплавких АТ-богатых участков ДНК ori. C 3) вербует на расплетенные участки ori. C Dna. B (хеликазу) Структура белка Dna. A E. coli. I-IV - области гомологии бактериальных белков Dna. A 1 - домен олигомеризации 2 - области связывания с хеликазой Dna. В, 3 - сайт связывания нуклеотидов 4 - домен АТФазной активности 5 - участок взаимодействия с мембранными фосфолипидами 6 - домен связывания с ДНК
 Связывание инициаторного белка Dna A c ДНК Сначала с 9 pb последовательностью связывается мономер Dna. А, затем 20– 40 мономеров этого белка формируют большой агрегат. ДНК ориджина опоясывает его, и цепи ДНК разъединяются в области трех 13 pb последовательностей. На следующем этапе димер Dna. В/Dпа. С присоединяется к комплексу ori. С/Dna. А – формируется репликативная вилка.
Связывание инициаторного белка Dna A c ДНК Сначала с 9 pb последовательностью связывается мономер Dna. А, затем 20– 40 мономеров этого белка формируют большой агрегат. ДНК ориджина опоясывает его, и цепи ДНК разъединяются в области трех 13 pb последовательностей. На следующем этапе димер Dna. В/Dпа. С присоединяется к комплексу ori. С/Dna. А – формируется репликативная вилка.
 присоединение Dna. А (инициирующий белок) b) присоединение Dna. B (хеликазы) c) Присоединение и") а) присоединение Dna. А (инициирующий белок) b) присоединение Dna. B (хеликазы) c) Присоединение и Dna. G (праймазы) – образование праймосомы , комплекса, содержащего хеликазу и праймазу
а) присоединение Dna. А (инициирующий белок) b) присоединение Dna. B (хеликазы) c) Присоединение и Dna. G (праймазы) – образование праймосомы , комплекса, содержащего хеликазу и праймазу
 Dna. В расплетает двухцепочечную ДНК при использовании энергии АТФ. Скорость расплетения составляет около 6000 нукл. в сек.
Dna. В расплетает двухцепочечную ДНК при использовании энергии АТФ. Скорость расплетения составляет около 6000 нукл. в сек.
 SSB-белки удерживают одноцепочечные ДНК от ассоциации. По мере продвижения репликационной вилки SSB диссоциируют с одного места и присоединяются на другом. Этот процесс не требует затрат энергии АТФ.
SSB-белки удерживают одноцепочечные ДНК от ассоциации. По мере продвижения репликационной вилки SSB диссоциируют с одного места и присоединяются на другом. Этот процесс не требует затрат энергии АТФ.
 Формируется праймосома. Фермент праймаза синтезирует праймеры. ДНК-полимераза III присоединяет к праймеру первый нуклеотид.
Формируется праймосома. Фермент праймаза синтезирует праймеры. ДНК-полимераза III присоединяет к праймеру первый нуклеотид.
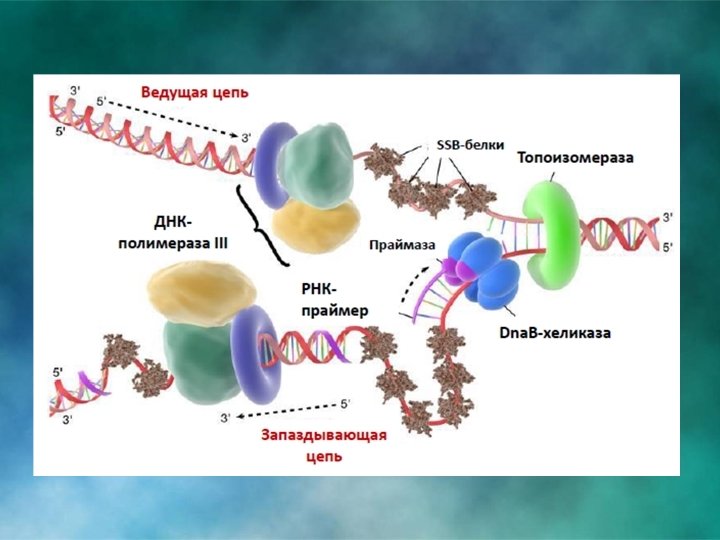
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Стадия элонгации у прокариот
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Стадия элонгации у прокариот
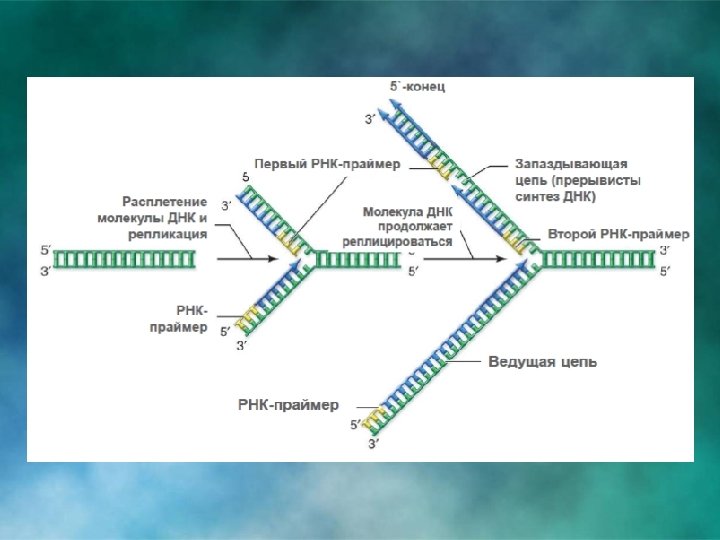
 ДНК-полимераза III присоединяет к праймеру следующий нуклеотид. Идет синтез ведущей и отстающей цепи. Обе цепи реплицируются синхронно, что достигается взаимодействием хеликазы, праймазы и ДНК-полимеразы и торможением синтеза лидирующей цепи во время синтеза праймеров.
ДНК-полимераза III присоединяет к праймеру следующий нуклеотид. Идет синтез ведущей и отстающей цепи. Обе цепи реплицируются синхронно, что достигается взаимодействием хеликазы, праймазы и ДНК-полимеразы и торможением синтеза лидирующей цепи во время синтеза праймеров.
 Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента ДНК-полимераза I удаляет праймер, имеющейся у полимераз 5‘ → 3' экзонуклеазной и 5’ → 3’ полимеразной активностью (исключение праймера). После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи ДНК-лигазы.
Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента ДНК-полимераза I удаляет праймер, имеющейся у полимераз 5‘ → 3' экзонуклеазной и 5’ → 3’ полимеразной активностью (исключение праймера). После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи ДНК-лигазы.
 МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Стадия терминации у прокариот
МОЛЕКУЛЯРНАЯ ГЕНЕТИКА. РЕПЛИКАЦИЯ Стадия терминации у прокариот
 36 к. Да Терминация репликации у прокариот") Организация репликона E. coli aattagtatgttgtaactaaagt tus (tbp) 36 к. Да Терминация репликации у прокариот происходит, когда «репликационная вилка» достигает участка ДНК с особыми сайтами ter и, если с ДНК этих сайтов соединится продукт гена tus.
Организация репликона E. coli aattagtatgttgtaactaaagt tus (tbp) 36 к. Да Терминация репликации у прокариот происходит, когда «репликационная вилка» достигает участка ДНК с особыми сайтами ter и, если с ДНК этих сайтов соединится продукт гена tus.
 Общая схема репликации у прокариот Последовательность событий 1. Dna. A узнает Dna. A Box (регуляторный элемент около Ori), что приводит к к расплетанию DNA в Ori 2. Хеликазы Dna. B и Dna. C связывают репликативную вилку с противоположных сторон 3. Dna. G (праймаза) присоединяется к комплексу, Dna. C высвобождается, остается комплекс xеликазы (Dna. B) и праймазы (Dna. G) 4. Хеликаза (Dna. B) расплетает DNA, присоединяются белки SSB 5. Dna. G (праймаза) синтезирует праймер 6. ДНК-полимераза III синтезирует лидирующую цепь и отстающую цепи 7. ДНК-полимераза I заменяет праймер, ДНК-лигаза сшивает фрагменты 8. Tus белки связываются с ter-последовательностями и останавливают репликацию 9. Топоизомераза IV разъединяет ДНК
Общая схема репликации у прокариот Последовательность событий 1. Dna. A узнает Dna. A Box (регуляторный элемент около Ori), что приводит к к расплетанию DNA в Ori 2. Хеликазы Dna. B и Dna. C связывают репликативную вилку с противоположных сторон 3. Dna. G (праймаза) присоединяется к комплексу, Dna. C высвобождается, остается комплекс xеликазы (Dna. B) и праймазы (Dna. G) 4. Хеликаза (Dna. B) расплетает DNA, присоединяются белки SSB 5. Dna. G (праймаза) синтезирует праймер 6. ДНК-полимераза III синтезирует лидирующую цепь и отстающую цепи 7. ДНК-полимераза I заменяет праймер, ДНК-лигаза сшивает фрагменты 8. Tus белки связываются с ter-последовательностями и останавливают репликацию 9. Топоизомераза IV разъединяет ДНК