

ДНК цитогенетика репликация 16.09.16.ppt
- Количество слайдов: 97
 Молекулярная генетика Горожанина Е. С.
Молекулярная генетика Горожанина Е. С.
 www. mma. ru Образование Лечебный факультет Кафедры Кафедра биологии и общей генетики Учебные материалы Лекции и модули
www. mma. ru Образование Лечебный факультет Кафедры Кафедра биологии и общей генетики Учебные материалы Лекции и модули
 • элементарная единица наследственности, содержащая информацию, равномерно") Понятие «ген» (термин предложен В. Йогансеном, 1909) • элементарная единица наследственности, содержащая информацию, равномерно распределяемую между дочерними клетками при делении; • фрагмент молекулы ДНК (ДНК – носитель генетической информации); • функционально активная единица молекулы ДНК, предназначенная для регулируемого синтеза молекулы РНК
Понятие «ген» (термин предложен В. Йогансеном, 1909) • элементарная единица наследственности, содержащая информацию, равномерно распределяемую между дочерними клетками при делении; • фрагмент молекулы ДНК (ДНК – носитель генетической информации); • функционально активная единица молекулы ДНК, предназначенная для регулируемого синтеза молекулы РНК
 Нуклеиновые кислоты открыты Ф. Мишером в 1868 г.
Нуклеиновые кислоты открыты Ф. Мишером в 1868 г.
 1928 г. опыты Ф. Гриффита
1928 г. опыты Ф. Гриффита
 Явление переноса генетической информации от одного штамма в другой, Гриффит назвал трансформацией. Механизм трансформации и природа трансформирующего фактора была неизвестна до 1944 г.
Явление переноса генетической информации от одного штамма в другой, Гриффит назвал трансформацией. Механизм трансформации и природа трансформирующего фактора была неизвестна до 1944 г.
 В 1929 году Н. К. Кольцов вывел теорию, что ген – это белок, и гены могут самокопироваться. В 40 -х годах стали накапливаться косвенные факты, что ген – это не белок, а ДНК
В 1929 году Н. К. Кольцов вывел теорию, что ген – это белок, и гены могут самокопироваться. В 40 -х годах стали накапливаться косвенные факты, что ген – это не белок, а ДНК
 О. Эвери разделил штаммы") 1944 г. опыты О. Эвери (совместно с Мак-Карти и Мак-Леодом) О. Эвери разделил штаммы пневмококков на отдельные компоненты (углеводы, липиды, белки и ДНК) и добавлял к штаммам гладких вирулентных. Добавление фермента, разрушающего ДНК, препятствовало трансформации. Показано, что трансформирующим агентом является ДНК, т. е. ДНК - носитель генетической информации После этого начинается эра изучения ДНК
1944 г. опыты О. Эвери (совместно с Мак-Карти и Мак-Леодом) О. Эвери разделил штаммы пневмококков на отдельные компоненты (углеводы, липиды, белки и ДНК) и добавлял к штаммам гладких вирулентных. Добавление фермента, разрушающего ДНК, препятствовало трансформации. Показано, что трансформирующим агентом является ДНК, т. е. ДНК - носитель генетической информации После этого начинается эра изучения ДНК
 Метили белки и ДНК бактериофагов. В") А. Херши и М. Чейз (1952 г. ) Метили белки и ДНК бактериофагов. В заражённую клетку проникает только НК фага, а новые фаги содержат такие же белки и НК, как исходный фаг ДНК - носитель генетической информации
А. Херши и М. Чейз (1952 г. ) Метили белки и ДНК бактериофагов. В заражённую клетку проникает только НК фага, а новые фаги содержат такие же белки и НК, как исходный фаг ДНК - носитель генетической информации
 Нуклеотид — структурная единица НК 1. Сахар – пентоза 2. Остаток фосфорной кислоты 3. Азотистое основание: А (аденин) Г (гуанин) Т (тимин) Ц(цитозин) У (урацил)
Нуклеотид — структурная единица НК 1. Сахар – пентоза 2. Остаток фосфорной кислоты 3. Азотистое основание: А (аденин) Г (гуанин) Т (тимин) Ц(цитозин) У (урацил)


 1950 г. правило Чаргаффа А +Г=Т+Ц А=Т Г Ц") Эрвин Чаргафф (1905 – 2002) 1950 г. правило Чаргаффа А +Г=Т+Ц А=Т Г Ц
Эрвин Чаргафф (1905 – 2002) 1950 г. правило Чаргаффа А +Г=Т+Ц А=Т Г Ц
 1953 г. Модель ДНК Дж. Уотсон и Ф. Крик
1953 г. Модель ДНК Дж. Уотсон и Ф. Крик
 и Морис Уилкинс Рентгеноструктурный анализ Спиральность Двуцепочечность") Розалинда Франклин (1920 — 1958) и Морис Уилкинс Рентгеноструктурный анализ Спиральность Двуцепочечность
Розалинда Франклин (1920 — 1958) и Морис Уилкинс Рентгеноструктурный анализ Спиральность Двуцепочечность
 1962 г. Нобелевская премия по физиологии и медицине Дж. Уотсон и Фр. Крик
1962 г. Нобелевская премия по физиологии и медицине Дж. Уотсон и Фр. Крик
 Образование фосфодиэфирной связи между нуклеотидами
Образование фосфодиэфирной связи между нуклеотидами
 Типы связей в молекуле ДНК
Типы связей в молекуле ДНК
 Антипараллельность 3` - конец одной расположен") Структура ДНК Двуцепочечность (струкурная основа передачи генетической информации) Антипараллельность 3` - конец одной расположен напротив 5` конца другой. Комплементарность А= Т Ц=Г
Структура ДНК Двуцепочечность (струкурная основа передачи генетической информации) Антипараллельность 3` - конец одной расположен напротив 5` конца другой. Комплементарность А= Т Ц=Г
 Между нуклеотидами 0, 34 нм Один виток — 3, 4 нм (в") Правозакрученность (β-форма) Между нуклеотидами 0, 34 нм Один виток — 3, 4 нм (в одном витке 10 нуклеотидов) Плоскости азотистых оснований перпендикулярны оси спирали Диаметр молекулы — 2 нм (пурин 1, 2 нм; пиримидин 0, 8 нм)
Правозакрученность (β-форма) Между нуклеотидами 0, 34 нм Один виток — 3, 4 нм (в одном витке 10 нуклеотидов) Плоскости азотистых оснований перпендикулярны оси спирали Диаметр молекулы — 2 нм (пурин 1, 2 нм; пиримидин 0, 8 нм)
 Основные формы ДНК
Основные формы ДНК
 Альтернативные формы ДНК β - форма В водных растворах низкой ионной силы, основная конформация α - форма В растворах с высокой ионной силой или в сухой ДНК, есть внутренняя пустота, диаметром 0, 5 нм z - форма Левозакрученная, содержит только Ц и Г пары
Альтернативные формы ДНК β - форма В водных растворах низкой ионной силы, основная конформация α - форма В растворах с высокой ионной силой или в сухой ДНК, есть внутренняя пустота, диаметром 0, 5 нм z - форма Левозакрученная, содержит только Ц и Г пары
 Функции ДНК : Носитель генетической информации. Воспроизведение генетической информации - репликация. Реализация генетической информации транскрипция и трансляция Вариабельность генетической информации
Функции ДНК : Носитель генетической информации. Воспроизведение генетической информации - репликация. Реализация генетической информации транскрипция и трансляция Вариабельность генетической информации
 Цитогенетика
Цитогенетика
 числом хромосом 2) размером 3)") Кариотип - набор хромосом ядра соматической клетки, характеризуется: 1) числом хромосом 2) размером 3) парностью 4) набором генов в хромосоме
Кариотип - набор хромосом ядра соматической клетки, характеризуется: 1) числом хромосом 2) размером 3) парностью 4) набором генов в хромосоме
 Кариотип человека
Кариотип человека
 Кариотип-биологическая константа
Кариотип-биологическая константа
") Структуру хромосом описал Сергей Гаврилович Навашин (1904 г. )
Структуру хромосом описал Сергей Гаврилович Навашин (1904 г. )
 Типы хромосом эукариот в зависимости от положения центромеры
Типы хромосом эукариот в зависимости от положения центромеры
 Эухроматин и гетерохроматин Эухроматиновые участки хромосом менее плотные и потенциально транскрибируемые. Гетерохроматиновые участки хромосом компактные и генетически инертные.
Эухроматин и гетерохроматин Эухроматиновые участки хромосом менее плотные и потенциально транскрибируемые. Гетерохроматиновые участки хромосом компактные и генетически инертные.

") Гетерохроматин Факультативный Конститутивный - в области центромеры - в теломерных участках хромосом (нетранскрибируемая ДНК) - половой хроматин одна из двух Х хромосом у человека (тельце Барра)
Гетерохроматин Факультативный Конститутивный - в области центромеры - в теломерных участках хромосом (нетранскрибируемая ДНК) - половой хроматин одна из двух Х хромосом у человека (тельце Барра)
.") Факультативный гетерохроматин Половой хроматин - одна из двух Х хромосом у женщины (тельце Барра). Гены этой хромосомы не транскрибируются. Количество телец Барра на 1 меньше числа Х-хромосом: 46 ХХ - 1 46 XY - 0 47 XXY - 1 47 XXX - 2
Факультативный гетерохроматин Половой хроматин - одна из двух Х хромосом у женщины (тельце Барра). Гены этой хромосомы не транскрибируются. Количество телец Барра на 1 меньше числа Х-хромосом: 46 ХХ - 1 46 XY - 0 47 XXY - 1 47 XXX - 2
 ХAХа Х A Х а Ха Y ХAY
ХAХа Х A Х а Ха Y ХAY

") Гетерохроматин Факультативный Конститутивный - в области центромеры - в теломерных участках хромосом (нетранскрибируемая ДНК) - половой хроматин одна из двух Х хромосом у человека (тельце Барра)
Гетерохроматин Факультативный Конститутивный - в области центромеры - в теломерных участках хромосом (нетранскрибируемая ДНК) - половой хроматин одна из двух Х хромосом у человека (тельце Барра)
 -") Конститутивный гетерохроматин: теломеры – - «запечатывают» концы хромосом - многочисленные нуклеотидные повторы (ТТАГГГ) - длина 2 -20 тыс. пар нуклеотидов n(TTAGGG)
Конститутивный гетерохроматин: теломеры – - «запечатывают» концы хромосом - многочисленные нуклеотидные повторы (ТТАГГГ) - длина 2 -20 тыс. пар нуклеотидов n(TTAGGG)
 Значение теломер - прикрепляют хромосому к внутренней ядерной мембране в строго - п определённом месте определённом - предотвращают слипание хромосом своими концами - определяют количество митотических циклов клетки, длина теломер – счетчик времени, определяющий старение
Значение теломер - прикрепляют хромосому к внутренней ядерной мембране в строго - п определённом месте определённом - предотвращают слипание хромосом своими концами - определяют количество митотических циклов клетки, длина теломер – счетчик времени, определяющий старение

 - «лимит") Теломеры укорачиваются при каждом делении (количество делений клеток человека – 20 -70) - «лимит Хайфлика» Теломераза не даёт укорачиваться хромосомам
Теломеры укорачиваются при каждом делении (количество делений клеток человека – 20 -70) - «лимит Хайфлика» Теломераза не даёт укорачиваться хромосомам
") Работа теломеразы (2009 г. Нобелевская премия)
Работа теломеразы (2009 г. Нобелевская премия)
 Теломеры укорачиваются с возрастом и влияют на продолжительность жизни особи Прогерия — раннее старение организма в результате уменьшения длины теломер в стволовых клетках
Теломеры укорачиваются с возрастом и влияют на продолжительность жизни особи Прогерия — раннее старение организма в результате уменьшения длины теломер в стволовых клетках
, приводящая к") Детская прогерия синдром Хатчинсона-Джилфорда Мутация гена, кодирующего ламин (белок оболочки клеточного ядра), приводящая к нарушению репарации ДНК
Детская прогерия синдром Хатчинсона-Джилфорда Мутация гена, кодирующего ламин (белок оболочки клеточного ядра), приводящая к нарушению репарации ДНК
 Прогерия взрослых синдром Вернера Дефект гена АТФ-зависимой хеликазы, ответственной за репарацию ДНК
Прогерия взрослых синдром Вернера Дефект гена АТФ-зависимой хеликазы, ответственной за репарацию ДНК
 3. Петельный (хромомерный) 4 -5.") 1. Уровни компактизации хромосом 1. Нуклеосомный 2. Соленоидный (нуклеомерный) 3. Петельный (хромомерный) 4 -5. Хромонемный, хромосомный
1. Уровни компактизации хромосом 1. Нуклеосомный 2. Соленоидный (нуклеомерный) 3. Петельный (хромомерный) 4 -5. Хромонемный, хромосомный
 1. Нуклеосомный уровень нуклеосома – структурная единица хромосомы Белки гистоны: H 2 A, H 2 B, H 3, H 4 белковый кор ДНК делает 1, 75 оборота (~146 п. н. ) Участок ДНК, не связанный с белковым кором из 50 -60 п. н. - линкер или линкерная ДНК
1. Нуклеосомный уровень нуклеосома – структурная единица хромосомы Белки гистоны: H 2 A, H 2 B, H 3, H 4 белковый кор ДНК делает 1, 75 оборота (~146 п. н. ) Участок ДНК, не связанный с белковым кором из 50 -60 п. н. - линкер или линкерная ДНК
 Нуклеосомная нить это связанные между собой нуклеосомы - диаметр нуклеосомной нити – 10 -11 нм - компактизация (укорочение) в 6 -7 раз
Нуклеосомная нить это связанные между собой нуклеосомы - диаметр нуклеосомной нити – 10 -11 нм - компактизация (укорочение) в 6 -7 раз

 2. Нуклеомерный или соленоидный уровень - идёт путем объединения 6 -7 нуклеосом в глобулу. Гистон Н 1 сближает соседние нуклеосомы, «сшивая и укорачивая» линкерную ДНК
2. Нуклеомерный или соленоидный уровень - идёт путем объединения 6 -7 нуклеосом в глобулу. Гистон Н 1 сближает соседние нуклеосомы, «сшивая и укорачивая» линкерную ДНК
 Диаметр соленоида 30 нм Компактизация в 40 -50 раз
Диаметр соленоида 30 нм Компактизация в 40 -50 раз

 связаны с негистоновыми белками (конденсин), образующими остов") 3. Хромомерный или петельный уровень Фибриллы (ДНП) связаны с негистоновыми белками (конденсин), образующими остов (каркас). Одна петля может соответствовать одному или нескольким генам В петле 50– 200 тыс. п. н.
3. Хромомерный или петельный уровень Фибриллы (ДНП) связаны с негистоновыми белками (конденсин), образующими остов (каркас). Одна петля может соответствовать одному или нескольким генам В петле 50– 200 тыс. п. н.
 уровень Диаметр 300 нм Компактизация в 700 раз") Хромомерный (петельный) уровень Диаметр 300 нм Компактизация в 700 раз
Хромомерный (петельный) уровень Диаметр 300 нм Компактизация в 700 раз

 уровень У человека во всех хромосомах 2600 петель Вернуться") Хромомерный (петельный) уровень У человека во всех хромосомах 2600 петель Вернуться
Хромомерный (петельный) уровень У человека во всех хромосомах 2600 петель Вернуться
 Петля обеспечивает экспрессию гена, т. е петля структурное и функциональное образование
Петля обеспечивает экспрессию гена, т. е петля структурное и функциональное образование
 4. Хромонемный уровень Конденсированные участки одной из метафазных хроматид Диаметр 700 нм Компактизация в 1500 раз
4. Хромонемный уровень Конденсированные участки одной из метафазных хроматид Диаметр 700 нм Компактизация в 1500 раз
 5. Хромосомный уровень Метафазная хромосома уже удвоена. Она состоит из двух хроматид. Каждая из них содержит молекулу ДНК Размер 1400 х 2000 нм (1, 4 х 2, 0 мкм) Укорочение ДНК (итоговая компактизация) в 10 000 раз
5. Хромосомный уровень Метафазная хромосома уже удвоена. Она состоит из двух хроматид. Каждая из них содержит молекулу ДНК Размер 1400 х 2000 нм (1, 4 х 2, 0 мкм) Укорочение ДНК (итоговая компактизация) в 10 000 раз


 Татсуйя Хирано 1997 г. открытие белкового комплекса конденсина Когезин формирует мост между хроматидами Конденсин конденсирует сестринские хроматиды
Татсуйя Хирано 1997 г. открытие белкового комплекса конденсина Когезин формирует мост между хроматидами Конденсин конденсирует сестринские хроматиды
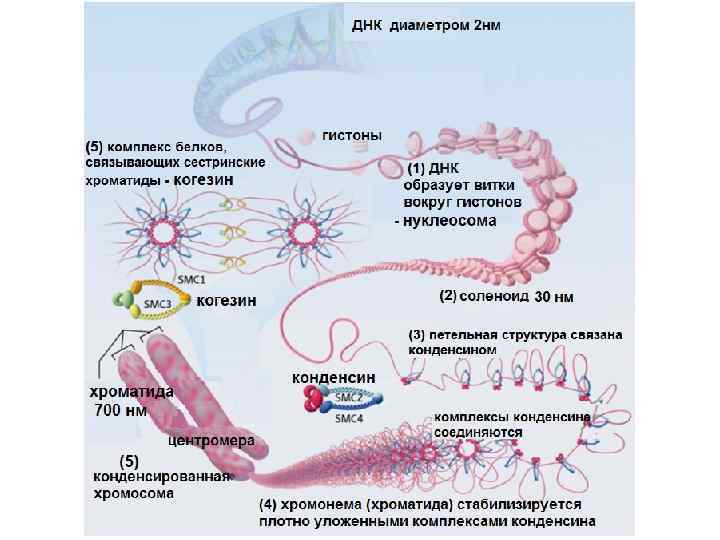
. Когда клетка готовится к") После репликации ДНК когезин связывает реплицированные пары ДНК (сестринские хроматиды). Когда клетка готовится к делению, почти весь когезин отделяется, за исключением небольшого количества вблизи центромеры. В анафазе отделяется остаточный пул когезина, и сестринские хроматиды разделяются и расходятся по двум дочерним клеткам.
После репликации ДНК когезин связывает реплицированные пары ДНК (сестринские хроматиды). Когда клетка готовится к делению, почти весь когезин отделяется, за исключением небольшого количества вблизи центромеры. В анафазе отделяется остаточный пул когезина, и сестринские хроматиды разделяются и расходятся по двум дочерним клеткам.
 Политенные хромосомы содержат многие сотни копий ДНК образуются в результате многократной репликации ДНК без последующего расхождения хроматид видны в интерфазе
Политенные хромосомы содержат многие сотни копий ДНК образуются в результате многократной репликации ДНК без последующего расхождения хроматид видны в интерфазе
 Пуф — вздутие в результате декомпатизации ДНК Кольца Бальбиани — крупный пуф
Пуф — вздутие в результате декомпатизации ДНК Кольца Бальбиани — крупный пуф
 Биологическое значение хромосомного уровня организации наследственного материала - даёт упорядоченность пространственной организации генетического материала по группам сцепления - сохраняет постоянство генетического материала при репликации и делении - в группах сцепления происходит рекомбинация генетического материала при мейозе и оплодотворении
Биологическое значение хромосомного уровня организации наследственного материала - даёт упорядоченность пространственной организации генетического материала по группам сцепления - сохраняет постоянство генетического материала при репликации и делении - в группах сцепления происходит рекомбинация генетического материала при мейозе и оплодотворении

 Нуклеоид прокариот
Нуклеоид прокариот
 Начало репликации – точка origin (ori)") Репликация У прокариот монорепликонный механизм (1 репликон) Начало репликации – точка origin (ori)
Репликация У прокариот монорепликонный механизм (1 репликон) Начало репликации – точка origin (ori)
 Полирепликонный механизм у эукариот (до 1000 репликонов в 1 молекуле ДНК, 10 000 ориджинов у человека)
Полирепликонный механизм у эукариот (до 1000 репликонов в 1 молекуле ДНК, 10 000 ориджинов у человека)
 Репликация ДНК
Репликация ДНК
 ДНК-хеликаза Раскручивает двойную спираль, разрывая водородные связи, используя энергию АТФ
ДНК-хеликаза Раскручивает двойную спираль, разрывая водородные связи, используя энергию АТФ
 ДНК-топоизомераза Разрывает одну из нитей ДНК, давая ей возможность вращаться вокруг второй цепи для снятия напряжения на сверхзакрученных участках ДНК. Она же восстанавливает целостность нити
ДНК-топоизомераза Разрывает одну из нитей ДНК, давая ей возможность вращаться вокруг второй цепи для снятия напряжения на сверхзакрученных участках ДНК. Она же восстанавливает целостность нити
 SSb-белки Связываются с одноцепочечной ДНК, взаимодействуя с сахарнофосфатным остовом. Не позволяют вновь соединиться двум нитям и не закрывают азотистые основания
SSb-белки Связываются с одноцепочечной ДНК, взаимодействуя с сахарнофосфатным остовом. Не позволяют вновь соединиться двум нитям и не закрывают азотистые основания
 для предоставления свободного 3′-ОН конца ДНК-полимеразе") РНК-праймаза Синтезирует короткие РНК-праймеры (РНК-затравки) для предоставления свободного 3′-ОН конца ДНК-полимеразе
РНК-праймаза Синтезирует короткие РНК-праймеры (РНК-затравки) для предоставления свободного 3′-ОН конца ДНК-полимеразе
 ДНК-полимераза Наращивает полинуклеотидные нити, присоединяя нуклеотиды друг к другу, на ведущей цепи непрерывно, на отстающей — фрагментами Оказаки
ДНК-полимераза Наращивает полинуклеотидные нити, присоединяя нуклеотиды друг к другу, на ведущей цепи непрерывно, на отстающей — фрагментами Оказаки
 Лидирующая цепь: матричная 3`-5`, на ней дочерняя 5`- 3` Отстающая цепь фрагментами Оказаки: матричная 5`- 3`, дочерняя 3`-5`
Лидирующая цепь: матричная 3`-5`, на ней дочерняя 5`- 3` Отстающая цепь фрагментами Оказаки: матричная 5`- 3`, дочерняя 3`-5`
 Эндонуклеаза Удаляет РНК-затравки, участвует в репарации
Эндонуклеаза Удаляет РНК-затравки, участвует в репарации
 Лигаза Сшивает фрагменты Оказаки
Лигаза Сшивает фрагменты Оказаки
 Механизм репликации ДНК
Механизм репликации ДНК
 ДНК – полимеразы у прокариот У E. coli – 3 типа ДНК – полимеразы у эукариот – 7 типов отвечают за репликацию и репарацию
ДНК – полимеразы у прокариот У E. coli – 3 типа ДНК – полимеразы у эукариот – 7 типов отвечают за репликацию и репарацию
 Репликация ДНК Полуконсервативная репликация
Репликация ДНК Полуконсервативная репликация

 Репарация - восстановление повреждений, возникающих в ДНК Репликация ДНК без репарации 1 ошибка на 107 скопированных нуклеотидов Репликация ДНК с участием репарации 1 ошибка на 109 нуклеотидов (99% исправлено)
Репарация - восстановление повреждений, возникающих в ДНК Репликация ДНК без репарации 1 ошибка на 107 скопированных нуклеотидов Репликация ДНК с участием репарации 1 ошибка на 109 нуклеотидов (99% исправлено)
 - эксцизионная (дорепликативная") Репарация восстановление повреждений, возникающих в ДНК Виды репарации: - световая (фотореактивация) - эксцизионная (дорепликативная и репликативная) - пострепликативная - SOS репарация
Репарация восстановление повреждений, возникающих в ДНК Виды репарации: - световая (фотореактивация) - эксцизионная (дорепликативная и репликативная) - пострепликативная - SOS репарация
 Световая репарация Под влиянием УФизлучения образуются димеры пиримидиновых оснований. Фермент фотолиаза восстанавливает структуру ДНК
Световая репарация Под влиянием УФизлучения образуются димеры пиримидиновых оснований. Фермент фотолиаза восстанавливает структуру ДНК
 G") Эксцизионная репарация это восстановления повреждённого участка ДНК, путем его вырезания, происходит в 1) G 1 -периоде (дорепликативная) 2) S-периоде (репликативная) Essential Cell Biology (© Garland Science 2010)
Эксцизионная репарация это восстановления повреждённого участка ДНК, путем его вырезания, происходит в 1) G 1 -периоде (дорепликативная) 2) S-периоде (репликативная) Essential Cell Biology (© Garland Science 2010)
 между двумя вновь образованными спиралями ДНК. Происходит в") Пострепликативная репарация - обмен фрагментами (рекомбинация) между двумя вновь образованными спиралями ДНК. Происходит в G 2 — периоде клеточного цикла
Пострепликативная репарация - обмен фрагментами (рекомбинация) между двумя вновь образованными спиралями ДНК. Происходит в G 2 — периоде клеточного цикла
") Гомологичная рекомбинация репарирует двуцепочечные разрывы Essential Cell Biology (© Garland Science 2010)
Гомологичная рекомбинация репарирует двуцепочечные разрывы Essential Cell Biology (© Garland Science 2010)
 SOS-репарация достраивание ДНК напротив дефектных звеньев матричной цепи. В результате ДНК оказывается удвоенной, хотя и с большим количеством ошибок, но это даёт возможность провести клеточное деление.
SOS-репарация достраивание ДНК напротив дефектных звеньев матричной цепи. В результате ДНК оказывается удвоенной, хотя и с большим количеством ошибок, но это даёт возможность провести клеточное деление.
 – снижение") Заболевания, обусловленные дефектами системы репарации: • Пигментная ксеродерма (пятна, короста, рак кожи) – снижение активности УФ-эндонуклеазы; • Синдром Блума (глубокие поражения капилляров на лице) – мутация ДНК-лигазы; • Злокачественные перерождения – нарушение репарации неспаренных нуклеотидов. Пигментная ксеродерма Синдром Блума
Заболевания, обусловленные дефектами системы репарации: • Пигментная ксеродерма (пятна, короста, рак кожи) – снижение активности УФ-эндонуклеазы; • Синдром Блума (глубокие поражения капилляров на лице) – мутация ДНК-лигазы; • Злокачественные перерождения – нарушение репарации неспаренных нуклеотидов. Пигментная ксеродерма Синдром Блума
 До 90% всех раковых заболеваний, возможно, связаны с нарушением репарации ДНК 6 -20 Essential Cell Biology (© Garland Science 2010
До 90% всех раковых заболеваний, возможно, связаны с нарушением репарации ДНК 6 -20 Essential Cell Biology (© Garland Science 2010
 Пигментная ксеродерма Возможно, снижение активности УФ-эндонуклеазы.
Пигментная ксеродерма Возможно, снижение активности УФ-эндонуклеазы.
 Синдром Блума мутация ДНК-лигазы
Синдром Блума мутация ДНК-лигазы
 Нобелевская премия по химии 2015 года за исследования механизмов репарации ДНК - Томас Линдаль, Пол Модрич и Азиз Санджар
Нобелевская премия по химии 2015 года за исследования механизмов репарации ДНК - Томас Линдаль, Пол Модрич и Азиз Санджар
 Генетическая информация реализуется • В пределах одной клетки • От родительской клетки к дочерним • Между клетками разных индивидуумов, в процессе размножения организмов
Генетическая информация реализуется • В пределах одной клетки • От родительской клетки к дочерним • Между клетками разных индивидуумов, в процессе размножения организмов
 Спасибо за внимание!
Спасибо за внимание!