молекулбиол -студ.pptx
- Количество слайдов: 99



Молекулярная биология Внутриклеточный поток генетической информации

• Жизнедеятельность клетки обеспечивается взаимосвязанными, упорядоченными в пространстве и во времени метаболическими и регуляторным процессами. • Клетка приобретает структуру и функции соответствующие этому уровню организации жизни благодаря потоку информации
 и")
• Непосредственными участниками потока информации являются: ЯДРО ( реакции репликации и транскрипции) и ЦИТОПЛАЗМА (реакции трансляции – образование полипептидов и посттрансляционные изменения белков (ФОЛДИНГ) приобретение белками вторичной, третичной или четвертичной структуры.

• В потоке наследственной информации ДНК выполняет ключевую роль: • 1. передача количественно и качественно полноценной наследственной информации в ряду клеточных поколений за счет воспроизводства (репликации) спиралей ДНК • 2. использование наследственной информации для организации клеточных функций путем синтеза белков

Направление потока генетической информации • 1. Синтез ДНК –репликация • 2. Синтез м РНК – транскрипция • 3. Синтез полипептидов трансляция

• РЕПЛИКАЦИЯ представляет собой симметричный процесс, который у эукариот протекает в синтетический период (S период) интерфазы. Обе молекулы биспирали выполняют роль матрицы для синтеза дочерних молекул в соответствии с правилами комплементарности. • Способ удвоения – полуконсервативный, Т. К. КАЖДАЯ вновь образовавшая биспираль состоит из одной материнской и одной дочерней молекул ДНК.

Полуконсервативный механизм репликации ДНК

Репликация у эукариотических организмов • ДНК эукариот удваивается не одним блоком от начала биспирали до ее конца, а отдельными участками – репликонами (около 1600 нуклеотидов). • В днк хромосом человека около 50 тысяч репликонов.

• РЕПЛИКОН – ЭТО УЧАСТОК МОЛЕКУЛЫ ДНК ОТ ТОЧКИ НАЧАЛА ОДНОЙ РЕПЛИКАЦИИ ДО ТОЧКИ НАЧАЛА ДРУГОЙ РЕПЛИКАЦИИ

• Репликация начинается в нескольких местах молекулы ДНК по 10 100 репликонов. Это сокращает время репликции ДНК. У эукариотических организмов этот процесс в среднем занимает 7 12 часов

РЕПЛИКОН

Задачи молекулярной биологии 1. КАК РАЗДЕЛЯЮТСЯ КОМПЛЕМЕНТАРНЫЕ ЦЕПИ, ЗАКРУЧЕННЫЕ ОДНА ВОКРУГ ДРУГОЙ? 2. КАКАЯ ФЕРМЕНТАТИВНАЯ СИСТЕМА ВОСПРОИЗВОДИТ ДНК С УЧЕТОМ АНТИПАРАЛЛЕЛЬНОСТИ ЕЕ ЦЕПЕЙ?

• Для репликации ДНК необходим большой набор разнообразных ферментов и белков — репликативный комплекс : • белки, раскручивающие спираль DNA, белки, стабилизирующие разделенные нити DNA, • белки, осуществляющие синтез дочерних молекул ДНК

Последовательность репликации ДНК • 1. ИНИЦИАЦИЯ - старт • 2. ЭЛОНГАЦИЯ- удлинение • 3. ТЕРМИНАЦИЯ- завершение

Инициация репликации 1. Инициация – начинается связыванием специфических «узнающих» белков с участками ДНК в точках инициации репликации 2. Сбрасывание супервитков молекулы ДНК называется РЕЛАКСАЦИЕЙ молекул (фермент топоизомераза) 3. Разделение (фермент ДНК – хеликаза) и удержание комплементарных цепей в расправленном состоянии (дестабилизирующий белок SSB) называется ЛОКАЛЬНОЙ ДЕНАТУРАЦИЕЙ или ПЛАВЛЕНИЕМ ДНК 4. образование репликативной вилки - репликативная вилка — участок ДНК, в пределах которого спираль раскручена и разделена на отдельные цепи. 5. синтез на матричной цепи ДНК затравочного полирибонуклеотидного короткого фрагмента (праймера – короткая молекула РНК)

Репликация ДНК

• Элонгация – матрицей для синтеза дочерней ДНК может служить только молекла ДНК, несущая спаренный с ней РНК праймер. Синтез новой цепи ДНК начинается от свободного 3 -ОН конца затравочного фрагмента РНК.

• Непрерывная репликация происходит только одной матричной цепи в направлении 5’ 3’ с образование ведущей (лидирующей) цепи • Отстающая (запаздывающая) цепь синтезируется ФРАГМЕНТАМИ ОКАЗАКИ в направлении 5’ 3’. Каждый фрагмент Оказаки инициируется коротким полирибонуклеотидом – затравкой

Репликативная вилка

• Репликация двунаправленная, вилки репликации удаляются друг от друга по мере синтеза ДНК до тех пор, пока не встретятся с вилками репликации других репликонов.

РЕПЛИКОН

• Терминация – состоит в удалении РНК праймеров и заполнении образовавшихся пустот в дочерней нити ДНК нуклеотидами. • Этот процесс «сшивания» фрагментов дочерней ДНК восстанавливает целостность молекулы. • У эукариот терминация репликации происходит при встрече репликативных вилок соседних репликонов.

БЕЛОК ФУНКЦИИ ТОПОИЗОМЕРАЗА Сбрасывание супервитков ДНК ХЕЛИКАЗА Раскручивание двойной спирали, денатурация ДНК ДЕСТАБИЛИЗИРУЮЩИЙ БЕЛОК SSB Стабилизация однонитевых разрывов БЕЛОК dna. B Инициация синтеза РНК затравки ДНК –ПРАЙМАЗА (РНК ПОЛИМЕРАЗА) Синтез РНК –затравок (праймеров) ДНК ПОЛИМЕРАЗА III Синтез ДНК –ПОЛИМЕРА I Удаление праймеров, заполнение однонитевых брешей ДНК ЛИГАЗА Ковалентное соединение фрагментов Оказаки

Репликация у прокариот • Бактериальная хромосома содержит один репликон. • ДНК реплицируется не прерываясь с образованием двух репликативных вилок в одной точке инициации. • Терминация репликации происходит при сязывании сайта TER – (короткая последовательност нуклеотидов) с продуктом гена TUS

у эукариот ДНК реплицируется не как таковая, а в виде хроматина, в котором она прочно связана с белками, принадлежащими к классу гистонов – нуклеосомами. • Нуклеосомы располагаются вдоль молекулы ДНК с интервалами 200 пар оснований. Этим объясняется тот факт, что новые фрагменты отстающей цепи ДНК закладываются у эукариот с интервалами в 10 раз более короткими (от 100 до 200 нуклеотидов), чем у бактерий (от 1000 до 2000 нуклеотидов). • Нуклеосомы служат барьерами, на время останавливающими продвижение ДНК-полимеразы, присутствие хроматина (а не голой ДНК) может, вероятно, объяснить и то, что репликационные вилки движутся у эукариот приблизительно в 10 раз медленнее, чем у бактерий. •

Репарация ДНК • Механизм репликации характеризуется высокой точностью, однако не исключены ошибки, которые приводят к нарушению правила комплементарности. • Самокоррекция (редактирование) ДНК текста позволяет найти ошибку и «выщипить» неправильный нуклеотид. • Поврежденные участки ДНК или ошибочно встроенные нуклеотиды удаляются в результате действия специальных эндо — и экзонуклеаз. • Образующиеся промежутки заполняются с помощью DNA—полимеразы и затем сшиваются лигазами с исходной нитью ДНК

СХЕМА РЕПАРАЦИИ ДНК

• Ген — единица наследственной информации, занимающая определенное положение в геноме или хромосоме и контролирующая выполнение определенной функции в организме.
 на")
транскрипция Транскрипция – процесс образования молекул РНК (и. РНК, р. РНК, т. РНК) на матричной полинуклеотидной цепи спирали ДНК

• Транскрипция- процесс переноса информации с двухцепочечной молекулы ДНК на одноцепочечные молекулы РНК. • МАТРИЦЕЙ для синтеза РНК служит только одна цепь ДНК, которая называется СМЫСЛОВОЙ ЦЕПЬЮ. • Другая цепь остается нетранскрибируемой

• Молекулы РНК считываются с определенных участков хромосомной ДНК, называемых «транскрипционными единицами» • Единица транскрипции транскриптон включает информативную часть, а также элементы необходимые для инициации, элонгации и терминации. • Синтез РНК транскрипта идет в направлении 5’ 3’

Эукариотический ген • Эукариотический ген можно рассматривать как совокупность сегментов ДНК, которые вместе составляют экспрессируемую единицу, ответственную за образование специфического функционального продукта — либо молекулы РНК, либо полипептида.

Стадии транскрипции • 1. Инициация • 2. Элонгация • 3. Терминация

• 1. Единица транскрипции – это участок ДНК, кодирующий первичный транскрипт. Он включает: а) последовательность, которая обнаруживается в зрелых функциональных молекулах РНК; б) интроны (для м. РНК); в) промежуточные последовательности спейсеры (для р. РНК). Интроны и спейсеры удаляются в ходе процессинга первичных транскриптов; г) 5' и 3' нетранслируемые последовательности (5' НТП и З' НТП).
 и конца транскрипции (терминатор).")
• 2. Минимальные последовательности, необходимые для начала транскрипции (промотор) и конца транскрипции (терминатор).

3. Последовательности, регулирующие частоту инициации транскрипции, ответственные за индуцибельность и репрессию транскрипции, а также клеточную, тканевую и временную специфичность транскрипции. Они разнообразны по строению, положению и функциям. • К их числу относятся энхансеры и сайленсеры это последовательности ДНК, расположенные в тысячах пар нуклеотидов от промотора эукариотического гена и оказывающие дистанционное влияние на его транскрипцию.

• многие гены эукариот имеют мозаичное строение. Под мозаичностью в данном случае подразумевается чередование кодирующих (экзоны) и некодирующих (вставочные последовательности, или интроны) последовательностей в пределах единицы транскрипции. Интроны чаще всего встречаются в генах, кодирующих белки.

Строение гена эукариот и прокариот

• Транскрибируемая часть генома человека составляет около 28 30% последовательностей нуклеотидов. • Из них только 5% принимают участие в потоке наследственной информации до молекул полипептидов. Эти участки ДНК называются экзонами. • 45% избыточных последовательностей нуклеотидов не транслируются в последовательность аминокислот и получили название ИНТРОНОВ. Интроны обязательно реплицируются и передаются в ряду поколений клеток

• В ходе синтеза РНК происходит расплетение примерно двух витков спирали ДНК. • В разных транскрипционных единицах ДНК матрицей для синтеза РНК могут служить разные комплементарные цепи. • Транскрипционные единицы ограничены с одной стороны участком инициации – промотором, с другой стороны – участком остановки транскрипции- терминатором.

Транскрипция ДНК с образованием м. РНК

МИНИМАЛЬНОЙ ФУНКЦИОНАЛЬНОЙ ЕДИНИЦЕЙ, КОДИРУЮЩЕЙ СТРОЕНИЕ ОДНОЙ ПОЛИПЕПТИДНОЙ ЦЕПИ является ЦИСТРОН

• Участок, с которым связывается РНК— полимераза, называется промотором, в его состав входят два участка консервативных последовательностей нуклеотидов (с постоянным составом и последовательностью) • 1. для узнавания РНК-полимеразы • 2 последовательность Прибноу – для тесного связывания РНК-полимеразы с промотором.

• Промотор относится к нетранслируемой части транскрипционной единицы. Функция промотора – создание условий для инициации транскрипции

Транскрипция генов эукариот • В клетках эукариот управляют синтезом р. РНК, м. РНК и т. РНК три различные РНК-полимеразы –А, В и С. • В результате транскрипции в ядре образуется м. РНК-предшественник, которая содержит как интроны, так и экзоны. • Перед перемещением из ядра интроны удаляются (процессинг), а экзоны соединяются друг с другом, сохраняя правильную последовательность кодонов (сплайсинг).
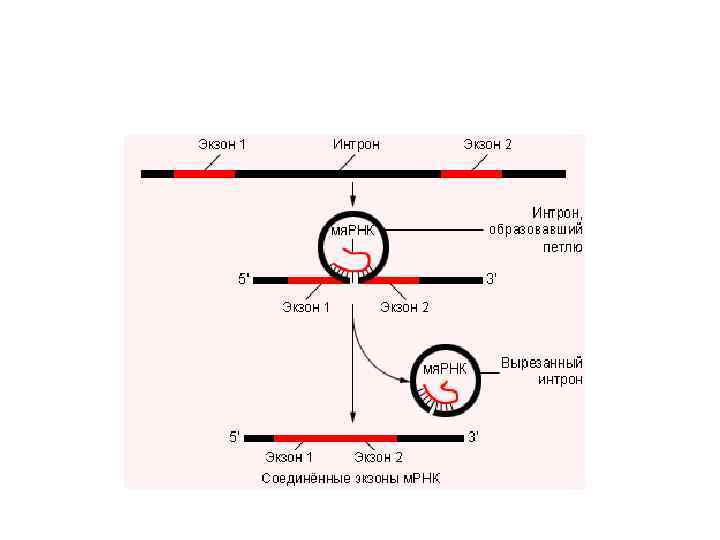

• В клетках многоклеточных эукариот происходит избирательная транскрипция генов: • 1. по месту – в клетках разных тканей • 2. по времени – в разные фазы клеточного цикла или онтогенеза особи • 3. по интенсивности – в зависимости от функционального состояния клетки.

Регуляция транскрипции генов у эукариот • каждый ген эукариот имеет собственный промотор и терминатор транскрипции • С промоторами эукариотических генов функционально связаны их специфические локусы – энхансены (усилители) и регуляторные элементы. • Энхансеры увеличивают число посадок РНК полимеразы на промотор гена в сотни раз. • Энхансеры тканеспецифичны

• Дополнительные регуляторные элементы присутствуют у индуцибельных генов, транксрибирующихся только при поступлении в клетку определенных веществ, они тесно сцеплены с промоторами. У высших многоклеточных организмов часто индукторами выступают молекулы гормонов.
 составляют повторяющиеся последовательности, имеющие определенную")
• Существенную часть генома эукариот (10 — 30%) составляют повторяющиеся последовательности, имеющие определенную структурную организацию и способные перемещаться в геноме как в пределах одной хромосомы, так и между хромосомами. Они получили название подвижных генетических элементов.

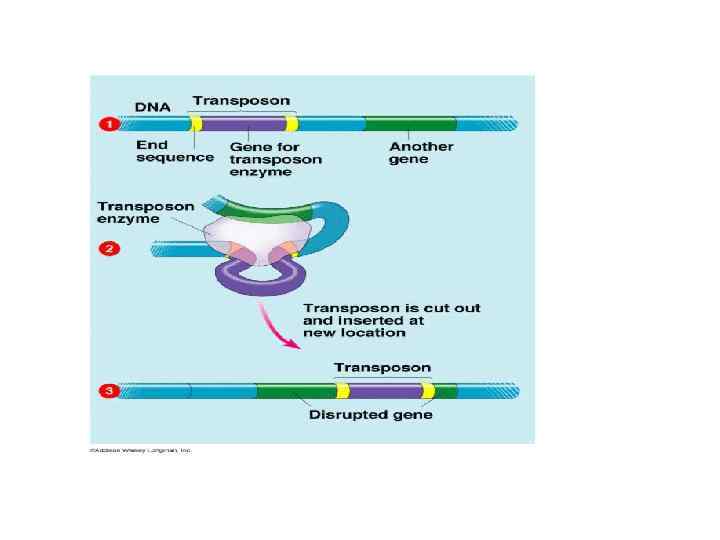
• Различают два основных класса подвижных генетических эле ментов: транспозоны и ретротранспозоны. Такая классификация основана на молекулярных механизмах, с помощью которых перемещаются эти элементы.
")
• Ретротранспозоны (мобильные генетические элементы первого типа, или транспозоны, перемещающиеся через РНК интермедиаты) — это генетические элементы, которые могут самовоспроизводиться в геноме и являются вездесущими компонентами ДНК многих эукариотических организмов.

Ретротранспозоны

Транспозоны • Транспозоны второго типа перемещаются по механизму «вырезать — вставить» , но не «копировать — вставить» , и используют при этом фермент транспозоназу. Разные транспозазы работают по разному. Некоторые связывают часть молекулы ДНК со случайным сайтом встройки, другие же связываются со специфическими последовательностями. [

Геномы органелл эукариот: ДНК митохондрий и хлоропластов Существуют два типа цитоплазматических ДНК: одни находят ся в митохондриях эукариот, другие в хлоропластах зеленых растений и водорослей. Как и все цитоплазматические элементы, они наследуются по материнской линии, а не по законам Менделя! Большая часть белков этих органелл, закодированная в ядерной ДНК, синтезируется в цитоплазме и затем переходит в органеллу. Однако некоторые белки митохондрий и хлоропластов и все их РНК кодируется в ДНК самих органелл и в них же синтезируют ся. Таким образом, органеллы — это результат объединенных усилий двух геномов и двух трансляционных аппаратов. РНК компо ненты рибосом органелл, а также т. РНК, использующиеся при трансляции, кодируются геномами митохондрий и хлоропластов.

Геном прокариот построен очень компактно. Количество некодирующих последовательностей нуклеотидов минимально. Многие механизмы регуляции экспрессии генов, использующиеся у эукариот, никогда не встречаются у прокариот. Простота строения генома прокариот объясняется их упрощенным жизненным циклом.
 В состав оперона входят: • промотор •")
Строение оперона (Ф. Жакоб, Ж. Моно, 1961) В состав оперона входят: • промотор • сайт связывания рибосом • инициирующий кодон • структурный ген • терминирующий кодон

• 5' конец прокариотического гена имеет характерную организацию регуляторных элементов, осо бенно на расстоянии 50 — 70 н. п. от точки инициации транскрип ции. Этот участок гена называют промотором. Он важен для транскрипции гена, но сам в РНК не транскрибируется. Противоположный 3' конец — терминаторная область, необходимая для терминации транскрипции. В РНК он также не транскрибируется.

Строение оперона

• Активность многих генов прокариот регулируется с помощью белковых факторов, взаимодействующих с регуляторными участками промоторов генов. При этом происходят как активация транскрипции генов, так и подавление считывания генетической информации РНК полимеразами. В первом случае регуляторные белковые факторы называют активаторами , осуществляющими позитивную регуляцию транскрипции , а во втором репрессорами. Регуляцию, связанную с подавлением транскрипции, называют негативной.
 — полицистронный оперон бактерий, кодирующий гены метаболизма лактозы.")
• Лактозный оперон (lac оперон) — полицистронный оперон бактерий, кодирующий гены метаболизма лактозы. • Регуляция экспрессии генов метаболизма лактозы у кишечной палочки (Escherichia coli) была впервые описана в 1961 году учеными Ф. Жакобом и Ж. Моно[1] (получившими в 1965 году Нобелевскую премию совместно с А. Львовым). Бактеральная клетка синтезирует ферменты, принимающие участие в метаболизме лактозы, лишь в том случае, когда лактоза присутствует в окружающей среде и клетка испытывает недостаток глюкозы.
 состоит из трех структурных генов, промотора, оператора и")
• Лактозный оперон (lac operon) состоит из трех структурных генов, промотора, оператора и терминатора. Принимается, что в состав оперона входит также ген регулятор, который кодирует белок репрессор. • Структурные гены лактозного оперона — lac. Z, lac. Y и lac. A: • lac. Z кодирует фермент β галактозидазу, которая расщепляет дисахарид лактозу на глюкозу и галактозу, • lac. Y кодирует β галактозид пермеазу, мембранный транспортный белок, который переносит лактозу внутрь клетки. • lac. A кодирует β галактозид трансацетилазу, фермент, переносящий ацетильную группу от ацетил Кo. A на бета галактозиды. • Для катаболизма лактозы необходимы только продукты генов lac. Z и lac. Y, роль продукта гена lac. A не ясна.

Случай, когда есть глюкоза и нет лактозы

• РНК полимераза начинает транскрипцию с промоторного района, который перекрывается с операторным районом. В отсутствие или при низкой концентрации лактозы в клетке белок репрессор, который является продуктом моноцистронного оперона Lac. I, обратимо соединяется с операторным районом и препятствует транскрипции. Таким образом, в отсутствие лактозы в клетке ферменты для метаболизма лактозы не синтезируются. • Данный механизм регуляции активности лактозного оперона называют позитивной индукцией. Веществом индуктором служит лактоза; при её связывании с белком репрессором происходит его диссоциация от операторного участка.

Случай, когда есть глюкоза и нет лактозы

Если в клетке концентрация глюкозы достаточная для поддержания метаболизма, активация лактозного оперона не происходит. Промоторная последовательность лактозного оперона «слабая» , поэтому даже при отсутствии белка репрессора на операторном участке транскрипция практически не инициируется

Случай, когда нет глюкозы и нет лактозы

Случай, когда нет глюкозы и есть лактоза

• . В присутствии лактозы происходит экспрессия генов оперона. Данный механизм регуляции активности лактозного оперона называют негативной индукцией. «Негативным индуктором» служит глюкоза, которая подавляет активность лактозного оперона.

• Транскрипцию у бактерий осуществляет одна, общая для всех генов РНК-полимераза

• У прокариот трансляция м. РНК начинается с посадки рибосом, которая осуществляется благодаря комплементарности рибосомной РНК и лидерной части м. РНК. • Рибосомы считывают информацию со всех генов полигенной м. РНК • Перед каждым геном свой сайт связывания рибосом, инициирующий кодон и терминирующий кодон. • На каждом гене м. РНК синтезируется своя отдельная полипептидная цепь.

• ферменты для усвоения лактозы синтезируются в клетке кишечной палочки при двух условиях: • 1) наличие лактозы; • 2) отсутствие глюкозы. • Регуляция работы лактозного оперона в зависимости от концентрации лактозы происходит по принципу отрицательной обратной связи: • чем больше лактозы — тем больше ферментов для её катаболизма (положительная прямая связь); • чем больше ферментов — тем меньше лактозы, чем меньше лактозы — тем меньше производится ферментов (двойная отрицательная обратная связь).

• репликативная вилка — участок ДНК, в пределах которого спираль раскручена и разделена на отдельные цепи. • Репликация начинается в нескольких местах молекулы ДНК. • УЧАСТОК МОЛЕКУЛЫ ДНК ОТ ТОЧКИ НАЧАЛА ОДНОЙ РЕПЛИКАЦИИ ДО ТОЧКИ НАЧАЛА ДРУГОЙ РЕПЛИКАЦИИ НАЗЫВАЕТСЯ – РЕПЛИКОН • Бактериальная хромосома содержит один репликон, эукариотическая – много репликонов

• Непрерывная репликация происходит только одной матричной цепи в направлении 5’ 3’ с образование ведущей (лидирующей) цепи • Отстающая (запаздывающая) цепь синтезируется ФРАГМЕНТАМИ ОКАЗАКИ в направлении 5’ 3’. Каждый фрагмент Оказаки инициируется коротким полирибонуклеотидом – затравкой • После удаления затравки фрагменты ОКАЗАКИ сшиваются ковалентно.

Генетика – наука, которая изучает универсальные для всех живых существ закономерности –НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ • Наследственность – свойство живых систем сохранять из поколения в поколение сходные признаки и обеспечивать специфический характер индивидуального развития в определенных условиях среды • Изменчивость – свойство живых систем приобретать новые признаки, отличающие их от родительских форм

УРОВНИ ОРГАНИЗАЦИИ НАСЛЕДСТВЕННОГО МАТЕРИАЛА 1. Генный 2. Хромосомный 3. Геномный

ГЕН – единица наследственности и изменчивости • Ген является функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма. • Передача генов в ряду поколений клеток обеспечивает материальную преемственность – наследование потомками признаков родителей.

ГЕН – единица наследственности и изменчивости • Ген является функциональной единицей генетического аппарата, определяющей возможность развития отдельного признака клетки или организма. • Передача генов в ряду поколений клеток обеспечивает материальную преемственность – наследование потомками признаков родителей.

МАТЕРИАЛЬНЫМ СУБСТРАТОМ НАСЛЕДСТВЕННОСТИ И ИЗМЕНЧИВОСТИ являются НУКЛЕИНОВЫЕ КИСЛОТЫ – полимерные молекулы, состоящие из нуклеотидов.

Строение ДНК

Принцип комплементарности

СВОЙСТВА ГЕНА • 1. Дискретность – определяет возможность развития отдельного качества, присущего данной клетке • 2. Целостность при программировании синтеза полипептида выступает как неделимая частица) • 2. Специфичность – зашифрована информация определенного полипептида • 3. Плейотропность множественность действия • 4. Дозированность действия – количественное выражение признака в зависимости от дозы соответствующего аллеля этого гена • 5. Изменчивость – минимальной единицей изменчивости гена является МУТОН • 6. Стабильность – относительная устойчивость

ПОТОК ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ • 1. Последовательность нуклеотидов в ДНК определяет структуру всех белков клетки • 2. Участки ДНК, кодирующие определенные полипептиды (гены) копируются (транскрибируются) в виде полинуклеотидной цепи м РНК. • 3. м РНК служит матрицей для синтеза белка – генотип трансформируется в фенотип

• Для репликации ДНК необходим большой набор разнообразных ферментов и белков — репликативный комплекс : белки, раскручивающие спираль DNA, и белки, стабилизирующие разделенные нити DNA.

Уровни компактизации наследственного материала
 • Разделение (фермент")
• Сбрасывание супервитков молекулы ДНК называется РЕЛАКСАЦИЕЙ молекул (фермент топоизомераза) • Разделение (фермент ДНК – хеликаза) и удержание комплементарных цепей в расправленном состоянии (дестабилизирующий белок SSB) называется ЛОКАЛЬНОЙ ДЕНАТУРАЦИЕЙ или ПЛАВЛЕНИЕМ ДНК

• Бактериальная хромосома содержит один репликон, эукариотическая – много репликонов
 происходит по полуконсервативному механизму. Каждая нить двойной спирали")
Репликация ДНК • Репликация ДНК(воспроизведение генотипа) происходит по полуконсервативному механизму. Каждая нить двойной спирали выступает в роли матрицы для синтеза новой цепи. Следовательно, вновь образованные двухспиральные молекулы состоят из одной новой и одной старой цепи.

Последовательность репликации ДНК • 1. Инициация – синтез на матричной цепи ДНК затравочного полирибонуклеотидного короткого фрагмента (праймера) • 2. Элонгация – синтез новой цепи ДНК начинается от свободного 3 ОН конца затравочного фрагмента РНК. • В процессе элонгации происходит удаление праймера • 3. Репликация двунаправленная, вилки репликации удаляются друг от друга по мере синтеза ДНК до тех пор, пока не встретятся с вилками репликации других репликонов.

• Непрерывная репликация происходит только одной матричной цепи в направлении 5’ 3’ с образование ведущей (лидирующей) цепи • Отстающая (запаздывающая) цепь синтезируется ФРАГМЕНТАМИ ОКАЗАКИ в направлении 5’ 3’. Каждый фрагмент Оказаки инициируется коротким полирибонуклеотидом – затравкой • После удаления затравки фрагменты ОКАЗАКИ сшиваются ковалентно.

Репликация ДНК

БЕЛОК ФУНКЦИИ ТОПОИЗОМЕРАЗА Сбрасывание супервитков ДНК ХЕЛИКАЗА Раскручивание двойной спирали, денатурация ДНК ДЕСТАБИЛИЗИРУЮЩИЙ БЕЛОК SSB Стабилизация однонитевых разрывов БЕЛОК dna. B Инициация синтеза РНК затравки ДНК –ПРАЙМАЗА (РНК ПОЛИМЕРАЗА) Синтез РНК –затравок (праймеров) ДНК ПОЛИМЕРАЗА III Синтез ДНК –ПОЛИМЕРА I Удаление праймеров, заполнение однонитевых брешей ДНК ЛИГАЗА Ковалентное соединение фрагментов Оказаки

• Транскрипция- процесс переноса информации с двухцепочечной молекулы ДНК на одноцепочечные молекулы РНК. • МАТРИЦЕЙ для синтеза РНК служит только одна цепь ДНК, которая называется СМЫСЛОВОЙ ЦЕПЬЮ. • Другая цепь остается нетранскрибируемой

Стадии транскрипции • 1. Инициация • 2. Элонгация • 3. Терминация

• В ходе синтеза РНК происходит расплетение примерно двух витков спирали ДНК. • В разных транскрипционных единицах ДНК матрицей для синтеза РНК могут служить разные комплементарные цепи. • Транскрипционные единицы ограничены с одной стороны участком инициации – промотором, с другой стороны – участком остановки транскрипции- терминатором.

ГЕНЕТИЧЕСКАЯ СИСТЕМА КООРДИНАТ • НУКЛЕОТИДЫ НУМЕРУЮТСЯ ОТ ГРАНИЦЫ МЕЖДУ ТРАНСКРИБИРУЕМОЙ И НЕТРАНСКРИБИРУЕМОЙ ОБЛАСТЯМИ ДНК • Первый транскрибируемый нуклеотид и все нуклеотиды, находящиеся от него справа, имеют номера со знаком (+) • Нетранскрибируемые нуклеотиды нумеруются влево со знаком (-) • Состав нуклеотидов в гене указывается не по смысловой цепи, а по комплементарной цепи.

Транскрипция ДНК с образованием м. РНК

нуклеосома
молекулбиол -студ.pptx