08 Репликация (1).pptx
- Количество слайдов: 44



Молекулярная биология Лекция 7. Репликация и репарация. Скоблов Михаил Юрьевич

Часть 1. Репликация ДНК

Эксперимент Мезельсона и Сталя 1958 год

ДНК-полимераза • В 1956 г. Корнберг выделил из клеток бактерии Е. coli фермент ДНК-полимеразу (ДНК-полимераза I). • Этот фермент осуществлял синтез ДНК при наличии в реакционной смеси всех четырех дезоксинуклеозидтрифосфатов: АТФ, ГТФ, ТТФ, ЦТФ и молекулы ДНК • В 1959 г. получил нобелевскую премию «За открытие механизмов биологического синтеза рибонуклеиновой и дезоксирибонуклеиновой кислот» . Артур Корнберг

Репликация ДНК — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. «Каждая цепь двуцепочечной ДНК служит матрицей при синтезе комплементарной цепи и в результате образуются две пары цепей, в каждой из которых только одна является родительской» – Уотсон и Крик. Основные этапы репликации: 1. Инициация 2. Элонгация 3. Терминация

Основные ферменты репликации • ДНК-полимераза - фермент катализирующий полимеризацию дезоксирибонуклеотидов на матрице ДНК по принципу комплементарности • ДНК-лигаза – фермент катализирующий образование фосфодиэфирных связей между 5'-фосфорильной и 3'гидроксильной группами соседних дезоксинуклеотидов в местах разрыва двуцепочечной ДНК • ДНК-хеликаза – фермент разделяющий цепи двухцепочечной ДНК на одинарные. • ДНК-топоизомераза - фермент изменяющий степень сверхспиральности ДНК, путем внесения одноцепочечных разрывов в ДНК. • ДНК-праймаза — это фермент РНК-полимераза, синтезирующий короткий фрагмент РНК, называемый праймером, комплементарный одноцепочечной матрице ДНК.

Ориентация цепей в ДНК
 separates a ds. DNA")
Репликация ДНК у E. coli STEP 1: Dna. B helicase(blue) separates a ds. DNA into two ss. DNAs, as cutting the hydrogen bands between base pairs. Topoisomerase(lime green) has a role in rewinding the twist of double helices which was generated by the ds. DNA separation. STEP 2: The separated ss. DNA has a tendency of annealing. For preventing annealing, Single-Stranded DNA Binding Protein: SSB bind to separated ss. DNA. STEP 3: Dna. G primase(purple) is activated by binding to Dna. B helicase, and synthesizes a short RNA primer approximately 10 nucleotides long using a ss. DNA as a template. STEP 4: DNA polymerase III elongates a new ss. DNA strand by adding a deoxyribonucleotide at a time in the 5'-3' direction to the RNA primer, using a ss. DNA as a template.

Репликация ДНК у E. coli • Лидирующая цепь удлиняется ДНКполимеразой в направлении 3’-конца • Отстающая цепь удлиняется по иному механизму:

Репликация ДНК у E. coli

Репликации у прокариот Инициация репликации • Репликация ДНК начинается в специфических местах называемых точками начала репликации (origin). • Чем больше размер генома организма, тем большее количество точек репликации ему необходимо: у E. coli, бактериофага лямбда, вируса SV 40 – один origin, в эукариотических клетках – множественные начала репликации расположеные на расстоянии 20 т. п. н. . Элонгация • Скорость репликации у бактерий порядка 100 000 пар нуклеотидов в минуту, у эукариот - 500— 5000. • Репликация E. coli занимает около 40 минут, в то же время клетки делятся примерно каждые 20 минут – репликация ДНК инициируется при не законченной предыдущей.

Репликации у прокариот Терминация репликации Топоизомераза II также принимает участие в рахсождении двух дочерних кольцевых ДНК - конкатамеров

Репликация ДНК по принципу катящегося кольца Одноцепочечная ДНК Двухцепочечная ДНК

Репликации у эукариот Ori - AT-богатые, легкоплавкие участки длинной ~200 пн, расположенные, как правило, между генами в промоторных областях.
")
Репликация теломер Теломе ры (от др. -греч. τέλος — конец и μέρος — часть) — концевые участки хромосом Нобелевская премия по физиологии и медицине 2009 года американских учёных Элизабет Блэкбёрн (Elizabeth H. Blackburn), Кэрол Грейдер (Carol W. Greider) и Джек Шостак (Jack W. Szostak) «за открытие того, как теломеры и фермент теломераза защищают хромосомы»

Репликация теломер • При каждом клеточном делении теломеры теряют примерно 100 оснований • По расчетам при такой скорости приблизительно после 125 митотических делений теломеры должны полностью исчезнуть, что приведёт к клеточной гибели. • В 1961 году Хейфлик определил границу количества делений соматических клеток , которая для большинства человеческих клеток составляет 52 деления. Это явление получило название "лимит Хейфлика". • При искусственной оверэкспрессии теломеразы в клетках, они начинают делится порядка 100 раз.
 к")
Репликация теломер Теломераза — фермент, добавляющий особые повторяющиеся последовательности ДНК (TTAGGG у позвоночных) к 3'-концу цепи ДНК на участках теломер, которые располагаются на концах хромосом в эукариотических клетках.

Репликация теломер • В работе использовали образцы ДНК 356 мужчин и 551 женщин, с возрастом от 18 до 92 лет. • Как и ожидалось длина теломер негативно коррелировала с возрастом (r = -0. 40; P < 0. 001). При этом не было значительной разницы в данных для мужчин и женщин. • Также была обнаружена позитивная корреляция между длинной теломер у отцов и детей (r = 0. 46, P < 0. 001; beta = • 0. 22, P = 0. 006), в то же время корреляция между длинной теломер у матерей и детей была меньше (r = 0. 18, P = 0. 04). Также была найдена позитивная корреляция между длинной теломер у дочерей и продолжительностью жизни отцов (r = 0. 20, P < 0. 001), но не между длинной теломер у дочерей и продолжительностью жизни матерей (r = -0. 01, P = not significant). Telomere length is paternally inherited and is associated with parental lifespan. Njajou OT, Cawthon RM, Damcott CM, Wu SH, Ott S, Garant MJ, Blackburn EH, Mitchell BD, Shuldiner AR, Hsueh WC. Proc Natl Acad Sci U S A. 2007 Jul 17; 104(29): 12135 -9.
 теломераза активна,")
Теломеры и теломераза в онкогенезе • В большинстве опухолевых клеток (80 -90%) теломераза активна, в остальных случаях считается, что клетки могут использовать альтернативный механизм поддержания длины теломер, основанный на рекомбинации. • Опухолевые клетки чрезвычайно гетерогенны, и представляют собой популяцию клеток, которая активно делится, при этом у них укорачиваются теломеры, но затем выживают те из них, у которых активируется теломераза. • Отсюда получается, что активность теломеразы может служить неким маркёром опухолевой прогрессии.

Теломеры и теломераза в онкогенезе • Если в исследуемом типе клеток теломераза была активна, то при трансформации в опухолевую клетку эта активность только увеличивается. • Механизмы активации теломеразы: • Амплификация генов • Эпигенетически: • Метелирование Cp. G островка в промоторной области гена теломеразы • модификации гистонов • Активация онкогеном Myc • Транскрипционными факторами Sp 1 и SP 3 • Активация онкосупрессорами Р 53 и Р 73 • Активация вирусами

Клеточный цикл и его регуляция • Период существования клетки от одного деления до другого называется митотическим или клеточным циклом. • У высших организмов он занимает от 10 до 30 часов, у человека 24 часа • У бактерий – менее часа, около 20 минут

Разница в репликации у прокариот и эукариот

Часть 2. Репарация

Источники повреждения ДНК • У бактерии кишечной палочки E. coli известно более 50 генов, контролирующих процессы репарации. • В среднем в процессе репликации генома млекопитающего длинной 3 млрд нуклеотидов возникает не более 3 ошибок • Ежедневно в молекулах ДНК каждой клетки человеческого тела около 100000 звеньев повреждаются за счет разнообразных эндогенных процессов и экзогенных генотоксичных воздействий. • Менее 1 повреждения ДНК из 1000 превращается в мутацию.

Репарация ДНК • Репарация генетических повреждений – свойство живых организмов восстанавливать нарушения и повреждения, возникшие в ДНК в результате ошибок репликации, а также при воздействии разнообразных эндогенных и внешних мутагенных факторов. • Повреждение ДНК – это не мутация. • Мутация – это наследственное (фиксированное) изменение в нуклеотидной последовательности генома организма.

Основные повреждения ДНК

Наследственные заболевания • • Частота генетических заболеваний варьируется от 1: 10 000 до 1: 1 000 Бывают и уникальные заболевания, 1 на 6 000 000 Общая частота генетических заболеваний в популяции составляет около 3% В арабских странах около 6% (за счет близкородственных браков) Генетический груз популяции каждый человек является носителем в среднем трех аутосомно -рецессивных аллелей

Болезни ассоциированые с дефектами системы репарации

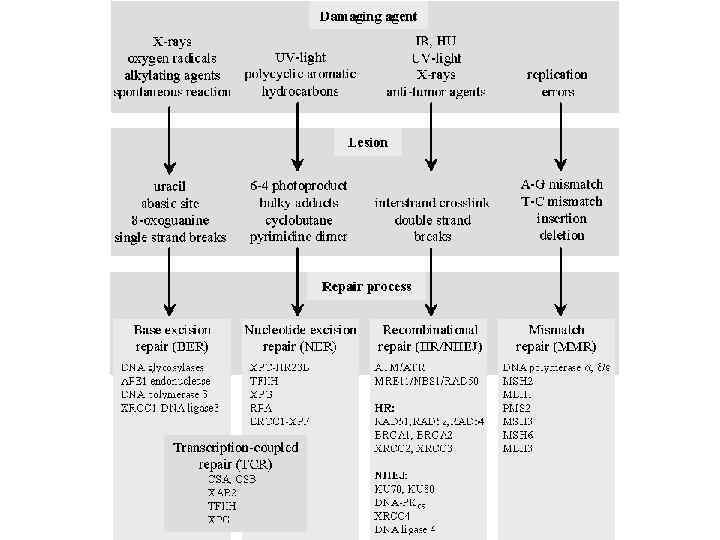
Репарация ДНК 1. Репарация неспаренных оснований. 2. Восстановление исходной структуры. 3. Эксцизионная репарация • Вырезание оснований • Вырезание нуклеотидов. 4. Пострепликативная репарация • Рекомбинационная репарация • SOS-репарация – мутагенный или «ошибочный» путь репарации.
 или 6")
Прямая репарация • В следствии УФ-излучения возникают пиримидиновые димеры (часто тиминовые димеры) или 6 -4 -фотопродукт • В основном они блокируют транскрипцию, и только небольшая часть приводит к мутациям. Тиминовый димер 6 -4 -фотопродукт

Фотореактивация • В фотореактивации участвует фермент фотолиаза, мономерный флавинзависимый фермент, и кофакторы : FADH- и 5, 10 -метенилтетрагидрофолат (5, 10 -MTHF) • Фотолиаза связывается в темноте с димерами ТТ • На свету кофактор абсорбирует фотон • Используя эту энергию фотолиаза расщепляет ТТ димер • Фотолиаза освобождает ДНК

Фотолиазы • Принадлежат большому семейству фотолиазкриптохромов. • Представители этого семейства широко распространены во всех царствах • У растений криптохромы регулируют рост, регулируемый синим светом, а у животных – циркадные ритмы.
 Nucleotide excision repair (NER)")
Эксцизионная репарация Mismatch repair Base excision repair (BER) Nucleotide excision repair (NER)
 Мисматч репарация: выявляет некомплементарную пару только на дочерней цепи ДНК и")
Mismatch repair (MMR) Мисматч репарация: выявляет некомплементарную пару только на дочерней цепи ДНК и производит замену неправильного основания только на дочерней цепи.
 Перед репликацией ДНК находится в метилированной форме, вновь синтезированная цепь -")
Mismatch repair (MMR) Перед репликацией ДНК находится в метилированной форме, вновь синтезированная цепь - неметилирована
 Основные белки метил-направляемой MMR в E. Coli: • Mut S и")
Mismatch repair (MMR) Основные белки метил-направляемой MMR в E. Coli: • Mut S и Mut L узнают ММ • Mut H - • Узнает полуметилированный • сайт GATC и делает надрез • Mut. U (Uvr. D) –геликаза II раскручивает дуплекс и освобождает надрезанную область

Mismatch repair у человека

Mismatch repair у человека • Система MMR у эукариот организована сложнее функционирует эффективнее по сравнению с бактериями. • У эукариот MMR исправляет все некомплементарные пары оснований и, кроме того, репарирует делеции или инсерции в рекомбинационных гетеродуплексах размером до 12 н. • У бактерий MMR неспособна исправлять пары С*С и репарирует делеции/инсерции не более 3 н. в рекомбинационных гетеродуплексах. • Ключевые белки MMR – Mut. L и Mut. S высококонсервативны, их гомологи обнаружены у всех организмов от E. coli до человека. • Если у E. coli эти белки (и кодирующие их гены) уникальны, то у эукариот имеется по несколько их гомологов (паралогов). Например, у дрожжей Saccharomyces cerevisiae обнаружены 3 гомолога Mut. L и 6 гомологов Mut. S, у человека – 11 гомологов Mut. L и 4 Mut. S.

Эксцизионная репарация Base excision repair - BER • ДНК гликозилазы, распознают аномальные основания ДНК и катализируют гидролитическое расщепление N-glycosyl связи, между основанием и сахаром. • Образуется AP-сайт (apurinic/apyrimidinic) • АП-сайт распознается АП-эндонуклеазой, которая вводит в нить ДНК разрыв. • Фосфодиэстераза отщепляет от ДНК сахарофосфатную группу, к которой не присоединено основание. • Брешь размеров в 1 н. застраивается ДНК -полимеразой I и концы ДНК соединяются ДНК-лигазой. В каждой клетке млекопитающих за одну 20 -ти часовую генерацию спонтанно возникает около 10000 апуриновых сайтов и около 500 – апиримидиновых.

Эксцизионная репарация Nucleotide excision repair - NER • Узнавание повреждений • Связывание мультисубъединичного комплекса с поврежденным сайтом • Двойное надрезание поврежденной цепи на несколько нуклеотидов от поврежденного сайта в обоих направлениях 5' и 3' • Освобождение олигонуклеотида, содержащего повреждение между двумя надрезами • Заполнение образовавшейся бреши ДНК полимеразой • Лигирование

Эксцизионная репарация • У эукариот система эксцизионной репарации нуклеотидов функционирует по той же схеме, что и у бактерий, но организована сложнее и работает эффективнее, по сравнению с бактериями. • Эукариотическая эксцинуклеаза включает, по крайней мере 17 белков, и при эксцизии вырезаются 29 н.

Пострепликативная репарация • Этот способ восстановления целостности ДНК заключается в репарации пробелов, образующихся в дочерних цепях напротив не удаленных в ходе репликации димеров. • Основная часть таких пробелов репарируется путем рекомбинационных обменов между двумя сестринскими цепями. В процессе используются ферменты ДНКполимераза I и лигаза, и белок Rec. A.

SOS-репарация • Ключевая роль в SOS-индукции принадлежит белку Rec. A, который связывается с белком SSB и с однонитевой ДНК, что является сигналом, запускающим индукцию SOS-регулона (около 30 генов), продукты которых необходимы для выживания клетки при массовых повреждениях ДНК. • Такая репликация является неточной, склонной к ошибкам, что приводит к повышению частоты мутаций.
08 Репликация (1).pptx