Синапсы и их физиологические свойства.pptx
- Количество слайдов: 60



МОДУЛЬ IV. ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ И ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ д. б. н. , профессор Дарья Захаровна Шибкова

План: 1. Краткая история изучения вопроса. 2. Классификация синапсов. 3. Строение и механизм передачи возбуждения в электрических синапсах. 4. Особенности проведения возбуждения в химических синапсах. 5. Медиаторы, их многообразие и механизмы выделения. 6. Понятие о потенциале концевой пластинки, миниатюрном потенциале и ВПСП. 7. Механизм пресинаптического и постсинаптического торможения. 8. Физиологические свойства синапсов.
 – в переводе с греческого означает контакт. Каждая нервная клетка представляет собой")
Синапс (synapsis) – в переводе с греческого означает контакт. Каждая нервная клетка представляет собой отдельную функциональную единицу, следовательно, существуют механизмы, которые обеспечивают передачу возбуждения с одного нейрона на другой или на эффекторную клетку, например, мышечное волокно. Передача осуществляется через синаптическую связь.

Идея о синаптической передаче была высказана еще Галеном, который указал, что мышца сокращается в результате воздействия со стороны нерва; этот вопрос продолжал исследовать Клод Бернар (1844), доказавший парализующее действие кураре на уровне двигательной пластинки. Само понятие синапс было предложено английским физиологом Шеррингтоном (1897), и означает оно место контакта двух клеток.

Приоритет в учении о синапсах принадлежит отечественной школе физиологов. Д. С. Воронцов – исследовал электрической межнейрональной передачи. механизм И. О. Коштоянц – химические вещества не только передают возбуждение, но также влияют на регуляцию процессов жизнедеятельности. Г. И. Гинецинский – изучал потенциал действия концевой пластинки, показал, что яды парализуют холинэстеразу, способствуют накоплению в области синапса ацетилхолина.

А. В. Кибяков – роль ацетилхолина в передаче возбуждения через симпатические ганглии. А. Ф. Самойлов – химический мышечной передачи (1924). этап нервно- В. К. Луценко – исследует молекулярные механизмы патологии синапса. Т. М. Турпаев – идентифицировал холинорецептор с белком.

Синапсы классифицируются по структурным элементам: Аксонодендрические Аксосоматические Аксоаксональные

По механизму передачи возбуждения: Электрические Химические Смешанные или синапсы двоякого действия

Синапсы с электрическим механизмом передачи чаще встречаются у животных с примитивной нервной системой, в основном у беспозвоночных, а в некоторых случаях встречаются в нервной системе позвоночных. Число электрических синапсов уменьшается в процессе эмбрионального развития. У высших животных и человека большую часть синаптического аппарата ЦНС составляют синапсы с химическим механизмом передачи.

Критерии, доказывающие принадлежность синапса к электрическому или химическому типу, можно разделить на 3 группы: Электрофизиологические Морфологические Фармакологические

Основным электрофизиологическим критерием электрических синапсов является наличие электрической связи с пре- и постсинаптическими клетками. Обычно эту связь исследуют путем пропускания тока через мембрану постсинаптической клети и регистрации возникающего при этом изменения мембранного потенциала пресинаптической клетки. В химических синапсах пропускание тока через мембрану пресинаптической клетки вызывает деполяризацию постсинаптической мембраны.

В электрических синапсах передача возбуждения может быть двусторонняя, либо односторонняя. Кроме того, интервал времени между началом пресинаптического потенциала действия и постсинаптической деполяризации составляет 0, 05 -0, 1 мс. Это значительно меньше, чем в химических синапсах.

Морфологическими критериями электрических синапсов являются: v отсутствие или очень небольшая ширина синаптической щели (меньше, чем в химических синапсах); v наличие специализированных структур – щелевых контактов. Фармакологические критерии – отсутствие чувствительности электрической передачи к специфическим блокаторам хеморецепторов постсинаптической мембраны.

Электронно-микроскопическими исследованиями с применением методики сколов замороженных срезов было показано, что в месте электрического синапса обнаружены гидрофильные каналы, соединяющие обе клетки. Через гидрофильные каналы, по-видимому, ряд веществ может проходить из одной клетки в другую.

Рис. Схема щелевого контакта: М 1 и М 2 – мембраны соседних клеток; ЩК – щелевой контакт; Щ – щель (20 – 30 нм); ГК – гидрофильные каналы, проходящие через обе мембраны и щель; ГС – глобулярные структуры.

Тесное соединение мембран делает возможным электрическую передачу импульса, т. к. в области щелевого контакта резко уменьшено сопротивление электрическому току при его прохождении через синаптическую щель. Это позволяет значительной части тока, генерируемого пресинаптической структурой, проникать в постсинаптическую клетку. Электрическая реакция в мембране постсинаптической клетки начинается немедленно, как только импульс приходит в пресинаптическое окончание.

Физиологическая роль электрических синапсов Сравнение функций электрических синапсов у животных различных систематических групп показывает, что они обычно входят в такие пути нервной системы, которые обслуживают срочные и наиболее быстрые реакции организма, например реакции защиты. Так речной рак осуществляет быструю реакцию подгибания брюшка, позволяющую животному мгновенно уплыть в направление «задомна-перед» .

Химические синапсы Идея о возможности химической передачи активности от одной клетки на другую впервые была высказана австрийским физиологом О. Леви в 1921 году для синаптических соединений, образуемых окончанием блуждающего нерва в сердце. О. Леви показал, что при раздражении блуждающего нерва в сердце появляется особое химическое вещество, которое оказывает на сердечную мышцу такое же действие, как и естественный нервный импульс. Этим веществом, как показали дальнейшие исследования, является ацетилхолин.

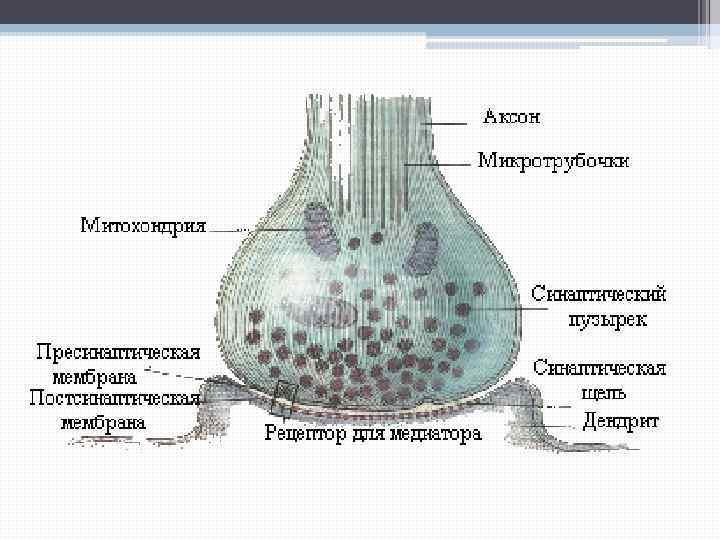
Позже было показано, что и в таких широко распространенных периферических соединениях, как нервно-мышечные синапсы скелетной мускулатуры, механизм передачи активности тоже является химическим и основан на выделении ацетилхолина. В настоящее время детально изучена структура нервно-мышечного синапса. Подходя к мышечному волокну, диаметр которого составляет 100 – 150 мкм, нервное волокно теряет миелиновую оболочку и разветвляется на его поверхности. Концевые безмиелиновые веточки диаметром до 1, 5 мкм ложатся в желобки мышечного волокна, и между ними образуется первичная синаптическая щель.
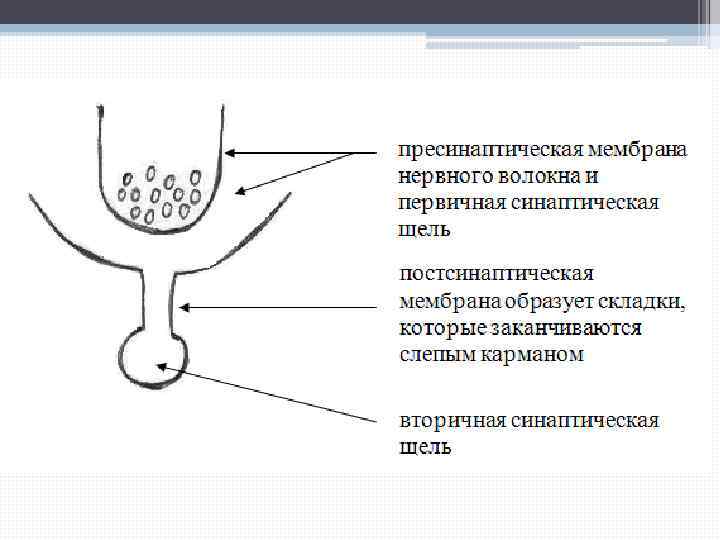

В аксоплазме нервного окончания находят множество синаптических пузырьков или везикул, которые скапливаются у утолщенных участков пресинаптической мембраны – так называемых активных зон. Между плазматическими мембранами нервного окончания и мышечного волокна имеется промежуток от 50 до 100 нм – первичная синаптическая щель.

В щели обнаруживают базальную мембрану – которая является производным постсинаптической мембраны. Поверхность мышечного волокна, прилегаемая к нервному окончанию в области синапса, называется концевой или двигательной пластинкой. Эта поверхность образует складки, которые заканчиваются слепыми карманами – это вторичная синаптическая щель.

Скопления везикул пресинаптической мембраны располагаются прямо над вторичной синаптической щелью, т. е. положение активных центров пресинаптической мембраны совпадает с расположением карманов на постсинаптической мембране.

Весь синапс одевается шванновскими клетками, которые дают отростки. Эти отростки проникают в первичную синаптическую щель и делят ее на отдельные сегменты. Каждый такой сегмент содержит 1 – 2 активных центра. В среднем в одном синапсе насчитывается до 1 – 2 тысяч активных центров.

Нейрон и его компоненты. А — нервная клетка, аксон, мышца; Б — строение нервного волокна

Строение нервномышечного синапса. А — нервномышечный синапс в разрезе; Б — распределение синапсов соматического аксона в волокнах скелетной мышцы; В — распределение синапсов симпатического аксона в клетках гладкой мышцы

В пресинаптической части находится фермент холинацетилаза, который участвует в синтезе ацетилхолина. Катионная головка ацетилхолина, соединенная с остатком уксусной кислоты Активностью обладает катионная головка ацетилхолина.

Условия, необходимые для выделения ацетилхолина: - чтобы произошло освобождение ацетилхолина, везикулы должны провзаимодействовать с пресинаптической мембраной. Мембрана на этом участке должна быть комплементарна везикуле. - вторым условием необходимым для выхода ацетилхолина является вход Са 2+ из синаптического пространства внутрь пресинаптического волокна, где Са 2+ находится в концентрации 10 -7 моль, если возникает возбуждение, концентрация Са 2+ увеличивается до 10 -4 моль.

Кальций снижает энергетический барьер для слияния везикул с пресинаптической мембраной, в результате этого слияния везикулы лопаются и ацетилхолин изливается в первичную синаптическую щель. А на месте везикулы образуется утолщение, которое потом отшнуровывается сначала образуя цистерны, от которых отшнуровываются везикулы: - прохождение ионов кальция через пресинаптическую мембрану обеспечивается ее деполяризацией.
: А — ацетат,")
Синаптические процессы в невозбужденном и возбужденном синапсах (по Л. Щельцыну, 1980): А — ацетат, Х — холин, Хэ — холинэстераза

В целом весь описанный процесс схематически можно представить так Деполяризация пресинаптического волокна Ионы кальция Столкновение везикул с пресинаптической мембраной Выделение ацетилхолина в синаптическую щель

Чем больше деполяризация пресинаптической мембраны, тем больше потенциал на постсинаптической мембране. На постсинаптической мембране имеются белки – холинорецептивные участки. Холинорецепторы постсинаптической мембраны обладают разной структурой и функцией: одни способствуют увеличению выделения ацетилхолина, другие могут затормозить его выделение.

Катионная головка ацетилхолина взаимодействует с анионным участком холинорецептора, что ведет к изменению конформации белковых молекул, встроенных в постсинаптическую мембрану и формирующих ионные каналы. При этом взаимодействии каналы открываются, и изменяется проницаемость для ионов Na+, Ca 2+, K+. Основное значение имеет Na+, его вхождение в постсинаптическую мембрану сопровождается деполяризацией и формированием распространяющегося импульса.

В синапсе постоянно присутствует фермент холинэстераза, который разрушает ацетилхолин. Используя фармакологические вещества можно блокировать или усиливать действие и выделение ацетилхолина. Например: галантолин (получаемый из подснежников) предохраняет ацетилхолин от разрушения. В пресинаптических окончаниях в состоянии покоя непрерывно протекает медленный синтез и медленное выделение ацетилхолина. При возбуждении, одновременно с резким увеличением выделения ацетилхолина возрастает и скорость его синтеза.

В 1950 году Фетт и Катц обнаружили, что даже без раздражения пресинаптического волокна в области концевой пластинки можно обнаружить спонтанно возникающие потенциалы концевой пластинки, в среднем амплитуда их около 0, 5 м. В.
. Средняя частота их возникновения составляет")
Они получили название миниатюрных потенциалов концевой пластинки (м. ПКП). Средняя частота их возникновения составляет около одного в секунду, вызывается м. ПКП спонтанным выделением ацетилхолина в виде квантов. Один квант – это освобождение одной везикулы, в которой содержится от 6 до 10 тыс. молекул ацетилхолина. При высвобождении кванта медиатора возникают только локальные синаптические потенциалы: деполяризационные (ВПСП) или гиперполяризационные (ТПСП). Из-за малой величины они носят название миниатюрных.

В реальной ситуации синхронно высвобождаются несколько сотен квантов медиатора. Суммарный постсинаптический потенциал в этом случае оказывается достаточным для активации потенциал – зависимых каналов. Возникает распространяющийся потенциал действия.
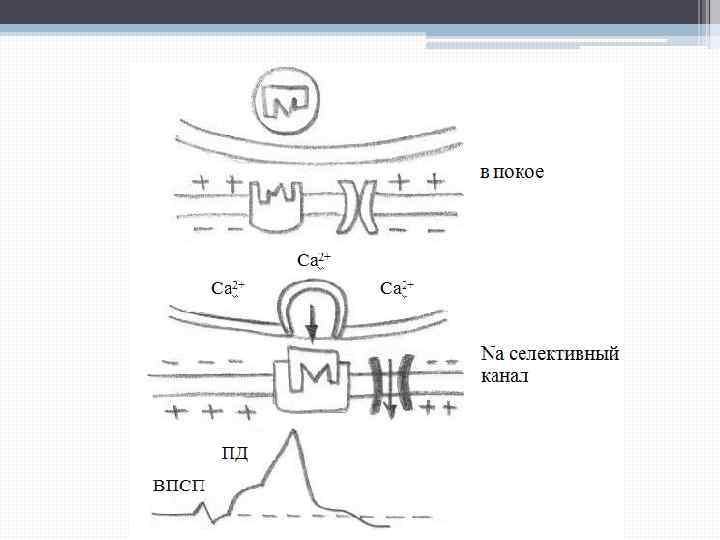

Если рецептор будет сопряжен с К+-каналом, открывается ион-селективный К+-канал, что ведет к развитию тормозного постсинаптического потенциала (ТПСП).

Проведение возбуждения в синапсах центральной нервной системы ВПСП представляет собой основной процесс, обеспечивающий передачу возбуждающих влияний от одной клетки на другую. Такой ВПСП отражает действие не одного, а целого ряда синаптических окончаний. Это обусловлено либо ветвлением аксона одной клетки на многие терминали, каждая из которых дает ряд синаптических окончаний; либо схождение к одной и той же клетке разветвлений многих афферентных волокон.

Во всех без исключения случаях синаптическая передача в центральной нервной системе связана с генерацией в постсинаптической клетке ВПСП. Выделение медиаторов в центральных синапсах происходит таким же квантовым способом, как и в нервно-мышечном синапсе. В большинстве центральных нейронов местом первичной генерации импульсов является начальная часть аксона – это наиболее возбудимая область, лишь после ее возбуждения импульс распространяется в аксон, в сому и в дендриты. Возбуждение в начальном сегменте аксона может возникнуть при деполяризации в 10 м. В, а в соме при деполяризации в 30 м. В.

Возникшее возбуждение, дойдя до клеткиэффектора может вызвать как возбуждение, так и торможение. Один и тот же медиатор, действуя на разные рецепторы постсинаптической мембраны в разных нервных клетках, может вызвать как возбуждение, так и тормозные процессы.

В ЦНС позвоночных животных можно выделить синапсы, которые являются возбуждающими, например: центральные отростки первичных афферентных нейронов оказывают только возбуждающее действие на нейроны спинного мозга. Деполяризация мотонейрона в результате действия возбуждающего медиатора достигается не только за счет увеличения проницаемости его мембраны для ионов натрия (или кальция), но также за счет уменьшения проницаемости для ионов калия.
 показало, что")
Изучение процессов торможения (И. М. Сеченов, 1863; Н. Е. Введенский; Ч. Шеррингтон) показало, что торможение – это самостоятельный нервный процесс, вызываемый возбуждением и проявляющийся в подавлении другого возбуждения. В отличие от возбуждения, которое может иметь форму локального потенциала и потенциала действия, торможение всегда носит форму локального процесса. Торможение всегда связано с существованием специфических тормозных синапсов. Примером тормозных нейронов спинного мозга являются вставочные нейроны Реншоу.
 или")
В ЦНС торможение может развиться вследствие изменения свойств постсинаптической мембраны нейрона (постсинаптическое торможение) или на пресинаптическом уровне (пресинаптическое торможение). Пресинаптическое торможение достигается путем угнетения процесса высвобождения медиатора. Осуществляется это угнетение специальными тормозными вставочными нейронами.

Аксоаксонный синапс, образованный аксоном тормозного вставочного нейрона и аксональным окончанием возбуждающего нейрона. Аксон тормозного нейрона является пресинаптическим, он высвобождает тормозной медиатор (γ-аминомасляную кислоту).

Этот медиатор увеличивает проницаемость мембраны возбуждающего аксона для ионов хлора, что сопровождается уменьшением амплитуды потенциала действия, что в свою очередь уменьшает количество высвобождаемого им медиатора. При этом падает амплитуда ВПСП, трансмембранная разность потенциалов постсинаптической мембраны не достигает критического уровня, потенциал действия не возникает.

Постсинаптическое торможение: медиатор тормозных синапсов, выделяемый пресинаптическимим окончаниями, подавляет способность постсинаптической мембраны генерировать ВПСП или потенциал действия. Изменение в постсинаптической мембране обозначается как ТПСП. Тормозной медиатор повышает проницаемость постсинаптической мембраны для хлорид ионов. Когда концентрация хлорид ионов в клетке превышает его содержание в наружной среде, тормозной медиатор приводит к выходу ионов хлора из клетки наружу, что сопровождается ее деполяризацией. Тормозной эффект тем значительнее, чем ближе к месту генерации потенциала действия расположен тормозной синапс.

Изучение синаптической передачи может дать ключ к пониманию механизма нарушения функций нервной системы. Например, при столбняке наблюдаются неконтролируемые судороги, а при ботулизме – мышечная слабость. А механизм действия обоих токсинов сходен: они повреждают механизм секреции медиаторов. Нейротоксин столбняка является причиной постоянного ("столбнячного") сокращения мышц или спазмов.

Ботокс – препарат американской фирмы Allergan, изготовленный из очищенного нейротоксина типа А, который вырабатывают микроорганизмы Clostridium botulinum. Нейротоксин блокирует передачу двигательного импульса с нерва на мышечное волокно. Расслабляя располагающиеся под кожей мимические мышцы, препарат разглаживает морщины. Ботулиновый нейротоксин также применяется для лечения легких форм нервного тика.

Столбнячный токсин нарушает секрецию тормозных аминокислот в ЦНС, что приводит к беспрепятственному распространению возбуждения. При ботулизме блокируется секреция ацетилхолина в нервно-мышечных синапсах, что делает невозможным управление мышцами. Иногда при синаптической патологии тормозных дофаминергических нейронов возникает мышечный тремор (болезнь Паркинсона), делающий невозможным осуществление точных произвольных движений.

Проведение возбуждения в центральной нервной системе определяется физиологическими свойствами синапсов: Так как проведение волны возбуждения от одного нейрона к другому через синапс проходит с помощью медиатора, а последний содержится лишь в пресинаптической части, то возможно только одностороннее проведение нервных импульсов через синапс – от пресинаптической мембраны к постсинаптической. В связи с этим поток импульсов в рефлекторной дуге имеет определенное направление: от афферентных нейронов к вставочным, а затем к эфферентным мотонейронам и вегетативным нейронам.

Большое значение в деятельности ЦНС имеет замедленное проведение возбуждения через синапс. Это связано с затратой времени на процессы, происходящие от момента прихода пресинаптического импульса до появления изменений поляризации постсинаптической мембраны. Этот интервал называется синаптической задержкой и составляет в мотонейронах примерно 0, 3 мс. В вегетативной нервной системе длительность синаптической задержки более 10 мс.

Синаптическая задержка обусловлена освобождением медиатора из везикул, диффузией медиатора через синаптическую щель, увеличением под его влиянием ионной проницаемости постсинаптической мембраны. После этого еще требуется 1, 2 мс на развитие ВПСП до максимума и возникновение потенциала действия. При утомлении, охлаждении и др. воздействиях длительность синаптической задержки увеличивается. И так как некоторые реакции организма осуществляются с участием большого числа нейронов (т. е. синаптических связей), то суммарная задержка проведения возбуждения по нервным центрам составляет доли секунды или даже целые секунды.

Если раздражение одиночно, то в пресинаптической части синапса освобождается небольшое количество медиатора. При этом в постсинаптической мембране нейрона возникает ВПСП подпороговой величины. Чтобы ВПСП достиг величины порога, требуется суммация подпороговых ВПСП на постсинаптической мембране. Суммация может быть пространственной в случае одновременного поступления нескольких импульсов к одному и тому же нейрону по разным пресинаптическим волокнам.

Обычно требуется одновременная активация 50 – 100 афферентных волокон для получения ответов мотонейрона спинного мозга. Последовательная суммация наблюдается при активации одного и того же афферентного пути серией последовательных импульсов. Если интервалы между импульсами достаточно коротки и ВПСП от предыдущего импульса не успевает затухать, то последующие ВПСП накладываются на них и деполяризация мембраны достигает критического уровня и возникает потенциал действия.

Нервные клетки обладают свойством изменять частоту передающихся импульсов, т. е. свойством трансформации ритма. Так при низком уровне возбудимости и лабильности нейронов происходит урежение ритма. Это обусловлено тем, что несколько приходящих импульсов суммируются, чтобы достичь порога для возникновения потенциала действия. При высокой возбудимости нейрона импульсация может учащаться, возникает мультипликация ритма. Эти изменения ритма могут усиливать или ослаблять ответные реакции организма на внешние раздражения.

Схема выброса медиатора и процессов, происходящих в гипотетическом центральном синапсе Синаптический Медиатор взаимодействует с пузырек высвобождает рецептором. медиатор Канал открывается Перемещение ионов Na+ и К+

Пресинаптическое окончание поглощает медиатор обратно Антагонист блокирует рецептор
Синапсы и их физиологические свойства.pptx