Kopia_El_Genez_2-2_Potentsial_PP_i_PD.ppt
- Количество слайдов: 133


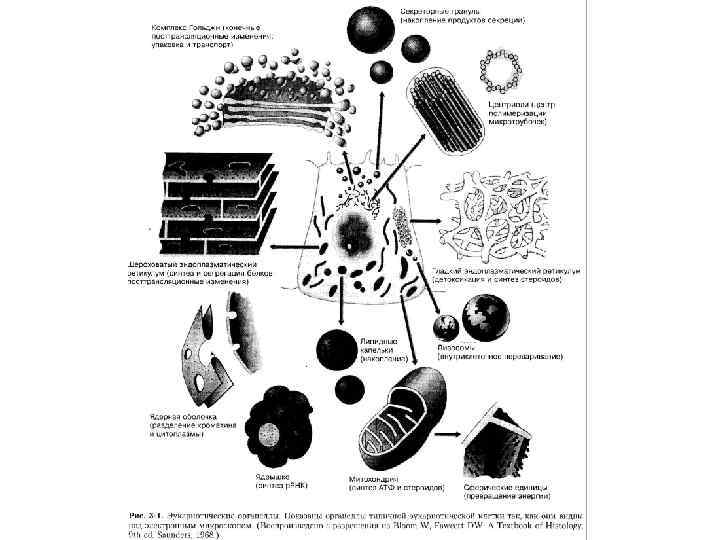
 • 2. Билипидный слой (Гортер и")
Модели мембран: • 1. Липидный слой (Овертон, 1902) • 2. Билипидный слой (Гортер и Грендел, 1925) • 3. «Бутербродная» модель (Даниэли и Девсон, 1935)

В 1964 г. Дж. Робертсон предложил трехслойную модель, добавив к наружному белковому слою мембраны - молекулы гликопротеидов.

1966 г. Дж. Ленард и С. Сингер – предложили жидкомозаичную модель. Согласно этой модели белки «плавают» на поверхности липидного слоя в виде отдельных глобулярных молекул или частиц. G. Vanderkooi, D. Green, 1970 г. Белково-кристаллическая модель. Отличается от жидкокристаллической лишь постулированием существования в мембране жесткой белковой структуры, возникающей в результате дальнодействующих белок-белковых связей.
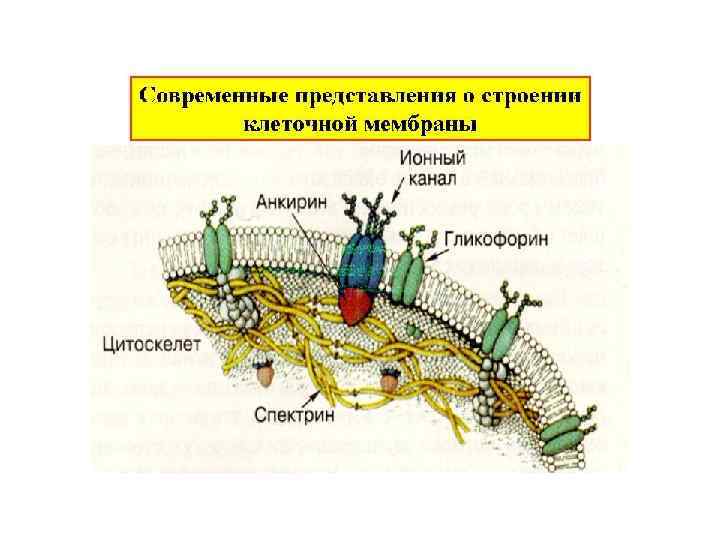

Цитоскелет клетки
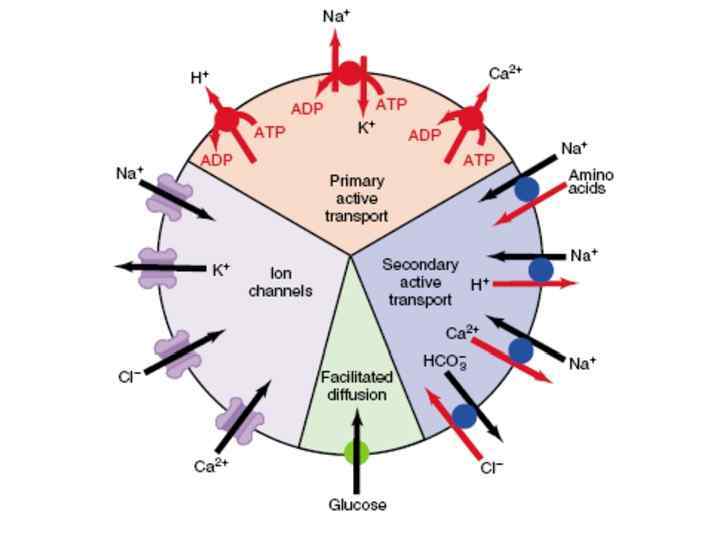
 транспорт:")
• Виды мембранного транспорта: • Прямой • • • Опосредованный Прямой (пассивный) транспорт: 1. простая диффузя; 2. фильтрация; 3. осмос; 4. электроосмос.

Пассивный транспорт Идет без затраты энергии АТФ 1. 2. 3. Движущие силы: Градиент концентрации вещества (химический градиент) Градиент концентрации заряженных частиц (электрохимический градиент) Гидростатическое давление

• Диффузия – самопроизвольный процесс проникновения вещества из области большей концентрации в область меньшей его концентрации, в результате теплового хаотического движения молекул. • Математически этот процесс описывается формулой Фика: • dm/dt = -D·S·dс/dx • dm/dt – скорость диффузии; • • • D – коэффициент диффузии, зависит от природы и молекулярной массы вещества и растворителя, от температуры, свойств мембраны и ее функционального состояния. S – площадь сечения через которую осуществляется диффузия. dс/dx – градиент концентрации, т. е. изменение концентрации вещества с расстоянием.

Фильтрация Это движение растворенных веществ под действием гидростатического давления

• Осмос — поток воды через полупроницаемую мембрану из компартмента с меньшей концентрацией растворённых в воде веществ в компартмент с большей их концентрацией. • Поток воды через биологические мембраны (осмос) определяет разность осмотического и гидростатического давлений по обе стороны мембраны. • Осмотическое давление определяют как давление раствора, прекращающее его разведение водой через полупроницаемую мембрану. Численно осмотическое давление при равновесном состоянии (вода перестала проникать через полупроницаемую мембрану) равно гидростатическому давлению.
 С участием")
Опосредованный транспорт Активный транспорт Без изменения структуры Облегченная диффузия (без затраты энергии) С участием каналов С участием переносчиков унипорт Потен/чувств. Хемо/чувств. котранспорт антипорт АТФ-азы: 3 Nа+ /2 К+ ; 2 H +/Са 2+ С изменением структуры эндоцитоз симпорт экзоцитоз

• Ионные каналы – интегральные белки мембраны, выполняющие функции транспорта для соответствующих ионов. • Они различаются по избирательной пропускной способности к соответствующим ионам (селективности) и типу активации - электрическим или химическим стимулом, соотвественно разделяясь на потенциало- и хемочувствительные. • Селективность канала определяется специфическими свойствами его устья – селективнстью фильтра и знаком заряда , а также геометрией канала. По селективности каналы делятся на: натриевые, кальциевые и хлорные. • •

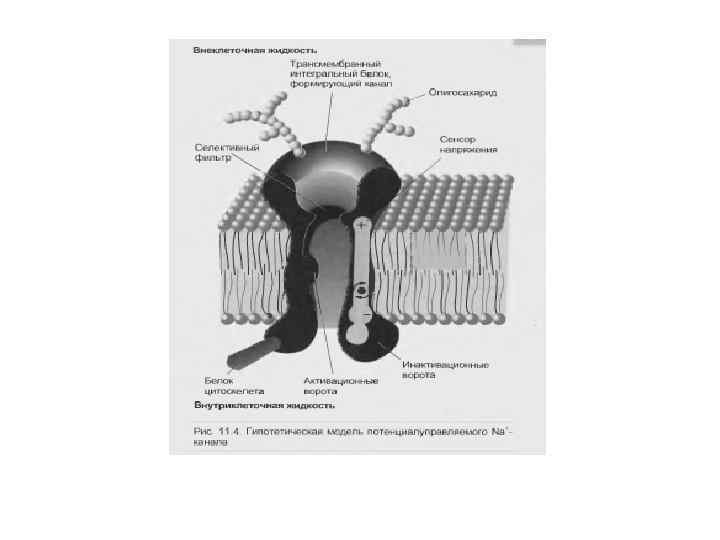
Строение потенциалозависимого ионного канала: 1 — липидный бислой, 2 — сенсор напряжения, 3— ворота, 4 — белковая макромолекула, 5 — якорный белок, 6—углеводные цепи, 7 — селективный фильтр, 8 — водная пора, Р — участок фосфорилирования канала, А — наружный раствор, Б — цитоплазма.

Основы электрофизиологии Общая характеристика возбудимых тканей. Потенциал покоя. Потенциал действия.

Основные проявления жизнедеятельности § § Физиологический покой Физиологическая активность Раздражение Возбуждение Торможение

• • • Общая характеристика возбудимых тканей. Все клетки нашего организма обладают свойством раздражимости. Раздражимость – это способность клеток переходить из состояния физиологического покоя в состояние функциональной активности при действии внешних или внутренних стимулов. • Три вида тканей: нервная, мышечная и секреторная обладают особой формой раздражимости – возбудимостью. • Возбудимость – способность ткани реагировать возбуждением на внешние стимулы. • Возбуждение – реакция на раздражение путем изменения мембранного потенциала.

Раздражение и виды раздражителей. • Раздражение – это процесс воздействия на живой объект внешних по отношению к нему факторов. • Раздражители – факторы внешней среды, вызывающие переход биосистемы в активное состояние. • Ø Ø Ø Раздражители разделяют по следующим факторам: природе раздражителей; биологическому значению; количественному признаку.

По природе раздражителя: • 1. физические ( температура, звук, свет, электрический ток и т. д. ). • 2. химические (соли, кислоты, щелочи и т. д. ). • 3. физико-химические (изменение осмоса, парциального давления газов, р. Н). • 4. биологические (вирусы, бактерии, токсины антитела и т. д. ).

• По биологическому значению: • 1. адекватные – раздражитель к которому в естественных условиях у биосистем есть специализированные воспринимающие структуры (рецепторы). • Свет – для фоторецепторов; • Упругие механические колебания среды – для рецепторов слуха. • 2. неадекватные – раздражители не являющиеся в естественных условиях средством возбуждения для данных биоструктур.

• По количественному признаку или порогу раздражения: • В зависимости от силы действия раздражители делятся на: • 1. пороговые – минимальная сила раздражителя, способная вызвать возбуждение. подпороговые – сила раздражения ниже порога возбуждения. • 2. максимальные – минимальная сила раздражителя, вызывающая максимальный ответ ткани. Раздражители, сила которых меньше или больше максимальной называются, соответственно: субмаксимальными и супермаксимальными.

Все возбудимые ткани обладают общими свойствами: • • Возбудимостью Проводимостью Лабильностью Аккомодацией

• Возбудимость количественно характеризуется порогом возбуждения, который определяется минимальной силой раздражения, способной вызывать возбуждение. • Проводимость – способность проводить возбуждение. Оценивается по скорости распространения возбуждения (м/с). • Лабильность – функциональная подвижность ткани. Оценивается по максимальному числу импульсов, которое возбудимая ткань способна воспроизвести в соответствии с частотой раздражения. Нерв – до 1000 П Д/с, мышца – 500 ПД/с, синапс – 50 ПД/с. • Аккомодация – повышение порога возбуждения к медленно нарастающему или постоянно действующему раздражителю.

Законы раздражения: Ø Закон силы Ø Закон времени Ø Закон крутизны нарастания раздражителя

Закон силы Чем сильней раздражение, тем до определенного предела сильнее ответная реакция биосистемы. Закон постулирован для целостного биологического объекта. Например, нервного ствола, состоящего из отдельных аксонов; скелетной мышцы и т. д. Связано это с тем, что каждое отдельное волокно отвечает на раздражитель по типу «все или ничего» , но порог возбуждения у них отличается. Поэтому суммарная активность объекта будет находится в градуальной зависимости от силы раздражителя.

Закон времени • Чем длительнее раздражение, тем сильней до определенных пределов ответная реакция живой ткани. • В определенном диапозоне эта зависимость имеет характер гиперболы. На ней выделяют реобазу, полезное время и хроноксию. • Реобаза - соответствует порогу возбуждения. • Полезное время – минимальное время, в течение которого раздражитель, равный одной реабазе, должен действовать на ткань, чтобы вызвать возбуждение. • Хроноксия – время, в течение которого должен действовать раздражитель, равный двум реабазам, чтобы вызвать возбуждение.

ЗАКОН «СИЛА - ДЛИТЕЛЬНОСТЬ» Этот закон был экспериментально установллен и сформулирован независимо друг от друга тремя учеными: Лапик, Гоорвиг, Вейс.

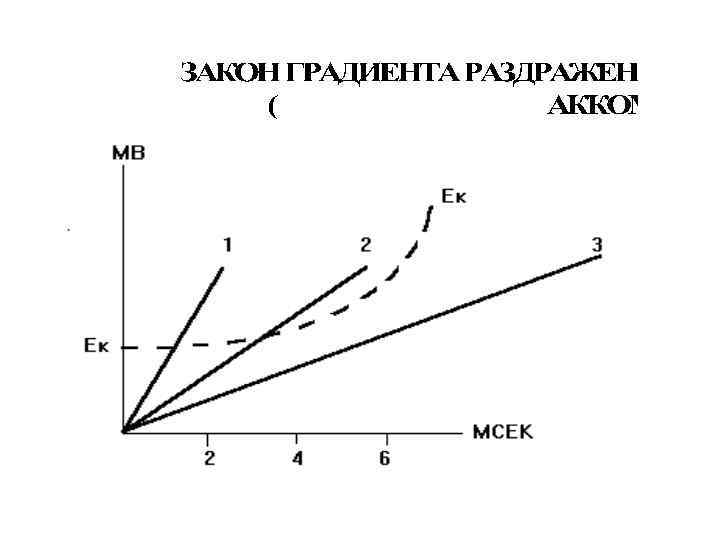
Закон крутизны нарастания раздражителя или градиента: • Чем выше крутизна нарастания раздражителя во времени, тем больше до известного предела величина функционального ответа. • В основе этого закона лежат физико-химические и функциональные изменения, вызываемые в раздражаемом объекте. • Возбуждение развивается в том случае, если активационные процессы достигают пороговой критической величины раньше инактивационных.

История открытия биоэлектри чества.


Первый опыт Гальвани При подвешивании нервно - мышечного препарата с помощью медного крючка, проходящего через спинной мозг препарата, имело место сокращение мышц лапки каждый раз, когда эта лапка соприкасалась с железным поручнем балкона.

ВОЛЬТОВ СТОЛБ Однако ему возразил его знаменитый соотечественник - физик А. Вольта, который считал, что в «балконном» опыте мышца является лишь чувствительным «электрометром» электричества, порождаемого контактной разностью потенциалов разных металлов, используемых в опытах Гальвани. Вольтов столб, состоящий из металлических дисков, разделенных кружками мокрой ткани.

Второй опыт Гальвани Позднее, защищая свою точку зрения от возражений оппонентов, Гальвани воспроизводит различные модификации опытов, в которых сокращение мышцы вызывалось путем набрасывания нерва с помощью стеклянной палочки на поврежденный и неповрежденный участок мышцы
. • Однако, Маттеучи обнаружил явление вторичного")
Открытия Гальвани были подтверждены в работах Маттеучи (1837). • Однако, Маттеучи обнаружил явление вторичного или индуцированного сокращения: при помещении нерва одного нервно-мышечного препарата на мышцу другого препарата и раздражителя нерва этого препарата, Маттеучи наблюдал сокращение мышцы обоих препаратов. •

Опыт Маттеучи
 В этих работах благодаря")
Работы Э. Дюбуа-Реймона ( выполненны в 1840 -1860 гг. ) В этих работах благодаря высокочувствительному гальванометру и ряду других технических новшеств удалось впервые определить электрические процессы в мышце, зарегистрировав потенциал наружной и внутренней поверхности мембраны клеток. • Он впервые установил, что наружная мембрана заряжена положительно по отношению к внутренней, и эта разность потенциалов изменяется при сокращении мышцы. •
 мв +30 0 -75 Ео")
Природа и происхождение потенциал покоя (Ео) мв +30 0 -75 Ео

• В 1896 г. В. Ю. Чаговец впервые высказал гипотезу о ионном механизме электрических потенциалов в живых клетках. Он сделал попытку применить теорию электролитической диссоциации Аррениуса - для их объяснения. • В 1902 г. Бернштейном была развита мембранно-ионная теория, согласно которой клеточная поверхность представляет собой полупроницаемую мембрану, которая в состоянии физиологического покоя проницаема для ионов калия и практически непроницаема для остальных вне - и внутриклеточных ионов.

Диффузионный концентрационный потенциал
![Уравнение НЕРНСТА Ек = RT n. F ℓn [ K +o ] [ K](https://present5.com/presentation/3/23632904_132457025.pdf-img/23632904_132457025.pdf-42.jpg "Уравнение НЕРНСТА Ек = RT n. F ℓn [ K +o ] [ K")
Уравнение НЕРНСТА Ек = RT n. F ℓn [ K +o ] [ K +i ] Ек – равновесный калиевый потенциал; R - универсальная газовая постоянная; T - температура среды; n - валентность иона ( для К+ = 1); F - число Фарадея; К+ - концентрация калия с внешней (о) и внутренней (i) стороны мембраны.

В 1936 году английский зоолог Джон Юнг обнаружил у кальмаров и каракатиц необычайно толстые аксоны, которые впоследствии стали называть "гигантскими аксонами". Их диаметр превышал 0, 5 мм, что позволило достаточно легко вводить в них микроэлектроды, проводить химический анализ содержащейся в них жидкости, вводить в них различные растворы и т. д. «Гигантские аксоны» стали излюбленным объектом для изучения биоэлектрических явлений в тканях, с их помощью было получено много новых и интересных данных.

Экспериментальная проверка гипотезы Бернштейна n В 1962 г. Ходжкин с сотрудниками подтвердил гипотезу Бернштейна: n Из аксона кальмара выдавили аксоплазму и спавшуюся оболочку заполнили искусственным солевым раствором. Когда концентрация калия была близка к внутриклеточной, - между внутренней и наружной мембраной установилась разность потенциалов близкая к ПП (-50/-80 м. В), и волокно проводило импульсы. n При снижении концентрации калия или увеличении – мембранный потенциал менялся, или даже изменял свой знак (потенциал становился положительным, если в наружном растворе [ K +o] > [ K +i] ). n Это доказывало, что концентрационный градиент калия является основным фактором, определяющим величину ПП нервного волокна.

Современные представления о природе биоэлектрических явлений в тканях базируются на результатах работ Алана Ходжкина, Эндрью Хаксли, Бернарда Катца. Эти ученые в 40 -50 годах нашего века модифицировали и экспериментально обосновали мембранно-ионную теорию Ю. Бернштейна. В настоящее время их взгляды о природе биоэлектрических явлений пользуются всеобщим признанием. Согласно их представлениям, наличие электрических потенциалов в живых клетках обусловлено различной концентрацией ионов Na+, K+, Ca 2+ и Cl- внутри и вне клетки, а также различной проницаемостью для них клеточной мембраны.

 Все клетки живого организма поляризованы. Между внутренней средой")
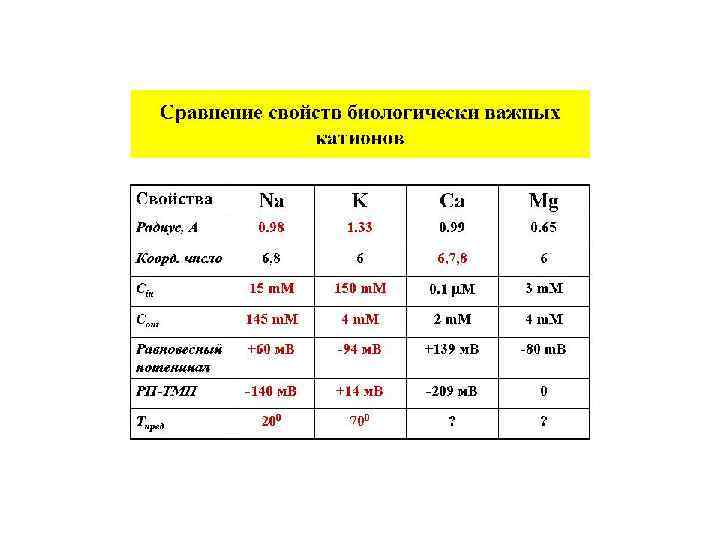
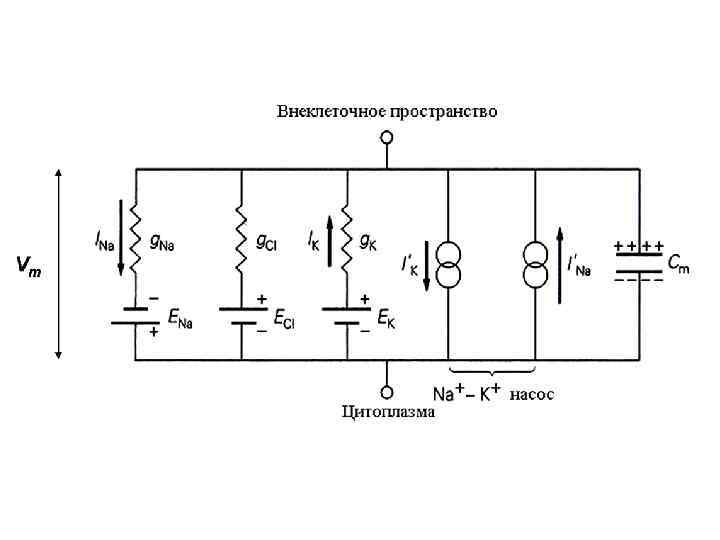
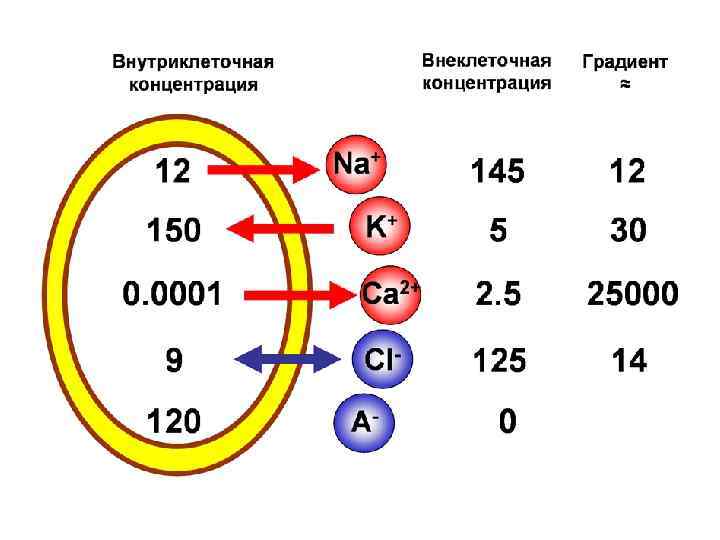
Мембранно-ионная теория происхождения потенциала покоя (ПП) Все клетки живого организма поляризованы. Между внутренней средой клетки и внешней средой существует разность потенциалов мембранный потенциал (потенциал покоя). Существование животного электричества открыл Гальвани в 1791 г. Авторы современной теории: Ходжкин, Катц, Хаксли 1949 -1952 гг. Положения их теории: Ионы вне и внутри клетки распределены неравномерно. Катионов калия внутри клетки больше в 40 -50 раз, натрия меньше в 8 -12 раз, анионов хлора меньше в 30 -40 раз. Разность концентраций ионов создается работой ионных насосов Проницаемость мембраны для разных ионов различна. В состоянии покоя проницаемость для калия, натрия и хлора неодинаковы: Р калия : Р натрия : Р хлора = 1 : 0, 04 : 0, 45
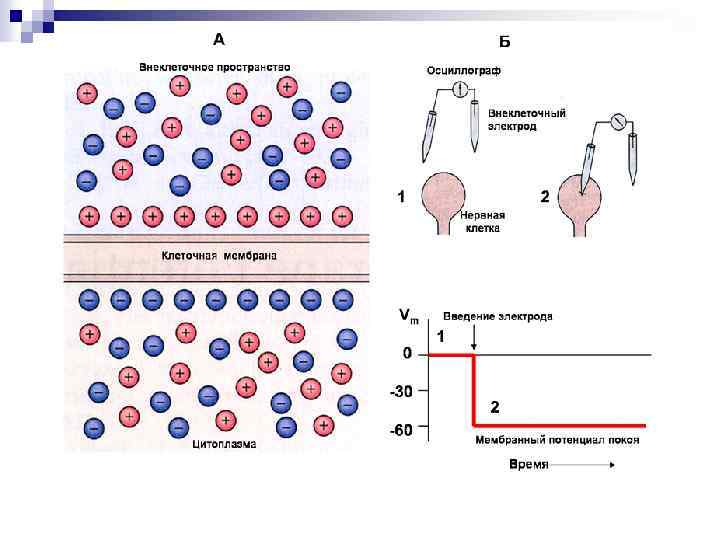

Концентрации Na+, K+ и Cl– внутри и вне клетки и пути проникновения этих ионов через клеточную мембрану : через специфические ионные каналы или с помощью Na/K–насоса. При данных концентрационных градиентах равновесные потенциалы ENa, EK и EСl– равны указанным, мембранный потенциал Em = – 90 м. В

Механизм поляризации мембраны в покое Наружная часть клетки К+ =10 Мэкв/л Na+ 350 Мэкв/л JNa = +30 m. V Клеточная мембрана + + + + К-Na -насос + + - Внутренняя часть клетки К+ =500 Мэкв/л JK= -90 m. V K+ Na+ 35 Мэкв/л
![Уравнение ГОЛЬДМАНА Ем RT = n. F ℓn Pk [ Ko+] + PNa [](https://present5.com/presentation/3/23632904_132457025.pdf-img/23632904_132457025.pdf-52.jpg "Уравнение ГОЛЬДМАНА Ем RT = n. F ℓn Pk [ Ko+] + PNa [")
Уравнение ГОЛЬДМАНА Ем RT = n. F ℓn Pk [ Ko+] + PNa [ Nao+]+PCl [ Cli- ] Pk[ Ki+]+ PNa [ Nai+] + PCl[ Clo–] Ем – равновесный калиевый потенциал; R - универсальная газовая постоянная; T - температура среды; n - валентность иона; F - число Фарадея; P – проницаемость иона для мембраны; К+ - концентрация калия с внешней (о) и внутренней (i) стороны мембраны. Na+ - концентрация натрия с внешней (о) и внутренней (i) стороны мембраны. Cl- - концентрация хлора с внешней (о) и внутренней (i) стороны мембраны.

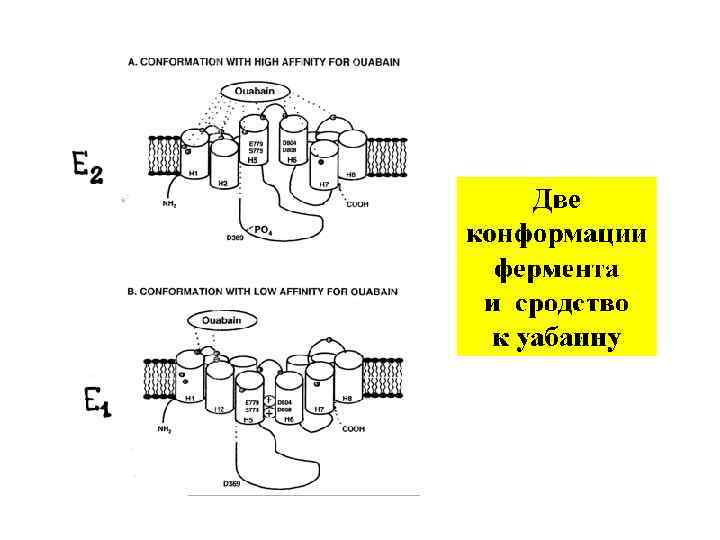
УАБАИН - блокатор Nа/К- АТФ-азы

Физическая модель мембраны

Потенциал покоя создается: 1. Неравновесным распределением калия, натрия, хлора и кальция внутри клетки по отношению к внешней среде. 2. Разной проницаемостью мембраны для этих ионов. 3. Основным потенциалобразующим ионом является катион калия. 4. Постоянство потенциала покоя поддерживается работой ионных насосов. 5. Электрогенность калий-натриевого насоса вносит дополнительный вклад в создание мембранного потенциала.

Потенциал действия
 ПД – это быстрое изменение мембранного потенциала, которые")
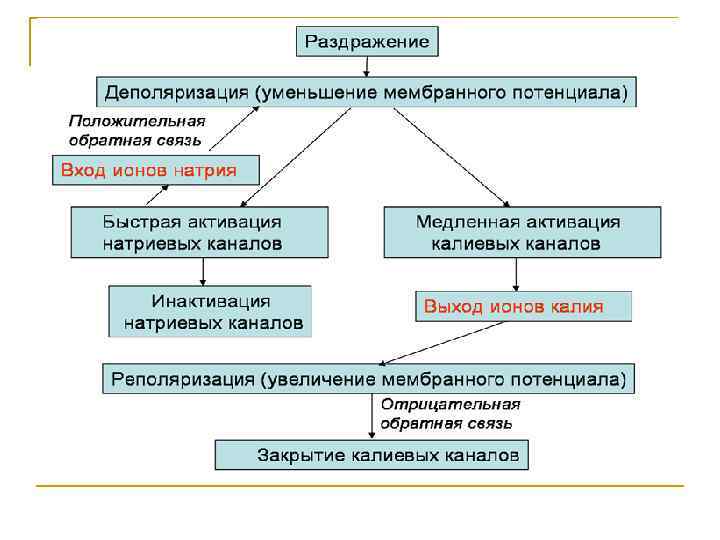
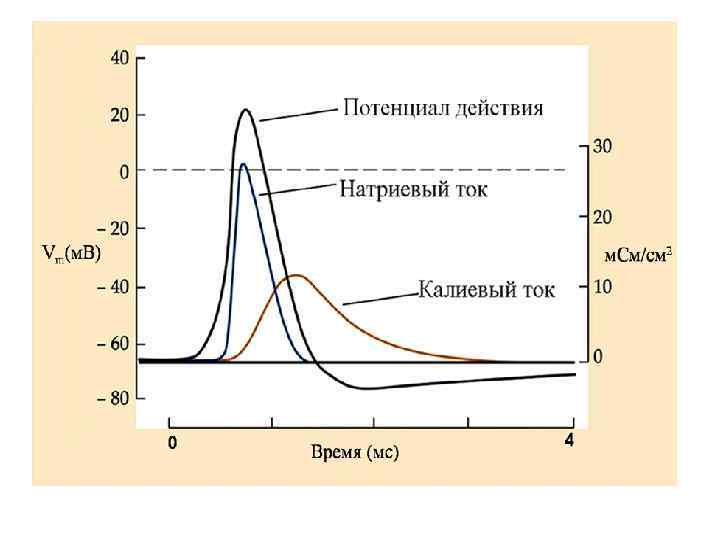
Мембранно-ионная теория происхождения потенциала действия (ПД) ПД – это быстрое изменение мембранного потенциала, которые возникают в месте нанесения раздражения и распространяются на всю мембрану клетки. В основе возбуждения лежит изменение проницаемости мембраны для натрия и калия, связанное с активацией потенциалзависимых натриевых и калиевых каналов.
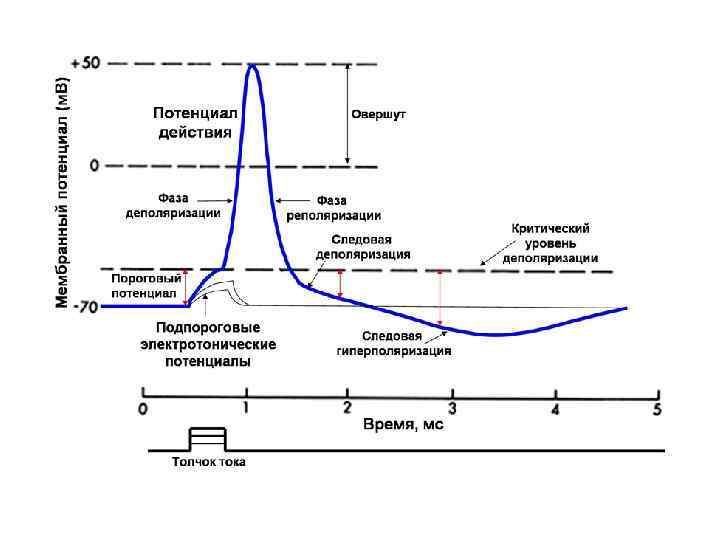

При действии на клетку раздражителя проницаемость мембраны для Ø Ø ионов Na+ резко повышается за счет активации (открывания) натриевых каналов. При этом ионы Na+ по концентрационному градиенту интенсивно перемещаются из вне - во внутриклеточное пространство. Вхождению ионов Na+ в клетку способствует и электростатическое взаимодействие. В итоге проницаемость мембраны для Na+ становится в 20 раз больше проницаемости для ионов К+. Поскольку поток Na+ в клетку начинает превышать калиевый ток из клетки, то происходит постепенное снижение потенциала покоя, приводящее к реверсии - изменению знака мембранного потенциала. При этом внутренняя поверхность мембраны становится положительной по отношению к ее внешней поверхности. Указанные изменения мембранного потенциала соответствуют восходящей фазе потенциала действия - фазе деполяризации.

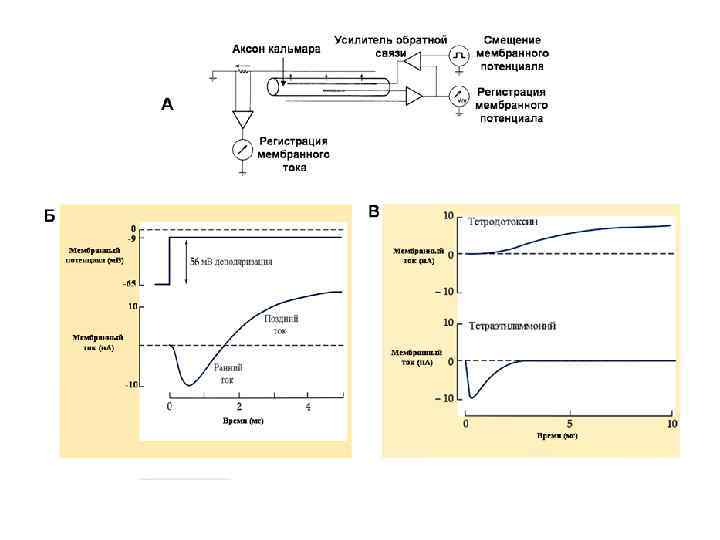
Экспериментальное доказательство натриевой природы деполяризации В 1939 г. Ходжкин и Хаксли установили, что мембранный потенциал во время развития ПД не просто уменьшается до нуля, а меняет свой знак на противоположный. Эти данные противоречили гипотезе Бернштейна о том, что во время ПД мембрана становиться проницаемой для всех ионов. Важным аргументом в пользу натриевой природы деполяризации ПД стал факт тесной зависимости амплитуды ПД от [ Nao+]. В опытах с перфузией аксонов кальмара изнутри солевыми растворами установлено: При замене аксоплазмы солевым раствором, в котором К+ частично заменен на Na+, - амплитуда ПД резко снижается. Ø При полной замене К+ на Na+ волокно полностью утрачивает способность генерировать ПД. Ø При замене внеклеточного Na+ на холин хлорид нервное волокно также полностью утрачивает способность генерировать ПД. Ø Все это позволило сделать вывод о зависимости величины ПД от градиента натрия.

n n Мембрана характеризуется повышенной проницаемостью для ионов Na+ лишь очень короткое время 0. 2 - 0. 5 мс. После этого проницаемость мембраны для ионов Na+ вновь понижается, а для К+ возрастает. В результате поток Na+ внутрь клетки резко ослабляется, а ток К+ из клетки усиливается. В течение потенциала действия в клетку поступает значительное количество Na+, а ионы К+ покидают клетку. Восстановление клеточного ионного баланса осуществляется благодаря работе Na+/К+-АТФ-азы. Итогом этих процессов и является реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к внешней поверхности мембраны.

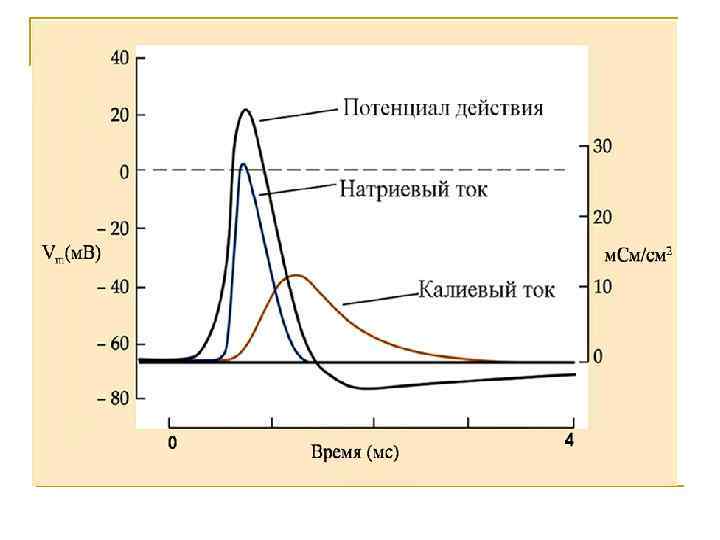
Мембранные проводимости во время потенциала действия в гигантском аксоне кальмара, g. Na и g. К рассчитывали, подавая серии деполяризующих скачков потенциала

ЦИКЛ ХОДЖКИНА-ХАКСЛИ Раздражитель деполяризация мембраны возрастание входящего Na+ - тока повышение Na+ проницаемости

m m h h Р Е А К Т И В А Ц И Я Na-канала

Потенциалзависимая инактивация Na+ – системы. По оси абсцисс отложены величины отклонения мембранного потенциала от потенциала покоя (— 60 м. В). От каждого из этих исходных значений потенциала мембрану деполяризовали до — 16 м. В и по оси ординат откладывали отношения возникающих максимальных Na+–токов (INa+ max) к величине INa+ max’, соответствующий полной +–системы. активации Na

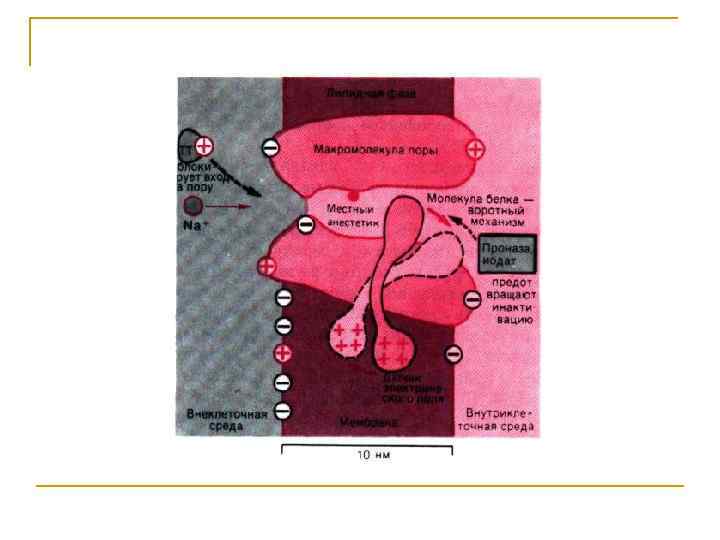
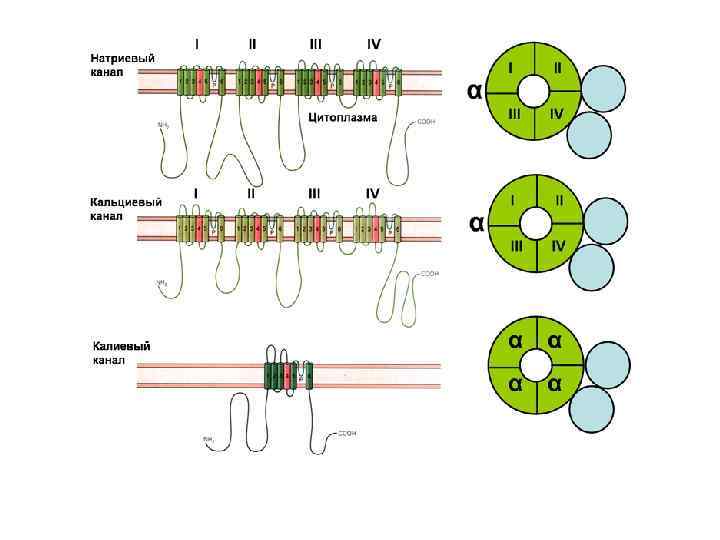
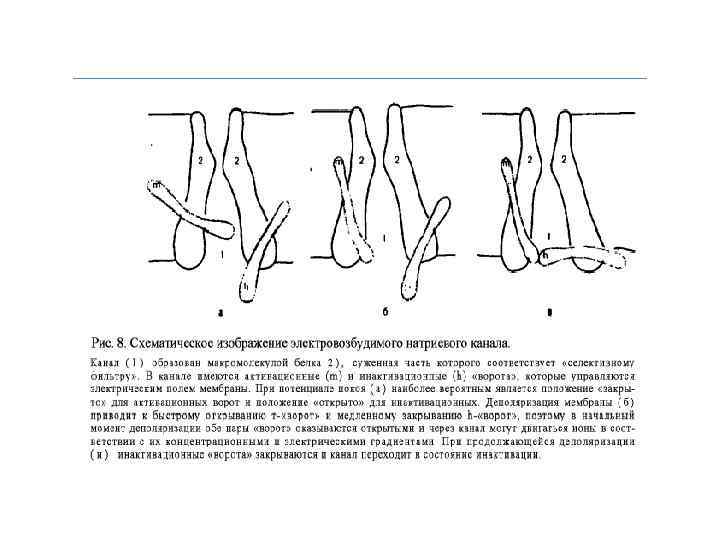
Исследование строения Na+-канала Узкая часть Na+-поры, служащая фильтром для ионов, - несет одиночный отрицательный заряд (СООН -группа). Этот заряд притягивает катионы и отталкивает анионы. Ион сопровождается молекулами воды, образующими водородные связи с порой. В поре имеется достаточно места для того, чтобы натрий мог войти в сопровождении 3 -х молекул воды и в тоже время образовать тесный контакт с 3 -я молекулами кислорода поры. В этом смысле Na+канал является «водной порой» , хотя молекулы воды в ней не могут также свободно двигаться, как в массе воды. Заряженные группировки белков Na+-канала реагируют на деполяризующее действие электрического тока конформационными перестройками определенных участков белковой молекулы. Именно они и лежат в основе воротных процессов, управляющих проводящим состоянием канала.
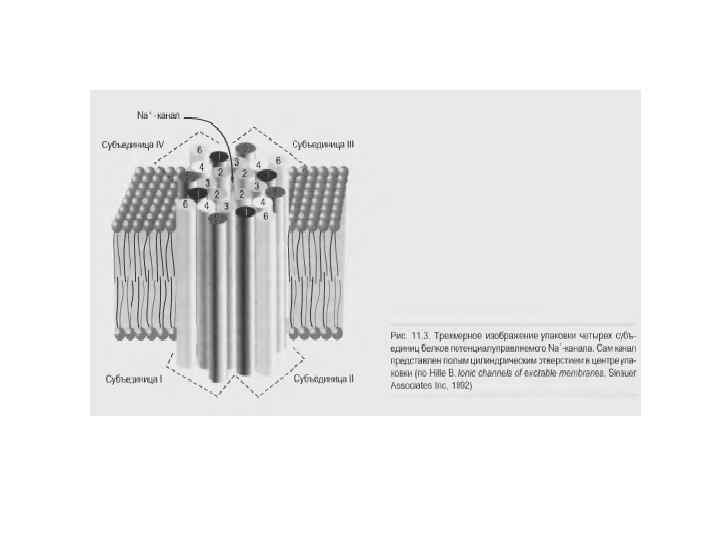

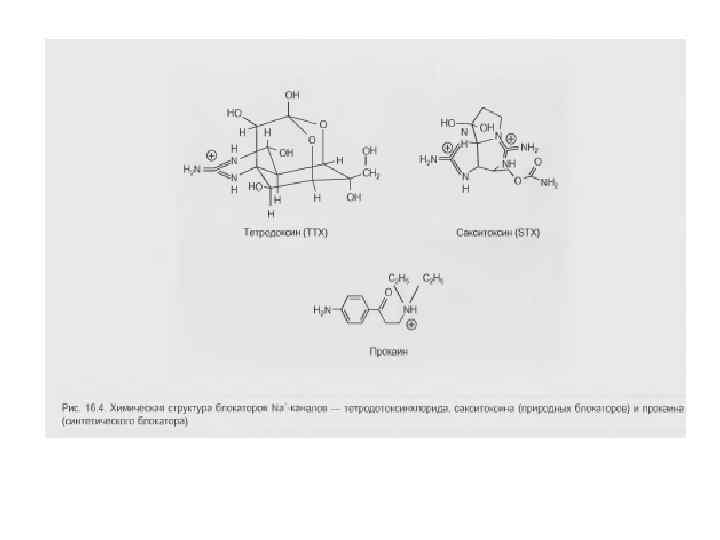
Инактивация Na+-канала развивается автоматически и степень ее зависит от мембранного потенциала и времени. Инактивацию можно подавить введя в цитоплазму протеолитический фермент (проназу). Это предполагает, что в инактивации участвует белковая структура, расположенная у внутреннего входа Na+-канала. Кинетика блокирования ТТХ Na+-каналов показала, что на участке мембраны площадью в 1 мкм 2 располагается ~ 100 Na+-каналов. Если считать, что d канала ~ 0, 5 нм, - общая площадь их сечения составляет менее 1/50 000 от поверхности мембраны. Активированный Na+-канал пропускает примерно 6000 ионов/мс. Время нахождения канала в открытом состоянии – менее 1 мс и зависит от мембранного потенциала: при смещении деполяризующего потенциала в положительную сторону это время уменьшается.

Nа+ - потенциалозависимые каналы, в мембране нервного и скелетного мышечного волокна, характеризуются, как время - так и потенциалозависимостью. Активируются на уровне ~ -60 м. В, полностью инактивируются уже при деполяризации мембраны до ~ -40 м. В. Их работа подчиняется закону цикла Ходжкина - Хаксли и обеспечивает кратковременный, но мощный быстрый входящий Nа- ток.

m m m Блокада калиевых каналов тетраэтиламмонием резко удлиняет процесс реполяризации

Экспериментальные исследования структуры К+канала Для проверки гипотезы о том, что выходящий ток по своей природе является калиевым, Ходжкин и Хаксли в 1953 г. , используя радиоактивный калий, изучили перемещение К+ через мембрану под действием постоянного тока. Оказалось, что деполяризующий ток приводил к большому выходу калия (К 42). Диаметр К+-канала меньше натриевого и составляет 0, 3 нм. Ион калия проходит через такой канал без гидратной оболочки, а ион Nа пройти не может, хотя и меньше по диаметру, из-за стерического несоответствия. Установлено, что проводимость одиночного К+ -канала в 3 раза выше, чем у натриевого. Однако, плотность калиевых каналов на единицу площади мембраны в 5 раз меньше.

КАЛИЕВЫЙ КАНАЛ Схема белка, образующего калиевый канал, погруженный в липидный бислой плазматической мембраны. Четыре отрицательных заряда зафиксированы на «стенке» канала. Б. Схематический энергетический профиль канала, показанного на рис. А. По оси ординат отложены величины кинетической энергии, необходимые для прохождения канала; по оси абсцисс –расстояния между внутренней и наружной поверхностями мембраны. Энергетические минимумы соответствуют местам связывания положительно заряженных ионов с фиксированными отрицательными зарядами в стенке канала. Энергетические максимумы соответствуют препятствиям диффузии в канале. Предполагается, что конформация канального белка спонтанно осциллирует; варианты энергетических профилей изображены сплошными и штриховыми линиями; эти осцилляции в значительной степени облегчают связывание ионов при преодолении энергетического барьера.

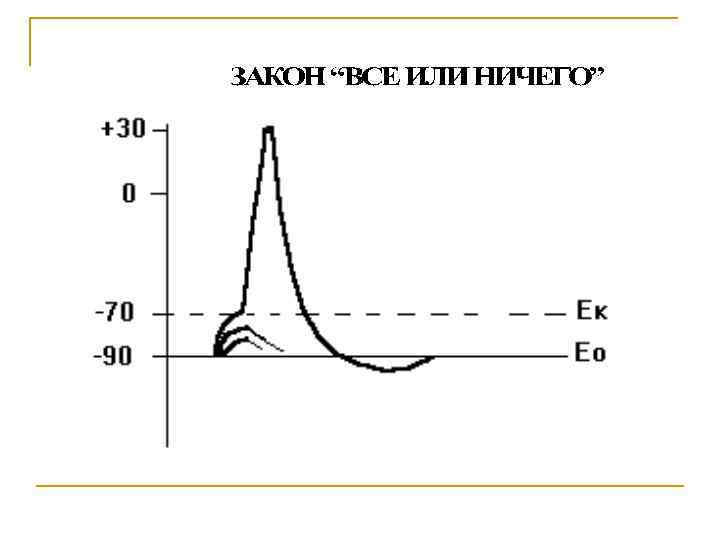
Потенциал действия: 1. Возникает в ответ на пороговые и сверхпороговые раздражители. 2. Обусловлен опережающим увеличением проницаемости мембраны для натрия по сравнению с калиевой. При этом деполяризация определяется натриевым током, реполяризация - калиевым. 3. Вслед за основными фазами потенциала действия возникают следовые потенциалы. Следовой отрицательный связан с повышенной проводимостью мембраны для натрия, положительный - для калия. 4. Потенциал действия распространяется и подчиняется правилу “Все или ничего”. 5. При возбуждении активируется натрий-калиевый насос.

ТРИ СОСТОЯНИЯ МЕМБРАНЫ
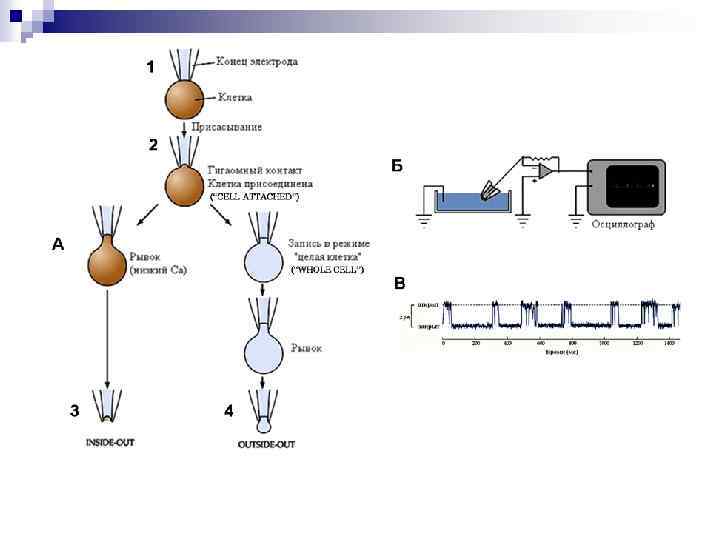
. Изображен продольный срез через регистрирующую")
Схема локальной фиксации мембранного потенциала ( «пэтч–кламп» ). Изображен продольный срез через регистрирующую микропипетку (обозначена черным цветом) с диаметром контактирующего с мембраной кончика ≈1 мкм. Если кончик электрода абсолютно чист и поверхность клетки освобождена от волокон соединительной ткани, то при подаче через пипетку отрицательного давления образуется тесный контакт, который создает электрическую изоляцию каналов находящегося в кончике пипетки микроучастка мембраны от остальной мембраны клетки (вставка). Таким способом можно регистрировать токи каналов с помощью усилителя обратной связи, соединенного с раствором электролита в пипетке.
 и калиевые (справа) каналы (схематическое изображение)")
Токи через натриевые (слева) и калиевые (справа) каналы (схематическое изображение)


ЛОКАЛЬНЫЙ ОТВЕТ. ЗАКОНЫ ДЕЙСТВИЯ ПОСТОЯННОГО ТОКА. Проведение возбуждения по нерву.

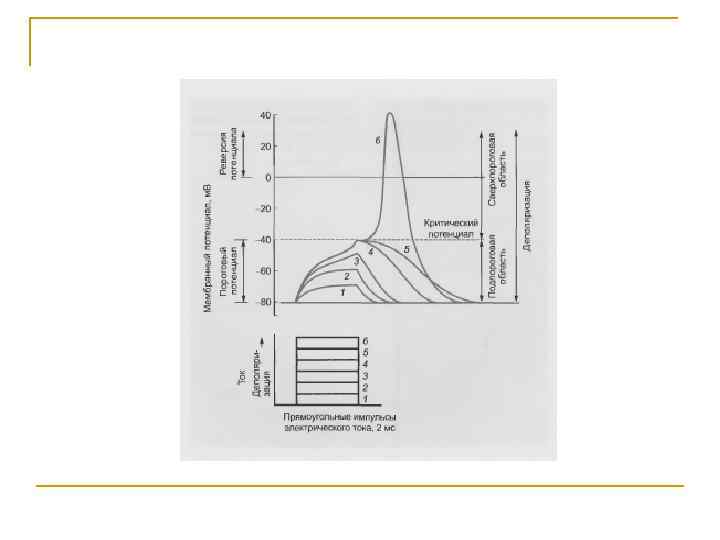
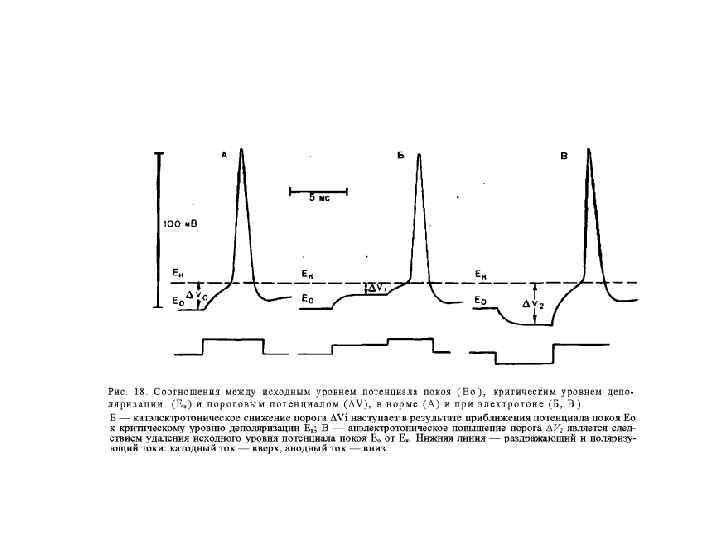
Отличия локального ответа от потенциала действия: Локальный ответ Eк Eo 1. Возникает на подпороговые раздражители. 2. Не распространяется. 3. Не подчиняется правилу “Все или ничего”. 4. Способен к суммации. 5. Возбудимость в период локального ответа повышена. Если локальный ответ достигнет уровня критической деполяризации, он перерастает в потенциал действия.

Основные электрофизиологические феномены в нервном волокне Аэт — анаэлектротон, КУД — критический уровень деполяризации, Кэт — катэлектротон, ЛО — локальный (подпороговый активный) ответ, МПП — мембранный потенциал покоя, ПД — потенциал действия, СП (отр и пол) — следовые потенциалы отрицательный и положительный (временные соотношения пика ПД и СП не выдержаны; отрицательный СП и особенно положительный СП значительно длительнее).

Локальные ответы и закон силы
 -75 м.")
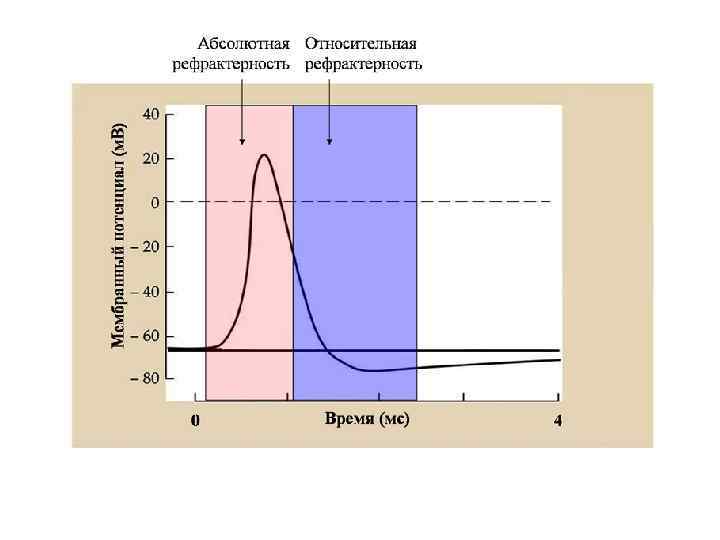
СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИ С ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ 0 Екр( -60 м. В) -75 м. В

Полярный закон Пфлюгера Возбуждение возникает в момент замыкания цепи под катодом, а в момент размыкания цепи под анодом.

Полярный закон действия тока – возбуждение возникает под катодом при замыкании и под анодом при размыкании тока Цепь событий, развивающихся под катодом раздражающего тока: • пассивная деполяризация мембраны • повышение натриевой проницаемости • усиление потока Na++ внутрь волокна • активная деполяризация мембраны • локальный ответ • достижение критического уровня (Ек ) • регенеративная деполяризация • потенциал действия (ПД).

- - - - - - - + + - - - - + + + - - - - КАТОД АНОД «-» «+» Пассивные изменения
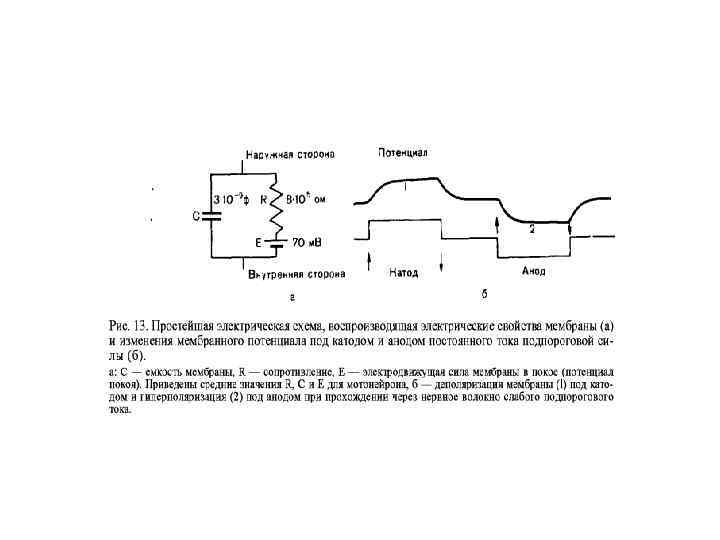

Закон физиологического электротона В момент замыкания цепи возбудимость и проводимость под катодом увеличиваются – катэлектротон; а под анодом – уменьшаются – анэлектротон; При размыкании цепи возбудимость под катодом уменьшается – обратный катэлектротон; а под анодом – увеличивается – обратный анэлектротон.

Физиологический электротон

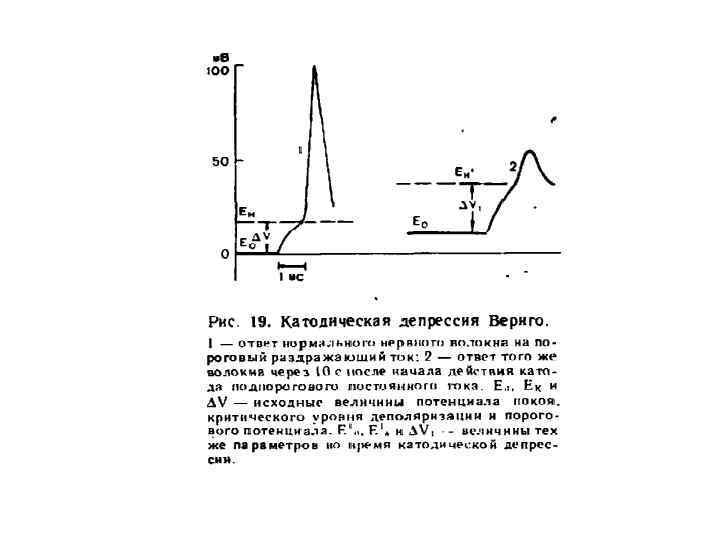
Изменения возбудимости при длительном действии катода Катодическая депрессия Вериго Катодзамыкательное возбуждение Ек-2 Ек-1 Ео замыкание размыкание

Изменения возбудимости при длительном действии анода Ек-1 Ек-2 Ео замыкание размыкание
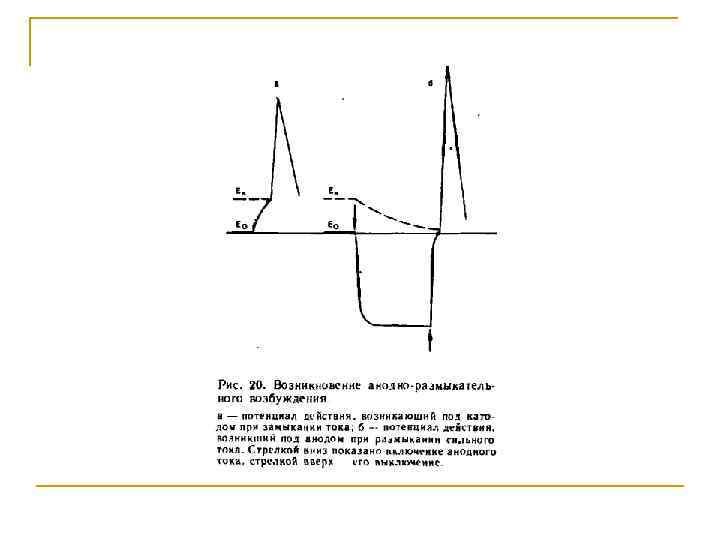

ПОТЕНЦИАЛ ПОКОЯ АНОД ВКЛЮЧЕН АНОД ВЫКЛЮЧЕН Увеличение числа реактивированных Nа+-каналов Выходящий К+-ток значительно превышает входящий Na+-ток Уменьшение количества открытых К+-каналов Входящий Nа+-ток превышает Выходящий К+-ток
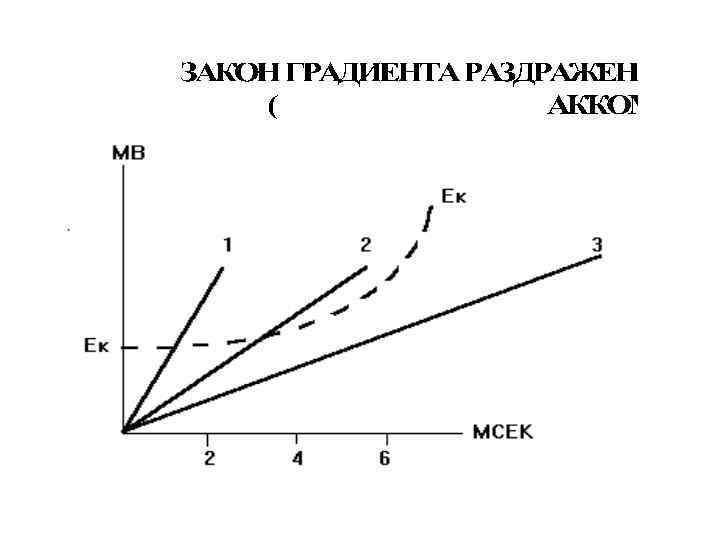

Критический наклон равен отношению реобазы тока с минимальной скоростью нарастания силы раздражителя к реобазе прямоугольного толчка тока КН = Реобаза

Парабиоз Местное нераспростроняющееся и углубляющееся во времени возбуждение. Обнаружен Введенским при исследовании способности нерва проводить высокочастотные разряды импульсов после воздействия на нерв различных химических агентов. В дальнейшем было показано, что состояние парабиоза можно вызвать действием раздражителя любой природы.

Парабиоз и его фазы • Уравнительная • Парадоксальная • Тормозная нерв NH 4
")
сила раздражения (Гц)
 -75 м.")
СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИ С ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ 0 Екр( -60 м. В) -75 м. В

l Введенский впервые обратил внимание на то, что способность ткани воспроизводить задаваемый ритм связана с ее функциональным состоянием – лабильностью. l Явление парабиоза широко распространено в природе – зимняя спячка (анабиоз), как хладнокровных, так и теплокровных (медведи). l Использование наркотических веществ позволяет проводить многочасовые операции, а анестетики – блокируют болевые ощущения. l Открытие парабиоза, а затем пессимального торможения, позволило Введенскому выдвинуть положение о тормозных явлениях в нервах, как о стойком нераспростроняющемся возбуждении.

Образование миелинового волокна

Миелиновое нервное волокно миелин перехваты Ранвье

Скорость передачи сигнала по нервным волокнам
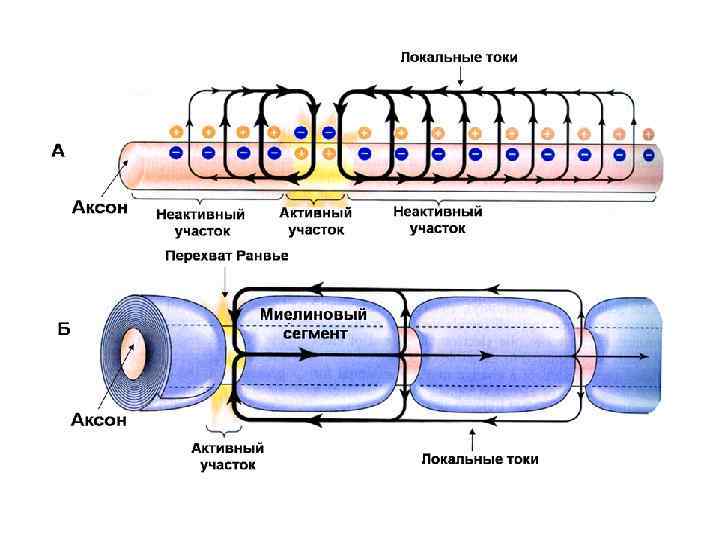

В основе распространения возбуждения - возникновение местных токов между деполяризованным и покоящимся участками. Между различно заряженными участками мембраны возникает локальный ионный ток, который деполяризует мембрану до критического уровня. Причем подпороговый деполяризующий ток идет по аксоплазме, то есть - изнутри. В результате этой подпороговой деполяризации открываются Nа-каналы и возрастает входящий Nа ток. Отношение величины ПД к пороговому току называется фактором надежности: Амплитуда ПД, м. В Величина Екр, м. В Плотность Nа+-каналов в перехватах Ранвье: 10 000/мм 2 , что в 200 раз больше, чем нервном волокне гигатского аксона кальмара. n Входящий Nа+-ток, пронизывающий невозбужденную мембрану в непосредственной близости от ее возбужденного участка в 5 -6 раз выше порогового тока. То есть фактор надежности настолько высок, что позволяет перескакивать ПД через несколько перехватов Ранвье, при их блокаде анестетиками. n
 – мякотные толстые моторные")
Классификация нервных волокон • Волокна типа А (ά, β, δ) – мякотные толстые моторные волокна, скорость проведения возбуждения до 120 м/сек. • Волокна типа В –тонкие мякотные волокна, чаще чувствительные, скорость проведения 3 -18 м/сек. • Волокна типа С – безмякотные, вегетативные, скорость проведения не больше 3 мсек.
 Миелинизация Скорость проведения")
Типы нервных волокон, их свойства и функциональное назначение Тип Диаметр (мкм) Миелинизация Скорость проведения (м/с) Функциональное назначение А 12– 20 сильная 70– 120 Двигательные волокна соматической НС; чувствительные волокна проприорецепторов А 5– 12 сильная 30– 70 Чувствительные волокна кожных рецепторов А 3– 16 сильная 15– 30 Чувствительные волокна проприорецепторов А 2– 5 сильная 12– 30 Чувствительные волокна терморецепторов, ноцицепторов В 1– 3 слабая 3– 15 Преганглионарные волокна симпатической НС С 0, 3– 1, 3 отсутствует 0, 5– 2, 3 Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов

• Составной потенциал действия подкожного нерва млекопитающего, зарегистрированный с помощью внеклеточного электрода. Все волокна нерва одновременно подвергаются стимуляции на некотором расстоянии от участка отведения. До участка отведения в первую очередь доходят потенциалы действия наиболее быстро проводящих волокон группы А; потенциалы действия медленных волокон группы С появляются примерно через 38 мс. Вслед за колебанием потенциала, соответствующим волокнам группы С, возникает продолжительный гиперполяризующий следовой потенциал. Отдельные зубцы волны, соответствующей А–волокнам, отражают активность волокон подгрупп α, β, γ, δ

Законы проведения возбуждения 1. 2. 3. 4. Закон физиологической непрерывности Закон двустороннего проведения Закон изолированного проведения Закон бездекрементного (незатухающего) проведения возбуждения 5. Закон относительной неутомляемости нерва (открыт Введенским)
 на электрическом органе нильского сома.")
Двустороннее проведение возбуждения экспериментально доказано: Бабухиным А. И. (1877) на электрическом органе нильского сома. Кюне В. (1886) на икроножной мышце лягушки.

Опыт Бабухина А. И. 1 2 3 4 5 6

Опыт Бабухина А. И. 1 2 3 разрез 4 разрез 5 6 раздражение

Опыт Кюне В. разрез ие ражен разд

Kopia_El_Genez_2-2_Potentsial_PP_i_PD.ppt