Большой практикум.ppt
- Количество слайдов: 32



Мобильные генетические элементы

Прыгающие генетические элементы В сороковые годы американский генетик Барбара Макклин-ток получила экспериментальные данные о том, что принцип постоянства размещения генов в хромосомах не абсолютен. что существуют участки хромосом, способные путешествовать из одного места генома в другое. Существенную роль в создании генетического разнообразия принимают участие так называемые мобильные диспергиро-ванные гены (МДГ). Они не имеют определенного фиксиро-ванного «адреса» в хромосомах и могут перемещаться по геному, что и отражено в их названии. Диспергированные — значит расположенные в разных местах. Мобильные — значит подвижные. В то время никто из биологов не верил в существование подвижных генов. Открытие Б. Макклинток признали позже, когда оно было подтверждено молекулярно-биологическими методами.

В 1977 г. , в Институте молекулярной биологии Ю. Ильин и Н. Чуриков использовали метод гибридизации ДНК. Цепи ДНК некоторых генов дрозофилы разделяли, метили радиоактивной меткой и гибридизировали с ДНК гигантских хромосом. Меченая ДНК находила на хромосомах комплементарную последовательность и связывалась с ней. Покрыв стекло, на котором находились хромосомы, фоточувствительной эмульсией определяли, с какими именно участками хромосом связалась меченая ДНК — в этих местах при проявлении проступали зерна серебра. Оказалось, что изучаемые гены разбросаны по разным участкам всех хромосом дрозофилы, то есть они множествен-ны. Неожиданным явилось то, что один и тот же ген у разных мушек по-разному локализовался в хромосомах — в некоторых случаях координаты совпадали, но в большинстве отличались. У мух разных линий отличия были велики, у родственников совпадений было больше, но около трети генов одного и того же семейства располагались совершенно вразнобой. В 1979 г. , существование подвижных участков генома под-

Таким образом, некоторые гены не имеют определенного места в хромосомах — у разных особей одного и того же вида дрозофил они могут занимать разные позиции. За несколько десятков или сотен поколений, разделяющих двух родственников, некоторые гены успевают сменить свое местоположение в геноме. Подвижные участки хромосом назвали «мобильными диспергированными генетическими элементами» , или МДГ. Используются и другие термины — прыгающие или кочующие гены. В геноме дрозофилы открыто около 20 семейств МДГ по 10— 150 копий в каждом. Общее число МДГ велико — около тысячи, на их долю приходится примерно 5 % всего генетического материала. Семейства были пронумерованы: МДГ-1, МДГ-2, МДГ-3, МДГ-4 и так далее. Общим у всех МДГ оказалось то, что на их концах располагаются одинаковые отрезки ДНК, называемые длинными концевыми повторами (ДКП). Кочующие гены состоят из 5— 10 тысяч нуклеотидных пар, из которых на концевые повторы приходится по 300— 600 пар. Концевые повторы играют большую роль в работе этих генов.

• В конце XX века стало очевидно, что подвижные гены присутствуют у всех организмов, включая человека. В каждый момент времени геном наших клеток можно рассматривать как комбинацию положений разнообразных подвижных генов на хромосомах. Количество таких возможных комбинаций, вероятно, очень велико. Вклад подвижных генов в функционирование и эволюцию организмов только начинает проясняться, но очевидно, что он может быть значительным. Мобильные диспергированные гены влияют на работу соседних с ними генов и вызывают наследуемые изменения — по сути, мутагенез. Оказалось, что в ДКП присутствуют все элементы управления работой генов: промотор, терминатор и усилитель. Более того, они есть в обеих цепях их ДНК, то есть длинные концевые повторы могут вызвать считывание информации в двух направлениях, по каждой цепи ДНК. Поскольку аппарат управления расположен на обоих концах МДГ, он может включить в работу не только мобильные элементы, но и соседствующие с ними гены.

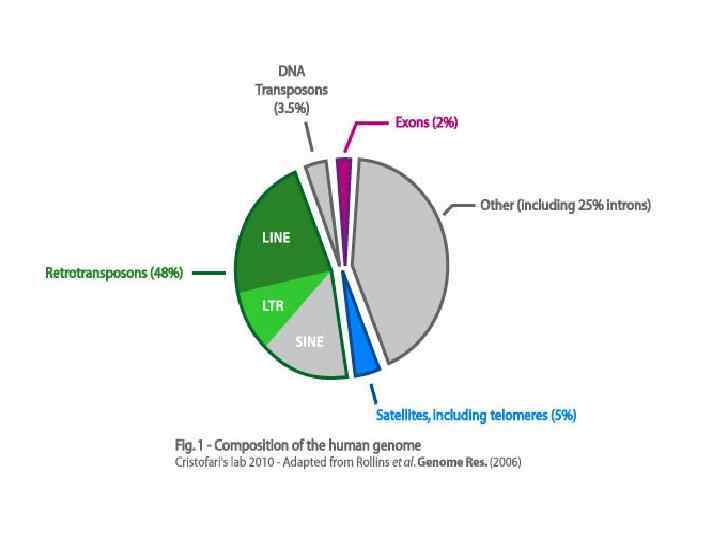
Повторы в геноме человека ~45% генома занято повторяющимися элементами 4 основнык класса повторов: • длинные диспергированные элементы (LINE) • короткие диспергированные элементы (SINE) • элементы с длинными концевыми повторами (LTR-элементы) -ретровирус-подобные эл. • ДНК-транспозоны Длина Число копий Доля генома LINE автономные 6 -8 kb SINE неавтономные 100 -300 п. н. 1 500 000 LTR-элементы авт. 6 -11 kb неавт. ДНК-транспозоны авт. неавт. 850 000 21% 13% 450 000 8% 300 000 3% 1. 5 -3 kb 2 -3 kb 80 -3000 п. н.

LINE способны к автономному перемещению в геноме наиболее древние мобильные элементы в геноме человека Длина: 6 -8 kb Структура: промотор ДНК-полимеразы II, две откр. рамки считывания (обратная. транскриптаза), поли-А-хвост Цикл: транскрипция - трансляция - миграция комплекса РНК-белки в ядро обратная транскрипция - интеграция Эволюционная стратегия: вертикальный перенос - транспозиция в геноме человека Классификация: 4 класса (LINE 1 - LINE 4), только LINE 1 способны к транспозиции SINE не способны к автономному перемещению в геноме могут использовать машину обратной транскрипции LINE-элементов Длина: 100 -400 п. н. Структура: промотор, два мономера (А и В), поли-А-хвост, нет ORF Эволюционная стратегия: вертикальный перенос - транспозиция в геноме человека Классификация: 3 класса: Alu-элементы, MIR, Ther 2/MIR 3, только Alu способны к транспозиции
 два типа: автономные и неавтономные большое количество «ископаемых» элементов, состоящих из")
LTR-элементы (ретровирус-подобные элементы) два типа: автономные и неавтономные большое количество «ископаемых» элементов, состоящих из изолированных LTR Длина: 6 -11 kb (автономные) и 1. 5 -3 kb (неавтономные) Структура: два прямых LTR-повтора на концах, 3 гена - gag, pol и env gag и pol - протеаза, обратная транскриптаза, РНК-аза, интеграза env - белок оболочки Цикл: транспозиция по ретровирус-подобному механизму: обратная транскрипция в цитоплазме в вирус-подобной частице (в отличие от LINE, где обр. транскр в ядре); вырезание и встраивание в геном с помощью гомологичной рекомбинации между LTR-повторами Эволюционная стратегия: горизонтальный перенос (от вида к виду) и вертикальный перенос (транспозиция в геноме человека) Классификация: 4 класса - 3 класса эндогенных ретровирусов млекопитающих (ERV I - ERV III) и Ma. LR

ДНК-транспозоны два типа: автономные и неавтономные сходны с транспозонами бактерий Длина: 2 -3 kb (автономные) и 80 - 3000 п. н. (неавтономные) Структура: инвертированные повторы на концах, ген транспозазы (обеспечивает транспозицию по механизму вырезание - вставка) Цикл: синтез транспозазы в цитоплазме вместе с другими белками человека, возвращение в ядро и вырезание транспозона. В отличие от LINE и LTR нет механизма цис-предпочтения (связывание белков с той молекулы РНК, с которой белок был считан) -> малоэфективнвая транспозиция (транспозаза не может отличить активные и неактивные транспозоны). Эволюционная стратегия: относительно частый горизонтальный перенос (от вида к виду) Классификация: как минимум 7 классов разных ДНк-транспозонов и их «ископаемых» остатков

На долю подвижных элементов у эукариот приходится значительная часть генома: у дрозофилы – 20%, у человека – около половины. Перемещение мобильных элементов находится под жестким контролем как со стороны самих элементов, так, по-видимому, и со стороны организмов-хозяев. Частота транспозиции достаточно низка – в среднем 10 -4 -10 -7 транспозиций на клетку за клеточную генерацию.

Разнообразные рекомбинационные процессы, лежащие в основе перемещений мобильных элементов, объединены под общим названием «транспозиции» . Транспозиции осуществляются особыми белками, гены которых, в основном, локализованы в самих мобильных элементах. Гомология между мобильным элементом и последовательностью ДНК, в которую он перемещается (ДНК-мишень), как правило, отсутствует. Встраивание элементов, как правило, происходит в случайные сайты ДНК-мишени. Для мобильных элементов характерно пребывание в составе хромосом или плазмид.

Большинство прокариотических мобильных элементов перемещается с помощью нерепликативной транспозиции. Нерепликативная транспозиция заключается в вырезании элемента и его перемещении в новое место. При этом 2 молекулы транспозазы связываются с концами мобильного элемента и делают разрывы одновременно в обеих цепях ДНК на концах мобильного элемента и в ДНК-мишени. Далее транспозаза сводит вместе концы мобильного элемента и ДНК-мишень, 3 -OH-концы элемента соединяются с 5 -Рконцами ДНК-мишени, а между 3’-OH-концами ДНК-мишени и 5’-Р- концами элемента образуется брешь, которая заполняется с помощью репаративного синтеза ДНК, в результате чего на концах мобильного элемента возникают ДПП строго фиксированной длины. В исходном репликоне остается ДНР. Будет ли он репарирован – зависит хозяйской клетки. Этот механизм характерен для большинства мобильных элементов бактерий и эукариотических элементов с короткими ИП. По такому типу перемещаются многие IS-элементы и мобильные элементы, которые называют составными: Tn 5, Tn 9, Tn 10 и другие. Составные транспозоны отличаются тем, что у них инвертированные повторы представлены ISэлементами, которые находятся в обратной или (гораздо реже, например, Tn 9) в прямой ориентации.

Схема, демонстрирующая общий принцип реакций транспозиции репликативная транспозиция нерепликативная транспозиция

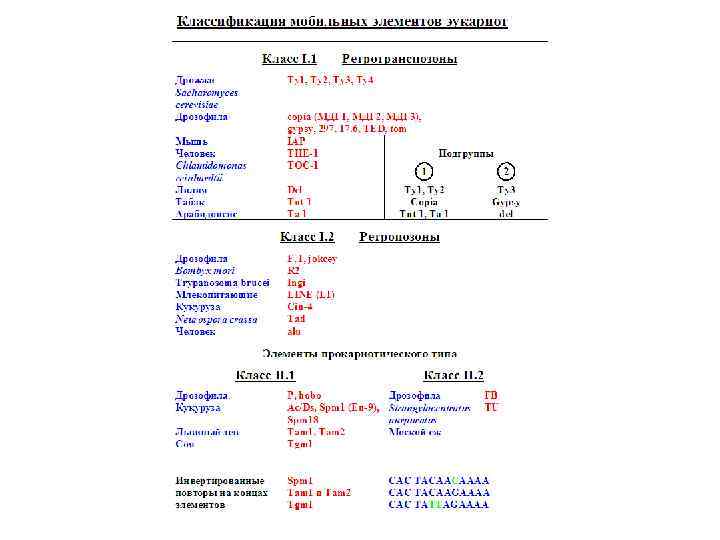
Основные типы мобильных элементов Структура Ключевой ген транспозиции Способ перемещения Примеры Кодирует транспозазу В виде ДНК, репликативная или нерепликативная транспозиция E coli. : IS 1, Tn 3 Кукуруза: Ac-Ds, Spm 1 Соя: Tgm 1 Львиный зев: Tam 1 Дрозофила: P, hobo Кодирует транспозазу В виде ДНК, нерепликативная транспозиция E. coli: Tn 5, Tn 9, Tn 10 Дрозофила: FB Кодирует обратную транскриптазу Через РНКинтермедиат Дрожжи: Ty 1, Ty 2, Ty 3 Кукуруза: Bs 1 Лилия: del Дрозофила: copia, МДГ 1, МДГ 2, gypsy Мышь: IAP Человек: THE 1 Через РНКинтермедиат Кукуруза: cin 4 Трипаносома: ingi Дрозофила: F, I Мышь: L 1 Md Человек: L 1 Hs

МОБИЛЬНЫЕ ГЕНЕТИЧЕСКИЕ ЭЛЕМЕНТЫ ПРОКАРИОТ: IS-элементы, транспозоны Для бактерий и плазмид характерны мобильные элементы с короткими или длинными ИП. Длина ДПП, как правило, 5 или 9 п. н. Бактериальные мобильные элементы можно разделить на две основные группы: 1. IS-элементы: небольшие (размером не более 2, 5 т. п. н. ) элементы, которые состоят из центральной части с геном транспозазы, фланкированной двумя инвертированными повторами. 2. Собственно транспозоны, которые несут, кроме транспозазы, другие гены, не имеющие отношения к транспозиции (чаще всего гены устойчивости к антибиотикам). Собственно транспозоны можно в свою очередь разделить на следующие группы 1) Сложные транспозоны (семейство Tn 3) – короткие ИП на концах, делают в ДНК-мишени ДПП из 5 п. н. и перемещаются по механизму репликативной транспозиции. 2) Составные транспозоны (Tn 5, Tn 9, Tn 10) с длинными ИП, представляющими собой различные IS-элементы. Длина ДПП обычно 9 п. н. Примеры прокариотических мобильных элементов приведены в следующей ниже таблице.

Общая структура мобильного элемента: В состав тела транспозона могут входить: • функциональный или «испорченный» ген транспозазы • ген резолвазы • ген устойчивости к антибиотикам • гены дополнительных ферментов клеточного метаболизма
, кодирующие транспозазу короткие концевые инвертированные повторы Дупликация")
IS-элементы: 750 -1600 п. о. ген (гены), кодирующие транспозазу короткие концевые инвертированные повторы Дупликация ДНК-мишени, возникающая при встраивании IS-элемента

Ферменты, которые способны синтезировать гены транспозона: • Транспозаза – фермент, катализирующий вырезание и интеграцию мобильного элемента • Резольваза – фермент, осуществляющий разделение коинтеграта на исходные репликоны Коинтеграт: resolvase
 и")
У эукариот широко распространены ретротранспозоны, в транспозициях которых задействованы фермент обратная транскриптаза (ревертаза) и РНК-копия элемента в качестве интермедиата. Ретроэлементы подразделяются на 2 группы: 1. Ретротранспозоны с длинными прямыми концевыми повторами (ДКП). Их структура соответствует ДНК-копиям геномов ретровирусов позвоночных, которые также являются мобильными элементами. 2. Ретроэлементы, не содержащие повторов на концах (некоторые авторы используют для них название «ретропозоны» ).
 8 11 4560 п. н. •")
Транспозоны эукариот: • автономные (Ac из Zea mays) 8 11 4560 п. н. • неавтономные (Ds из Zea mays) мутация гена, кодирующего транспозазу 11 8 Эксплуатация транспозазы

Перемещение мобильного элемента LINEтипа

Fig. 7. 24, Transposon effects on corn kernel color.

К этому же типу транспозонов относятся многие мобильные элементы растений: элементы Spm кукурузы, Tgm 1 сои, Tam 1 и Tam 2 львиного зева и др. Отметим двухкомпонентную систему Ac/Ds кукурузы (это самый первый обнаруженный мобильный элемент, описанную Барбарой Мак-Клинток): она включает автономно транспозирующийся элемент Ас (4565 п. н. , ИП из 11 п. н. , ДПП из 8 п. н. , ген транспозазы содержит 4 интрона) и гетерогенные по длине элементы Ds, которые являются делетированными производными Ас-элемента и перемещаются с помощью его транспозазы.

P-элемент Drosophila melanogaster 8 31 2900 п. н. 31 pre-m. RNA сплайсинг 5’ 3’ м. РНК транспозазы 8

Последствия транспозиции: Изменение окраски - последствия транспозиции:

Ретровирусы являются «прототипами» ретротранспозонов. Их цикл развития состоит из чередования РНК- и ДНК-стадий. Вирионный геном представлен РНК размером обычно 5 -6 т. п. н. с короткими прямыми повторами. Когда ретровирус проникает в клетку хозяина, то с помощью кодируемой им обратной транскриптазы на матрице его РНК синтезируется ДНК-копия, но уже с ДКП (в англоязычной литературе LTR – long terminal repeats) длиной обычно 200 -400 п. н. ДКП содержат двунуклеотидные инвертированные повторы на концах и еще ряд повторов на некотором расстоянии от концов, разнообразные регуляторные элементы (промоторы и терминаторы и энхансеры транскрипции). Наличием регуляторных элементов в ДКП обусловлены различные эффекты ретровирусов и ретротранспозонов, встроенных в хромосомы, на экспрессию соседних генов.

• Центральная часть ретровируса содержит 3 кодирующие рамки: • gag – кодирует структурный белок вирионного капсида; • pol – кодирует сложный полипептид, в котором слиты домены интегразы (ответственна за интеграцию ДНК-копии в хозяйский геном; интеграза соответствует транспозазе других подвижных элементов), обратной транскриптазы (ревертазы), РНКазы H (RNAse H удаляет РНК из гибрида ДНК-РНК) и протеазы (после транскрипции слитого полипептида протеаза «нарезает» его на отдельные функциональные полипептиды). • Env – белки хвостового отростка вируса, которые ответственны за адсорбцию ретровируса на поверхности клетки-хозяина и, соответственно, его вирулентность. Большинство ретровирусов не содержат гена env и, следовательно, неинфекционны.
,")
В последние годы А. И. Ким и др. открыли, что мобильный элемент МДГ-4 (gypsy), содержит ген env и обладает инфекционными свойствами. Затем французские исследователи выявили у дрозофилы аналогичные элементы ZAM, Idefix и др. , всего более 10. Таким образом, стало известно, что ретровирусы встречаются не только у позвоночных животных. Новые вирусы выделены в отдельную группу Errantiviruses – эндогенные ретровирусы беспозвоночных. У многих ретровирусов рамки считывания gag и pol перекрываются (а иногда они «сливаются» в общий транскрипт). Транспозоны из обеих групп встречаются среди всех групп живых организмов – от дрожжей до человека. Ретротранспозоны всегда делают в ДНК-мишени ДПП из 5 п. н.

ИНТЕРЕСНЫЕ ФАКТЫ Гомеозисная мутация antennapedia У представителей рода Drosophila, D. melanogaster и D. virilis теломеры, в отличие от других организмов, формируются путем последовательных транспозиций двух элементов LINE-типа: He. TA и TART. Ретровирус HIV-1 вызывает у человека синдром иммунодефицита.

СПАСИБО ЗА ВНИМАНИЕ!
Большой практикум.ppt