Тема_4_Нервный_импульс.ppt
- Количество слайдов: 42



Министерство образования Российской Федерации Международный университет природы, общества и человека «Дубна» Кафедра клинической психологии Тема 4. Проведение нервного импульса и нервно- мышечная передача
. Особенности метаболизма нейронов: Высокое потребление")
Основной структурно-функциональной единицей нервной системы является нервная клетка (нейрон). Особенности метаболизма нейронов: Высокое потребление О 2. Полная гипоксия в течение 5 -6 минут ведет к гибели клеток коры. Способность к альтернативным путям обмена . Способность к созданию крупный запасов веществ. Нервная клетка живет только вместе с глией. Способность к регенерации отростков (0, 5 -4 мк/сут).

Структура нейрона

нервные волокна – это отростки нейронов, имеющие оболочку и способные проводить нервный импульс все нервные волокна делятся на две основные группы - миелиновые (мякотные волокна) и безмиелиновые (безмякотные).

Строение миелинового нервного волокна 1 — осевой цилиндр 2 — миелиновый слой оболочки волокна 3 — цитоплазма леммоцита (шванновской клетки) 4 — ядро леммоцита (шванновской клетки) 5 — базальная мембрана

Схема строения миелинового нервного волокна 1 - перехват Ранвье 2 - шванновская оболочка 3 - шванновская клетка 4 - осевой цилиндр 5 - миелиновая оболочка

Строения безмиелинового нервного волокна 1 — ядро шванновской клетки 2 — осевые цилиндры 3 — мезаксон 4 — базальная мембрана

Потенциал действия – универсальный ответ нервной клетки на стимуляцию Длительность ПД на схеме составляет 1 мс. По ходу ПД можно выделить восходящую и нисходящую фазы (примерно по 0. 5 мс каждая) Восходящая фаза вход Na+ выход К+ (деполяризация): вход в клетку «порции» Na+. нисходящая восходящая Нисходящая фаза (реполяризация): выход фаза из клетки примерно такой же «порции» К+. ПП

Механизм распространения возбуждения по безмиелиновому нервному волокну

Механизм распространения возбуждения по миелиновому нервному волокну

Преимущества прерывистое распространение возбуждения Во-первых, скорость распространения возбуждения в волокнах, покрытых миелином, в 8 -10 раз быстрее, чем в безмякотных (безмиелиновых). Во-вторых, на распространение возбуждения прерывистого типа затрачивается меньше энергии, оно более экономично, что связано с большой плотностью натриевых каналов в перехватах Ранвье.
 зация ния")
Типы нервных волокон Скорость Диаметр Миелини- Тип проведе- Функциональное назначение (мкм) зация ния (м/с) Двигательные волокна А соматической НС; 12– 20 сильная 70– 120 alpha чувствительные волокна проприорецепторов А Чувствительные волокна кожных 5– 12 сильная 30– 70 beta рецепторов А Чувствительные волокна 3– 16 сильная 15– 30 gamma проприорецепторов А Чувствительные волокна 2– 5 сильная 12– 30 delta терморецепторов, ноцицепторов Преганглионарные волокна В 1– 3 слабая 3– 15 симпатической НС Постганглионарные волокна cимпатической НС;

Нервные волокна всех групп обладают общими свойствами: практически неутомляемые, так как процессы ресинтеза энергии в нем идут с достаточно большой скоростью и успевают восстановить траты энергии, происходящие при прохождении возбуждения; обладают высокой лабильностью, т. е. могут воспроизводить потенциал действия с очень высокой частотой.
 Нерв проводит возбуждение")
Законы распространения возбуждения по нерву Закон целостности нерва (закон непрерывности) Нерв проводит возбуждение только в том случае, если он сохраняет свою морфологическую и функциональную целостность. Закон двустороннего проведения возбуждения Нервное волокно способно проводить возбуждение от рецепторов к центрам и наоборот Закон изолированного распространения возбуждения Потенциал действия в волокнах, покрытых миелином, не перебрасывается с одного нервного волокна на другое благодаря хорошим изоляционным свойствам миелина

Восстановление нервного волокна А — волокно до повреждения; Б — в периферическом отрезке аксон дегенерирует, клетки в месте повреждения пролиферируют; В — регенерация аксона в центральном отрезке, прорастание веточек аксона в периферический отрезок; Г — полная регенерация нервного волокна и восстановление связей.

Электрические свойства нейронов. Потенциал покоя и потенциал действия. синапсы аксон клетка- мишень Сигнал по мембране нейрона передается в виде коротких элект- рических импульсов – потенциалов действия (ПД). Этот процесс можно сравнить с передачей информации с помощью включения и выключения фонарика (ПД = «вспышка света» ). Но для того, чтобы фонарик работал, нужна батарейка – источник электрической энергии. В случае нейрона таким источником является постоянный внутриклеточный заряд – 16 потенциал покоя (ПП).
 нейрона – его постоянный отрицательный заряд, равный в среднем -70")
Потенциал покоя (ПП) нейрона – его постоянный отрицательный заряд, равный в среднем -70 м. В. Измерить ПП можно с помощью тончайшей, особым образом вытянутой стеклянной трубочки-микроэлектрода. Его кончик имеет диаметр < 1 мкм, что позволяет практически без повреждения проткнуть мембрану клетки. показания вольтметра, м. В Микроэлектрод (в 0 т. ч. канал внутри кончика) заполнен раствором соли, проводящим эл. ток. Это позволяет -70 ПП оценить сравнить заряд цитоплазмы нейрона с зарядом межклеточной время, среды). момент мин вольтметр прокола мембраны 17

Наличие ПП – результат жизнедеятельности нейрона, совместного функционирования всех биополимеров и органоидов клетки; погибший нейрон быстро теряет ПП. Первопричина ПП – разность концентраций ионов K+ и Na+ внутри и снаружи нейрона. Эту разность создает работа особого белка-насоса Na+-K+-АТФазы (Na+-К+-насоса). Na+ межклеточная среда К+ АТФ Na+-K+-АТФаза обменива- ет находящиеся внутри АДФ клетки ионы Na+ на захва- ченные в межклеточной Na+ среде ионы К+, затрачивая значительное кол-во АТФ. К+ внутриклеточная 18 среда

В результате работы Na+-K+-АТФазы в нейроне оказывается примерно в 10 раз меньше Na+ и в 30 раз больше К+, чем в межклеточной среде. К+out : К+in = 1 : 30 Na+out : Na+in = 10 : 1 Несмотря на все это, до момента созревания (происходит на 2 -3 месяце эмбрионального развития) нейрон не имеет заряда, и количество положительных (прежде всего, К+) и отрица- тельных ионов в его цитоплазме примерно одинаково. Признак созревания – появление на мембране нейрона постоянно открытых К+-каналов (опреде- ляется включе- нием соотв. гена). В результате стано- вится возможной диффузия К+ внутриклеточная 19 из клетки. среда
")
Как долго идет диффузия К+ из нейрона? Очевидный вариант ( «до выравнивания концентраций» ) неверен, поскольку двигаются заряженные частицы, и выход К+ сопро- вождается накоплением в цитоплазме отрицательного заряда. Этот отрицательный ПП – это отрицате- заряд мешает диф- льный заряд цито- фузии и в конце плазмы, останавли- концов останавлива-ет вающий диффузию её. Возникает ионов К+ в межкле- состояние «динами- точную среду. ческого равновесия» : число ионов К+, покинувших клетку благодаря диффузии = числу ионов К+, втянутых в клетку отрицательным за- рядом цитоплазмы. внутриклеточная 20 среда
 коэффициент пропорциональности")
«Уравнение Нернста» : ПП ~ lg ( К+out / К+in ) коэффициент пропорциональности равен 61. 5 м. В для Т=36. 6°С; логарифм равен -1. 48 (для соотно- шения концентраций 1/30). ПП – это отрицате- С учетом этого ПП = -91 м. В льный заряд цито- ( «равновесный потенциал» для К+) плазмы, останавли- вающий диффузию ионов К+ в межкле- точную среду. Вальтер Нернст внутриклеточная 21 (Ноб. пр. 1921) среда

Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название – «ток утечки Na+ » ). ПП = -91 м. В ( «равновесный потенциал» для К+) В реальной клетке ПП находится ближе к нулю (в среднем -70 м. В). Причина: существование небольшого количества постоянно открытых каналов для ионов Na+. Избыток ионов Na+ в межклеточной среде, а также их притяжение к отрицательно заряженной цитоплазме приводят к входу Na+ в клетку. внутриклеточная 22 среда

Такой вход Na+ ведет к сдвигу заряда цитоплазмы вверх и частичной потере ПП (отсюда название – «ток утечки Na+ » ). Ограничивает вход Na+, во-первых, малое число постоян-но открытых Na+-кана- лов; во-вторых, работа Na+-K+ - АТФазы, кото- рая «откачивает» Na+ , обменивая его на К+ В целом ПП зависит от 3 -х главных факторов: - диффузии K+ из клетки; - диффузии Na+ в клетку; внутриклеточная 23 - работы Na+-K+-АТФазы. среда

Такой вход K+ изведет к сдвигу заряда цитоплазмы вверх Диффузия Na клетки определяется разностью + и частичной потере ПП (отсюда название – «ток утечки» Na+). концентраций К+out и К+in. Если увеличить К+out , то разность концентраций станет меньше, диффузия – слабее, и для ее остановки потребуется не столь значительный ПП (произойдет сдвиг заряда цито- Этот график плазмы вверх до достижения новой точки равновесия). можно по- лучить в Если снизить К out , то раз- + эксперимен- ность концентраций станет показания те, но в ре- вольтметра, м. В больше, диффузия – силь- альном мозге нее, и для ее остановки по- в норме такого требуется более значитель- 0 не происходит ный ПП (сдвиг заряда цито- плазмы вниз). В целом ПП зависит от 3 -х ПП главных факторов: - диффузии K+ из клетки; время, мин - диффузии Na+ в клетку; увеличение снижение 24 - работы Na+-K+-АТФазы. К+out

Такой вход Na++ в клетку сдвигу заряда цитоплазмыконцентрации Диффузия Na ведет к зависит, прежде всего, от вверх + и частичной потере ПП + (отсюда название – «ток утечки» Na ). постоянно открытых Na -каналов на мембране. Эта величина, в свою очередь, является стабильным свойством конкретного нейрона. Чем больше таких каналов, тем ПП ближе к нулю, чем меньше – тем ПП ближе к уровню -91 м. В. Чем ближе ПП к нулю, тем А: нейрон с большим числом показания возбудимее нейрон (такие вольтмет- Na+-каналов нужны, например, в центрах ра, м. В Б: нейрон со средним бодрствования); числом Na+-каналов чем ближе ПП к уровню -91 м. В, 0 (ПП= -70 м. В) тем ниже возбудимость (мини- В: нейрон с малым числом мальна в центрах, запускаю- Na+-каналов щих движения). В целом ПП зависит от 3 -х ПП нейрона А -60 главных факторов: ПП нейрона Б - диффузии K+ из клетки; -70 - диффузии Na+ в клетку; -80 ПП нейрона В 25 - работы Na+-K+-АТФазы. время, мин
 — место контакта между двумя нейронами")
Синапс (в переводе с греческого обнимать, обхватывать) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторой клеткой. Служит для передачи нервного импульса между двумя клетками.

Классификация синапсов I. По способу II. По местоположению и принадлежности передачи сигнала: соответствующим структурам: Химические a) периферические Электрические нервно-мышечные нейросекреторные (аксо-вазальные) Смешанные рецепторно-нейрональные b) центральные аксо-дендритические — с дендритами, в т. ч. III. В зависимости от аксо-соматические — с телами нейронов; медиатора: аксо-аксональные — между аксонами; аминергические, в дендро-дендритические — между дендритами; т. ч. адренергические IV. По характеру действия на холинергические воспринимающую структуру: пуринергические Возбуждающие пептидергические Тормозящие

Основные элементы синапса
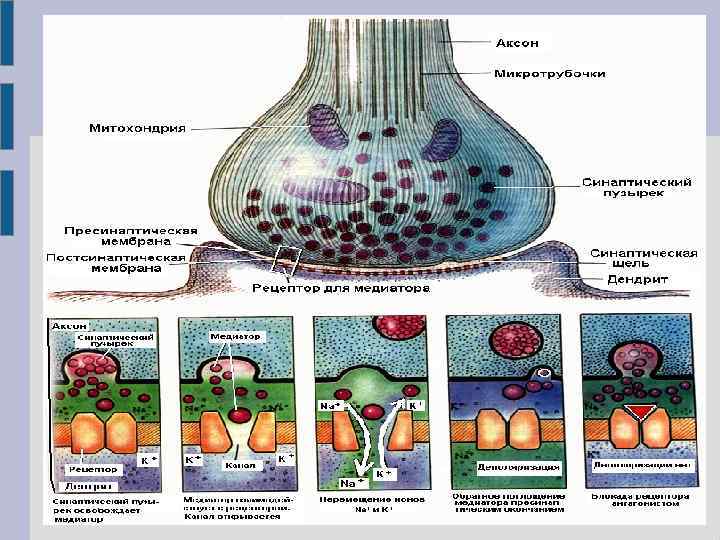

Мионевральный синапс
 в 1897 г. на")
СИНАПС Представление о синапсе сформулировано Чарльзом Шеррингтоном (Ch. Sherrington) в 1897 г. на основе изучения нервно-мышечных контактов. АЦЕТИЛХОЛИН (Ацх) Первый открытый медиатор; в ходе изучения эффектов Ацх Отто Лёви (O. Löwi) сформулировал идею химического синапса (1921 г. ). Блуждающий нерв Симпатический нерв 31

Главное «действующее лицо» в синаптической передаче – медиатор. Медиатор проходит в синапсе полный «жизненный цикл» , включающий 4 этапа: синтез и накопление в пресинаптическом окончании; выброс в синаптическую щель при появлении ПД; действие на рецепторы постсинаптической мембраны (запуск возбуждения или торможения постсинаптической клетки); инактивация (прекращение действия медиатора на рецептор). 32
, затем")
АЦЕТИЛХОЛИН Сначала была доказана его роль в работе ВНС (вегетативной нервной системы), затем – нервно-мышечных синапсов, позже – ЦНС (центральной нервной системы). В 1936 г. Отто Лёви и Генри Дейл (H. Dale) получили Ноб. Пр. за «открытие химического механизма синаптической передачи» . Какой он – АЦЕТИЛХОЛИН ? «Ацетил» – остаток уксусной кислоты СН 3 -СООН «Холин» – атом азота N, с которым соединены три Блуждающий нерв группы -СН 3 и одна группа этилового спирта -СН 2 -ОН Симпатический нерв 33

П 1 П 2 Мед П 1 – остаток уксусной кислоты, сое- диненный с коферментом А (Со. А). 2 П 2 – холин ( «витаминоид» : полу- АТФ чаем только с пищей). 1 Мед – ацетилхолин (Ацх); фермент: холин ацетил-трансфераза. Синтез – в пресинаптическом окон- чании, после чего Ацх переносится 3 внутрь везикул и готов к экзоцитозу. «Ацетил» – остаток уксусной кислоты СН 3 -СООН «Холин» – атом азота N, с которым соединены три Блуждающий нерв группы -СН 3 и одна группа этилового спирта -СН 2 -ОН Симпатический нерв 34
.")
П 1 – остаток уксусной кислоты, сое- диненный с коферментом А (Со. А). П 2 – холин ( «витаминоид» : полу- Ацх пресинапти- чаем только с пищей). ческое окончание Мед – ацетилхолин (Ацх); фермент: холин ацетил-трансфераза. ацетил-Со. А холин ацетил- трансфераза Синтез – в пресинаптическом окон- чании, после чего Ацх переносится внутрь везикул и готов к экзоцитозу. Ацх Появление ПД запускает выброс Ацх в синаптическую щель, после чего синапти- он действует на рецепторы ческая щель постсинаптической мембраны. Известны 2 типа рецепторов к Ацх: рецептор Ацх первый из них реагирует на Ацх и агонист никотин (токсин табака); второй реагирует на Ацх и агонист 35 мускарин (токсин мухомора).

Как может один и тот же медиатор действовать на несколько типов рецепторов? Как правило, это означает, что медиатор ( «ключ» ) разными частями своей молекулы соединяется с разными активными центрами рецепторов. медиатор рецептор 1 -го типа рецептор 2 -го типа Но если активные центры рецепторов разные, то агонисты Известны 2 типа рецепторов к Ацх: и антагонисты также будут различаться. Классический первый из них реагирует на Ацх и антагонист Ацх-рецепторов первого типа ( «никотиновых» ) – агонист никотин (токсин табака); курарин; классический антагонист Ацх-рецепторов второй реагирует на Ацх и агонист второго типа ( «мускариновых» ) – атропин 36 (курарин и атропин –мускарин (токсин мухомора). Ацх-подобные растит. токсины).
 Из представленных")
Он является главным медиатором Ацх-нейроны периферической НС. в ГМ (около 5%) Из представленных на рисунке пяти нейро- нов, образующих волокна периф. НС, четыре в качестве медиатора используют Ацх. (1) мотонейрон (2) симпатический пре- ганглионарный н. 1 (3) парасимпатический мото- преганглионарный н. нейрон мышца (4) парасимпатический постганглионарный н. И лишь симпатический симпа- 2 5 постганглионарный н-н (5) тическ. выделяет медиатор нейрон норадреналин. пара- симпа- (напр. , 4 внутр. мочевой тическ. 3 орган пузырь) нейрон 37

нервное окончание Инактивация Ацх проис- ходит с помощью фермента ацетилхолинэстеразы. Ацх-эстераза расположена на белок- постсинаптической мембране и «насос» холина в синаптической щели. Она очень быстро «разрывает» Ацх на холин и остаток уксусной кислоты (ацетат). холин На следующем шаге холин пе- ацетилхолинэстераза реносится с помощью особого ацетат белка-насоса обратно в преси- наптическое окончание и вновь холин используется для синтеза Ацх. Блокаторы Ацх-эстеразы активируют передачу сигнала в ацетилхолиновых синапсах, вызывая в больших дозах судороги (нервно-мышечные синапсы), спазм бронхов и 38 остановку сердца (парасимпатические синапсы).
 медиатора в синаптическую щель происходит после появления ПД, который вызывает открывание электрочувствительных")
Выброс (экзоцитоз) медиатора в синаптическую щель происходит после появления ПД, который вызывает открывание электрочувствительных Са 2+-каналов (примерно на 2 -3 мс). В результате в пресинаптическое окончание успевает войти несколько сот ионов Са 2+ , которые активируют белки, запускающие экзоцитоз. Для экзоцитоза одной везикулы требуется несколько (не < 4 -х) ионов Са 2+. Особые белки- насосы быстро удаляют Са 2+ из пресинаптического окончания (как в случае клеток серд- ца), иначе выброс медиатора не пре- кратится. Приход одного ПД в среднем вызывает выброс содержимого примерно 50 везикул. 39

Несколько дополнений: Увеличение концентрации Са 2+ в межклеточной среде ведет к его более активному входу в пресинаптическое окончание и росту выброса медиатора (Са. Cl 2 = хлорид кальция – мягкий стимулятор работы нервных и мышечных клеток, сердца). Ионы Mg 2+ способны проникать Бактерия ботулизма – почвен- через Са 2+-каналы, но не акти- ный микроб, анаэробный (не вируют белки, запускающие выносит О 2). Ее токсин блоки- экзоцитоз. Добавка Mg 2+ рует белки, отвечающие за в среду ведет к снижению экзоцитоз; отравление (если входа Са 2+ и падению выброса бактерия оказалась в консервах) медиатора (Mg 2+ конкурирует с ведет к слепоте, параличам и Са 2+ за вход в окончание аксона; смерти. Но БОТОКС используют Mg. SO 4 = магнезия – тормозит в клинике и косметологии работу синапсов и сердца, (блокада нервно-мышечных снижает тонус сосудов). 40 синапсов, снятие спазма мышц).
 является элементарной функциональ-")
И наконец… Еще раз подчеркну, что именно синапс (а не нейрон) является элементарной функциональ- ной единицей нервной системы. Чем больше синапсов, тем «умнее» мозг. «Легкий» мозг может иметь бóльшую плотность синапсов, чем «тяжелый» , и обладать существенно бóльшими возможностями (вóрон / собака). Нервные клетки не восстанавливаются (почти…). Но синапсы – образуются и исчезают; активно работающие нейроны формируют новые контакты, а «бездельники» – теряют их; поврежденные нейроны могут формировать новые синапсы. . . «По ходу жизни» многие синапсы способны изменять свои свойства, снижая либо увеличивая выделение медиатора и число рецепторов в ответ на определенные сигналы и ситуации. Эта способность, называемая пластичностью синапсов, лежит в основе обучения, созревания и старения мозга, развития многих видов нервных и психических отклонений. 41

Физиологические свойства нервно-мышечного синапса Одностороннее проведение возбуждения (от пре- к постсинаптической мембране. Синаптическая задержка проведения возбуждения (время между приходом импульса в пресинаптическое окончание и началом постсинаптического ответа). Низкая лабильность и высокая утомляемость синапса, обусловленная временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности. Высокая избирательная чувствительность синапса к химическим веществам, обусловленная специфичностью хеморецепторов постсинаптической мембраны.
Тема_4_Нервный_импульс.ppt