Тема_5 мышцы.ppt
- Количество слайдов: 49



Министерство образования Российской Федерации Международный университет природы, общества и человека «Дубна» Кафедра клинической психологии Тема 5. Мышечное сокращение © Дорофеева Саглара Михайловна

Типы мышечной ткани 1. Скелетные, поперечнополосатые 2. Сердечная мышца с характерной исчерченностью 3. Гладкие мышцы, образующие стенки полых внутренних органов и сосудов

Морфо‑функциональные различия типов мышечной ткани Основные типы мышечных волокон. 1 - волокно поперечнополосатой скелетной мышцы, 2 – миокардиоцит, 3 - гладкий миоцит.
 двигательная (динамическая и статическая); 2) обеспечения дыхания; 3) мимическая; 4)")
Функции поперечнополосатых мышц: 1) двигательная (динамическая и статическая); 2) обеспечения дыхания; 3) мимическая; 4) рецепторная; 5) депонирующая; 6) терморегуляторная. Функции гладких мышц: 1) поддержание давления в полых органах; 2) регуляция давления в кровеносных сосудах; 3) опорожнение полых органов и продвижение их содержимого. Функция сердечной мышцы – насосная, обеспечение движения крови по сосудам
. Формируется за счет дисков А, в обыкновенном")
Основные физические свойства: • Двоякое лучепреломление (анизотропия). Формируется за счет дисков А, в обыкновенном свете выглядят темными, а в поляризованном - светлыми, если свет пропускается в продольном направлении, и темными, если он проходит в поперечном направлении. • Растяжимость. • Эластичность. Заключается в том, что после растяжения мышца приходит в исходное положение. • Упругость. Это свойство мышцы связано с ее сжатием. После сжатия мышца способна приходить в исходное состояние. • Пластичность. Заключается в том, что мышца способна некоторое время сохранять приданную ей искусственную форму.

Физиологические свойства: 1. Возбудимость– это свойство клеточных 2. 3. 4. мембран отвечать на действие раздражителя изменением ионной проницаемости мембраны и величины мембранного потенциала Проводимость - способность ткани проводить возбуждение в виде потенциала действия Автоматизм - способность сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом Сократимость - способность мышцы укорачиваться, уменьшая свою длину или напрягаться без ее изменения

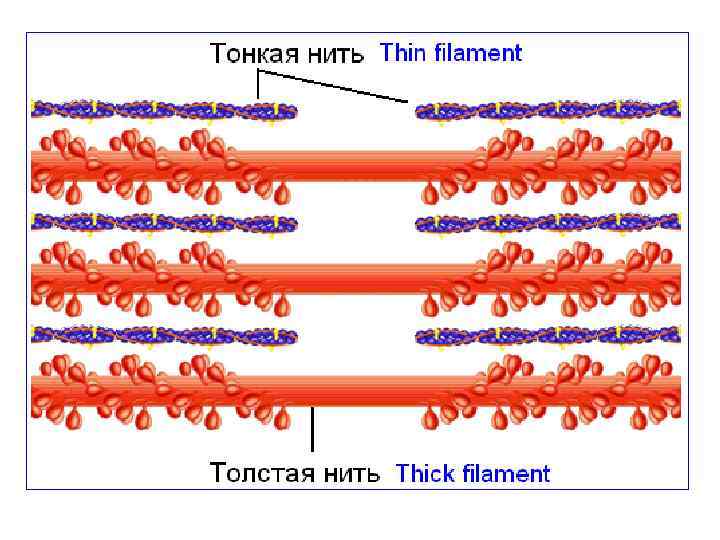
Иерархия структурных сократительных компонентов скелетной мышцы 1. Мышца 2. Мышечное волокно 3. Миофибрилла 4. Миофиламенты (актиновые и миозиновые нити)

Упрощённая схема саркомера

Расположение миофиламентов на поперечном срезе миофибриллы • Электронная микрофотография поперечного среза группы миофибрилл волокна скелетной мышцы (H. E. Haxley J. Mol. Biol. , 37: 507520. , 1968).

1. Молекулы миозина в растворе спонтанно образуют волокна. 2. Миозин обладает ферментативной активностью, гидролиз АТФ является непосредственным источником свободной энергии, необходимой для сокращения. 3. Миозин связывает актин - основной компонент тонких нитей.

С нитями актина связаны регуляторные белки мышечной ткани - тропомиозин и тропонин

Миофиламенты

Строение моторной единицы

Критерии классификации скелетных мышечных волокон и мышц • • По расположению – интрафузальные и экстрафузальные Характеру сокращения - фазные и тонические Скорости сокращения – медленные (развивают максимальное напряжение в течение 70 -100 мс) и быстрые (1020 мс) Механизму ресинтеза АТФ – окислительные (красные) и гликолитические (белые)

Экстрафузальные и интрафузальные мышечные волокна • Интрафузальные мышечные волокна вместе c чувствительными нервными окончаниями формируют мышечные веретёна. • Под световым микроскопом мышечное веретено представляет собой вытянутую структуру, расширенную посередине за счет капсулы и напоминающую по форме веретено, что и обусловило ее название. • Экстрафузальные мышечные волокна образуют основную массу мышцы и выполняют всю работу, необходимую для движения и поддержания позы.

• Фазные моторные единицы генерируют потенциал действия. Они представлены мотонейронами спинного мозга, которые образуют на мышечном волокне один - два синапса. • Тонические моторные единицы представлены -мотонейронами спинного мозга, образующими на мышечном волокне 10 -12 синапсов. Отсутствие возбудимых структур на этих волокнах не позволяет генерировать потенциал действия. Они способны на нервных окончаниях формировать только локальный ответ.

• Быстрые моторные единицы развивают максимальное напряжение в течение 1020 мс, медленные - в течение 70 -100 мс. В спинном мозге они представлены крупными, высоковозбудимыми L 1 мотонейронами. • Медленные моторные единицы развивают длительные и сильные мышечные сокращения и утомляются медленнее (красные мышцы). Они представлены мелкими, низковозбудимыми L 2 мотонейронами спинного мозга.

Сводная классификация мышечных волокон I тип медленные мышечные волокна II тип быстрые мышечные волокна Окислительные IIа тип Окислительные (красные) Неутомляемые Мало утомляемые IIб тип Гликолитичес кие Быстро утомляемые

Композиция скелетных мышц • В зависимости от преобладания в мышцах конкретного типа мышечных волокон скелетные мышцы относят к «красным» и «белым» либо «быстрым» и «медленным» . • Общая физиологическая характеристика мышцы – сила, скорость сокращения, выносливость – в большой мере определяется процентным соотношением в мышце типов волокон.

В мышечной ткани наблюдаются две фазы тепловых изменений: 1. Начальное теплообразование (30% всего образующегося тепла) не сопровождается потреблением кислорода (бескислородная фаза). Ø период активации (в этом периоде образуется тепло, связанное с переходом ткани от невозбужденного состояния к возбужденному), Ø период теплообразования, связанный с сокращением мышцы, Ø период теплообразования в период расслабления мышцы. 2. Запаздывающее теплообразование (восстановительное, 70 % тепла), сопровождается потреблением кислорода (кислородная фаза)

При возбуждении в мышцах имеют место метаболические изменения: Ø распад и ресинтез макроэргических соединений (АТФ, АДФ, креатинфосфат), Ø усиление гликолиза с накоплением лактата, Ø синтез и выделение БАВ (медиаторы, моноамины и др. ), Ø усиливается потребление кислорода и выделение углекислого газа, Ø усиливается распад жиров и углеводов как источников энергии.

Теория мышечного сокращения Механизм мышечного сокращения объясняется моделью скользящих нитей, авторами которой принято считать Х. Хаксли и Дж. Хансона (1954 г. ). Huxley H. E. , Hanson J.
 Длина как толстых, так и тонких нитей в ходе")
При исследовании удалось выявить: 1) Длина как толстых, так и тонких нитей в ходе мышечного сокращения не меняется. 2) В то же самое время длина дисков А, соответствующих миозиновым нитям, не меняется, а размеры дисков I, соответствующие актиновым нитям, уменьшаются. Следовательно, актиновые нити во время сокращения скользят между нитями миозиновыми, а не относительно друга. 3) Сила сокращения генерируется в результате активного движения нитей одного типа вдоль прилегающих нитей другого типа. Причем каждая актиновая нить скользит в пространстве между тремя миозиновыми.

Механизм мышечного сокращения на упрощённой схеме
 и сокращением мышечных")
Электромеханическое сопряжение • Совокупность явлений, обусловливающих связь между возбуждением (потенциалом действия) и сокращением мышечных волокон
. 2. Распространение ПКП в околосинаптическую")
Этапы электромеханического сопряжения 1. Образование потенциала концевой пластинки (ПКП). 2. Распространение ПКП в околосинаптическую область. 3. Генерация потенциала действия мышечного волокна в околосинаптической области. 4. Распространение потенциала действия по поверхностной мембране мышечного волокна 5. Распространение потенциала действия по мембранам поперечных трубочек.
 6. Реакция рецепторов на изменение мембранного потенциала. 7. Открытие каналов")
Этапы электромеханического сопряжения (продолжение) 6. Реакция рецепторов на изменение мембранного потенциала. 7. Открытие каналов Ca 2+ и выход его из саркоплазматического ретикулюма (цистерн) в саркоплазму (цитозоль). 8. Диффузия Ca 2+ в межфибриллярное пространство. 9. Связывание Ca 2+ с тропонином, расположенным на тонком, актиновом миофиламенте. 10. Конформационные изменения тропонина при связывании Са 2+ «заталкивание» тропомиозина в канавки актиновых нитей 11. Начинается цикл миозиновых мостиков

Электромеханическое сопряжение
 соединяется")
Цикл миозиновых головок 1. Головка миозина, несущая продукты гидролиза АТФ (АДФ + фосфат) соединяется с миозинсвязывающим участкам актиновой нити. 2. АДФ и фосфат покидают миозиновую головку. Головка миозина изменяет конформацию за счёт шарнирного участка в области шейки миозина. Происходит гребковое движение, продвигающее актиновую нить к центру саркомера. 3. Головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. 4. Гидролиз АТФ восстанавливает конформацию молекулы миозина, и она оказывается готовой вступить в новый цикл.

Цикл миозиновых головок

Различают два режима мышечных сокращений • Изотоническое сокращение - это сокращение, при котором тонус мышц не меняется, однако длина мышечных волокон уменьшается, наблюдается при свободном сгибании конечности. • Изометрическое сокращение - это сокращение, когда мышечный тонус увеличивается, а длина мышцы не изменяется (имеет место напряжение мышцы), наблюдается, когда человек пытается поднять непосильный груз.

Одиночное и тетаническое сокращения

• Тетанус возникает вследствие суммации одиночных мышечных сокращений. • Если раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, но не успела еще полностью расслабиться, то возникает зубчатый тип сокращения (зубчатый тетанус). • Если раздражающие импульсы сближены настолько, что каждый последующий приходится на время, когда мышца еще не успела перейти к расслаблению от предыдущего раздражения, то есть происходит на высоте ее сокращения, то возникает длительное непрерывное сокращение, получившее название гладкого тетануса.

Формирование тетануса в зависимости от частоты раздражения

• • • Физиологические особенности гладких мышц. Нет поперечной исчерченности. Менее возбудимы, распространяется с небольшой скоростью – 2 -15 см/с. Возбуждение может передаваться с одного волокна на другое. Сокращение происходит более медленно и длительно. Рефрактерный период более продолжителен. Большая пластичность, т. е. способность сохранять приданную растяжением длину без изменения напряжения. Способность к автоматической деятельности. Адекватным раздражителем является их быстрое и сильное растяжение. Высокая чувствительность к некоторым биологически активным веществам (ацетилхолин, адреналин, норадреналин, серотонин). Иннервируются симпатическими и парасимпатическими вегетативными нервами, которые оказывают противоположное влияние на их функциональное состояние.

Основные свойства сердечной мышцы. • Возбудимость сердечной мышцы. Менее возбудима, чем скелетная. • Проводимость. Возбуждение по волокнам сердечной мышцы проводится с меньшей скоростью, чем по волокнам скелетной мышцы. • Сократимость. Реакция сердечной мышцы не зависит от силы наносимых раздражений. Сердечная мышца максимально сокращается и на пороговое и на более сильное по величине раздражение. • Рефрактерный период. Имеет значительно выраженный и удлиненный рефрактерный период. Он характеризуется резким снижением возбудимости ткани в период ее активности. • Автоматизм сердца. Вне организма при определенных условиях сердце способно сокращаться и расслабляться, сохраняя правильный ритм.
")
Морфо‑функциональные различия типов мышечной ткани Вид сократительного аппарата: • Скелетная мышца - Миофибриллы (длинные) • Сердечная мышца - Миофибриллы (короткие) • Гладкая мышца – Миофиламенты
: • Сердечная мышца -")
Морфо‑функциональные различия типов мышечной ткани Энергетическое обеспечение сокращения (содержание митохондрий): • Сердечная мышца - Максимальное • Скелетная мышца - Высокое (выше, чем у гладкой, ниже, чем у сердечной) • Гладкая мышца - Низкое

Морфо‑функциональные различия типов мышечной ткани Источник иннервации: • Скелетная мышца - Соматическая нервная система, эфферентная (мотонейроны спинного мозга или ствола головного мозга) • Сердечная и гладкая мышца Автономная нервная система (симпатический, парасимпатический и метасимпатический отделы)

Морфо‑функциональные различия типов мышечной ткани Характер иннервации: • Скелетная мышца - Каждое нервное волокно снабжено нервным окончанием • Сердечная и гладкая мышца Небольшая часть клеток снабжена нервными окончанием
 раздражитель: Скелетная мышца - Медиатор ацетилхолин Сердечная")
Морфо‑функциональные различия типов мышечной ткани Физиологический (адекватный) раздражитель: Скелетная мышца - Медиатор ацетилхолин Сердечная мышца - Медленная диастолическая деполяризация клетки пейсмекера; ПД «соседнего» волокна нексус Гладкая мышца: • Медиатор ацетилхолин или норадреналин (ВПСП); • Гуморальные факторы (адреналин, ангиотензин, метаболиты); • Механическое растяжение.

Морфо‑функциональные различия типов мышечной ткани Место возникновения возбуждения: • Скелетная мышца - Околосинаптическая мембрана • Сердечная мышца - Область, расположенная около нексуса • Гладкая мышца - Вся мембрана

Морфо‑функциональные различия типов мышечной ткани Возможность передачи возбуждения от клетки к клетке • Скелетная мышца - Нет • Сердечная и гладкая мышца - Есть
 • Скелетная мышца")
Морфо‑функциональные различия типов мышечной ткани Способность к спонтанной генерации импульсов (автоматии) • Скелетная мышца - Нет • Сердечная мышца – Есть. Выраженность этой способности у атипичных и рабочих сердечных миоцитов существенно разнится. • Гладкая мышца - Есть

Морфо‑функциональные различия типов мышечной ткани Характер сокращения • Скелетная мышца - Тетанический произвольный • Сердечная мышца - Ритмический непроизвольный • Гладкая мышца – Тонический непроизвольный

Морфо‑функциональные различия типов мышечной ткани Источники Ca++, активирующего мышечное сокращение • Скелетная мышца – саркоплазматический ретикулум • Сердечная мышца - саркоплазматический ретикулум = внеклеточная жидкость • Гладкая мышца – внеклеточная жидкость > саркоплазматический ретикулум

Морфо‑функциональные различия типов мышечной ткани Рецепторный белок для Ca++, активирующего мышечное сокращение • Скелетная мышца, Сердечная мышца - тропонин • Гладкая мышца – кальмодулин

Морфо‑функциональные различия типов мышечной ткани Несократительные функции: • Скелетная мышца - Терморегуляция, углеводный обмен • Сердечная мышца - Синтез атриопептидов • Гладкая мышца – Продукция эластических волокон
Тема_5 мышцы.ppt