Презентация стебель 2.ppt
- Количество слайдов: 44



Микроскопическое строение стебля 1. 2. 3. 4. Общие закономерности строение стебля. Микроскопическое строение стебля двудольных травянистых растений. Микроскопическое строение стебля однодольных растений. Микроскопическое строение стебля покрытосеменных древесных растений. 5. Микроскопическое строение стебля голосеменных древесных растений. 6. Стелярная теория.

Стебель – осевой радиально-симметричный орган растения, осуществляющий связь между корнями и листьями. Основные функции: транспорт, запас веществ и опора. Характерные черты строения стебля: • • • Стебель образуется в конусе нарастания побега и защищен почечными чешуями и листьями. На стебле образуются листья (примордии), почки, цветки и плоды. Стебель способен к ветвлению. Стебель обладает положительным фототропизмом. Стебель морфологически разнообразен (травянистый – древесный, прямостоячий – стелющийся, жесткий – гибкий, однолетний – многолетний, тонкий – толстый, удлиненный – укороченный и т. д. ). Стебель имеет разнообразное микроскопическое строение (первичное и вторичное, сплошное – пучковое – переходное, индивидуальные – синтетические листовые следы).

Строение верхушки побега в продольном разрезе. Обозначения: з. п. п — зачаток пазушной почки, к. н — конус нарастания, л — листья, л. б — листовой бугорок, пкм — прокамбий, п. п — пазушная почка

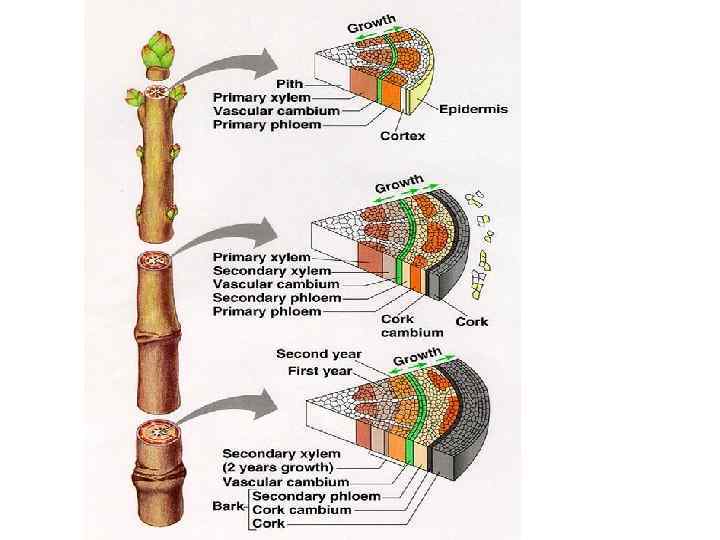
Первичное строение стебля обусловлено васкулярной меристемой – прокамбием, который образует закрытые коллатеральные пучки, содержащие ограниченное количество проводящих тканей, поэтому стебли не способны утолщаться (однодольные растения). Вторичное строение стебля связано с камбием, который вычленяется из прокамбия, в результате его деятельности образуются вторичные открытые коллатеральные и биколлатеральные пучки, количество проводящих тканей в которых ограничивается только внешними факторами, поэтому стебли способны к утолщению (голо- и покрытосеменные двудольные растения).

Стебель древесного растения первого года жизни

Распределение меристем в растении: 1 – апикальная меристема; 2 – интеркалярная меристема; 3 – прокамбий; 4 – незрелые элементы флоэмы; 5 – зрелые элементы флоэмы; 6 – незрелые элементы ксилемы; 7 - зрелые элементы ксилемы; 8 – камбий.

Стебель древесного растения второго года жизни

Стебель древесного растения на третьем году жизни

Типы строения стеблей у двудольных и однодольных растений зависят от количества и характера расположения проводящих пучков. У двудольных синтетические листовые следы – это проводящие пучки, вошедшие из листа в стебель. Листовые следы внедряются в стебель через листовые прорывы (лакуны, щели) – участки паренхимной ткани. Из листа входит несколько листовых следов (их число можно подсчитать на рубце). Проводящие пучки листа сливаются с проводящей системой стебля, которая располагаются на поперечном срезе стебля в один круг, или сплошным слоем, окружая сердцевину. У однодольных индивидуальные листовые следы – многочисленные проводящие листовые пучки, вошедшие в стебель и не слившиеся, а лишь в узлах переплетаются между собой; каждый из листовых пучков образует проводящую систему стебля. Проводящие пучки располагаются на поперечном срезе стебля диффузно.

Трехмерная реконструкция васкулатуры стебля ломоноса и ивы с трехпучковыми листовыми следами и трехлакунными узлами (А, Б) ); многопучкового трехлакунного у капусты (В); многопучкового многолакунного у щавеля (Г): 1 — пучки листового следа; 2 — лакуна.

Атактостела: А — трехмерная схема васкулатуры стебля однодольного растения; Б — поперечный срез стебля традесканции приречной: В — анастомозы проводящих пучков в узле у кукурузы; I — кортекс: 2 — проводящие пучки; 3 — стела
, прокамбий располагается непрерывным кольцом. Б – пучковое,")
Типы строения стел А – непучковое (сплошное), прокамбий располагается непрерывным кольцом. Б – пучковое, прокамбий закладывается в виде прерывистого кольца. В – переходное, в паренхиме сердцевинных лучей закладывается дополнительный камбий. Г – сплошное, при котором пучки разрастаются в тангентальном направлении.


Стебель подсолнечника
 выделяют пучковое, непучковое")
Типы строения стебля У двудольных в зависимости от расположения прокамбия (камбия) выделяют пучковое, непучковое и переходное строение стебля. • Пучковое – прокамбий закладывается в виде прерывистого кольца, кнаружи он откладывает флоэму, внутрь ксилему, образуя т. о. коллатеральные и биколлатеральные пучки, которые располагаются в один круг, чередуясь с широкими сердцевинными лучами. • Непучковое (сплошное) – прокамбий располагается непрерывным кольцом и равномерно откладывает сплошные слои кнаружи флоэмы, в ко внутри – ксилемы, сердцевинные лучи узкие, малозаметные. • Переходное – в паренхиме сердцевинных лучей закладывается дополнительный камбий, который образует вторичные коллатеральные пучки, они разрастаются и сливаются с первичными, образуются сплошные слои проводящей ткани.
 Травянистый стебель существует")
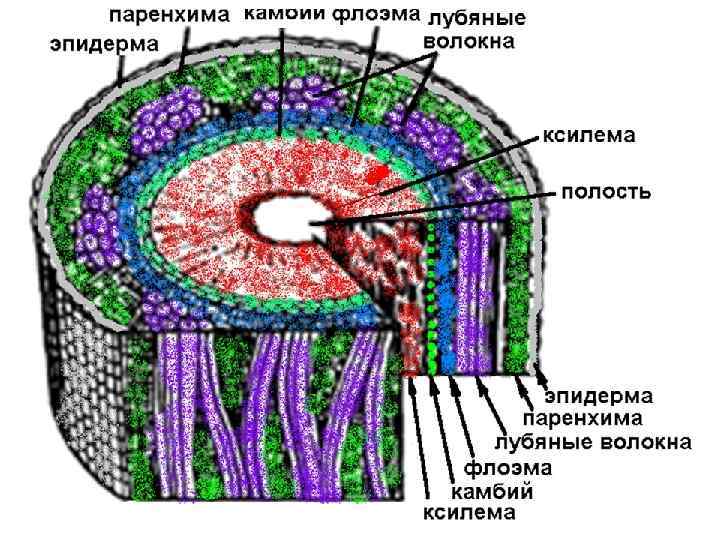
Микроскопическое строение стебля двудольного травянистого растения пучкового типа (кирказона обыкн. ) Травянистый стебель существует один вегетационный период; в анатомическом строении стебля четко выделяются первичная кора и центральный (проводящий) цилиндр; коллатеральные открытые проводящие пучки располагаются в один круг, окружая центральную сердцевину. Кора – наружный слой стебля, покрытый эпидермой, в состав которой входят колленхима, хлорофиллоносная и запасающая паренхима, а также мелкоклеточная эндодерма (крахмалоносное влагалище), граничащая с центральным цилиндром. Центральный цилиндр включает перицикл, коллатеральные открытые проводящие пучки, чередующиеся с сердцевинными лучами и центральную сердцевину. Перицикл участвует в образовании наружной перициклической толстостенной, плотно расположенной склеренхимы и крупноклеточной тонкостенной паренхимы, примыкающей к флоэмной части пучка. Эти ткани выполняют арматурную функцию по отношению к флоэме.

Коллатеральные пучки в своем составе имеют первичные проводящие ткани, образованные прокамбием и располагающиеся на полюсах пучков, а также вторичные, образованные камбием, и примыкающие непосредственно к камбию. Флоэма располагается с наружной стороны от камбия, она представлена ситовидными трубками с клетками-спутницами, лубяными волокнами и лубяной паренхимой, а ксилема – с внутренней, в ее составе сосуды, трахеиды, либриформ и древесная паренхима. В центре стебля расположена крупноклеточная паренхима сердцевины, выполняющая запасающую функцию.

Стебель кирказона
 Проводящие ткани образованы только прокамбием; в микроскопическом")
Микроскопическое строение стебля однодольных растений (кукуруза посевная) Проводящие ткани образованы только прокамбием; в микроскопическом строении стебля нет четкого деления на первичную кору и центральный цилиндр; сердцевинные лучи не выражены, а многочисленные закрытые коллатеральные пучки расположены на поперечном срезе диффузно. Под эпидермой располагается непрерывный слой склеренхимы. Всю внутреннюю часть занимает основная паренхима, в которую погружены многочисленные закрытые коллатеральные проводящие пучки. По периферии в мелкоклеточной паренхиме находятся мелкие пучки, а ближе к центру в крупноклеточной – более крупные. Проводящие пучки окружены обкладкой склеренхимных клеток. В проводящем пучке ситовидные трубки и сопровождающие клетки расположены в виде сеточки, лубяная паренхима отсутствует. Ксилема полуобъемлет флоэму, содержит два крупных и несколько более мелких сосудов, а также воздушную полость. У ржи образуется полый стебель типа соломина в результате разрушения клеток сердцевины. Утолщение стебля некоторых однодольных (пальмы, драцены, юкки) происходит в результате образования кольца утолщения в коре, состоящего из паренхимы и закрытых концентрических амфивазальных пучков, а паренхима постепенно одревесневает.


Поперечный срез стебля кукурузы, показывающий паренхимные ткани и разбросанные проводящие пучки. Большинство клеток пучка составляют сосуды [Magnified Approximately 250 X. ]

 Стебель многолетний, способен к ежегодному вторичному")
Микроскопическое строение стебля древесных покрытосеменных растений (липа сердцелистная) Стебель многолетний, способен к ежегодному вторичному утолщению, благодаря периодически возобновляемой деятельности камбия. Стебель имеет непучковое (сплошное) строение, в котором выделяют кору и древесину, граница между которыми проходит по камбию, а также центральную сердцевину. Покровная ткань у молодых растений эпидерма, затем формируется пробка, на смену которой приходит корка (ритидом). Кора по составу неоднородна, она включает первичную кору и вторичную кору. Первичная кора включает пластинчатую колленхиму, основную паренхиму и слабо выраженную эндодерму.


Поперечное сечение стебля липы, демонстрирующее большую сердцевину, многочисленные лучи и три различающихся кольца древесины
, разделена сердцевинными лучами на участки, имеющие форму трапеций, расширяющихся в")
Вторичная кора (флоэма, луб), разделена сердцевинными лучами на участки, имеющие форму трапеций, расширяющихся в сторону камбия. Каждый такой участок включает толстостенную перициклическую склеренхиму, чередующуюся с паренхимой, которые непосредственно примыкают к собственно флоэме. Флоэма представлена перемежающимися слоями, состоящими из ситовидных трубок с клетками-спутниками и паренхимы (мягкий луб) и толстостенных лубяных волокон (твердый луб). Функция луба нисходящий транспорт органических веществ. Камбий – латеральная меристема, расположенная между лубом и древесиной. Камбий образует материнские (инициальные) клетки флоэмы и материнские клетки ксилемы, которые участвуют в образовании флоэмы и ксилемы. Веретеновидные по форме инициали образуют прозенхимные элементы (трахеиды, трахеи, ситовидные трубки, волокна др. ), лучевые – паренхимные элементы. Ксилемных элементов откладывается в десятки раз больше, чем лубяных, поэтому камбий и кора отодвигаются к периферии. Ежегодно деятельность камбия в конце вегетационного периода приостанавливается, а в начале – возобновляется.

Строение стебля липы
 располагается внутрь от камбия и занимает до 90% объема ствола. Она представлена")
Древесина (ксилема) располагается внутрь от камбия и занимает до 90% объема ствола. Она представлена годичными кольцами (приростами) т. е. слоями древесины, образованными за один вегетационный период. Каждое кольцо включает внутренний слой – это ранняя (весенняя) ксилема и наружный слой – поздняя (летняя) ксилема. Ранняя древесина содержит широкопросветные проводящие элементы и древесную паренхиму, а поздняя – образована толстостенными узкопросветными трахеальными элементами и механическими волокнами. Различие между ранней и поздней древесиной хорошо заметно на поперечном срезе и позволяет определить возраст дерева. Функция древесины восходящий транспорт минеральных веществ. Сердцевина хорошо развита, к первичной древесине непосредственно примыкает ее перимедуллярная зона, составленная мелкими клетками, богатыми крахмалом, а в центре находятся более крупные клетки сердцевины. Функция сердцевины запас и выделение веществ. Сердцевинные (лубо-древесинные) лучи пересекают стебель в радиальном направлении и участвуют в горизонтальном транспорте веществ.



Возрастные изменения древесины и коры С годами древесина дифференцируется на ядро и заболонь. Внутренняя более темная ядровая часть древесины отмирает, трахеиды перекрываются торусом, сосуды заполняются тиллами, накапливаются пигменты, смолы, танины, ткани сильно лигнифицируются. Ядро выполняет механическую и защитную функцию. Наружная более светлая заболонная часть, примыкающая к камбию, осуществляет транспорт веществ. Увеличение окружности стебля вызывает разрастание паренхимных клеток коры (дилатация) в тангентальном направлении. Дилатация бывает лучевой (липа), диффузной (береза) или диффузно-лучевой (большинство растений). На стволах и старых ветвях образуется корка или ритидом, а первичная кора со временем исчезает, входя в состав отслаивающихся участков ритидома, поэтому на старых стволах кора включает только вторичную кору и ритидом.


Определите по спилу возраст ствола сосны.
 Покрывает стебель пробка, составленная из чередующихся слоев")
Анатомическое строение стебля голосеменных растений (сосна обыкновенная) Покрывает стебель пробка, составленная из чередующихся слоев тонкостенных опробковевших и толстостенных одревесневших клеток, что способствует ее легкому отслаиванию. Первичная кора включает гиподерму (аналогична колленхиме) и паренхиму. Луб (флоэма) состоит из ситовидных клеток, и лубяной паренхимы, лубяные волокна и клетки спутницы отсутствуют. Древесина (ксилема) располагается концентрическими слоями, которые представляют собой годичный прирост проводящих тканей. Во внутренней светлой, образованной за весенний период, части каждого кольца закладываются широкопросветные трахеиды, выполняющие водопроводящую функцию. В наружной, более темной части, образованной за летнее-осенний период, возникают толстостенные трахеиды, играющие механическую роль. Либриформ и древесная паренхима отсутствуют. В стебле располагаются смоляные каналы, выстланные эпителиальными клетками и заполненные живицей, играющие защитную функцию. В центре стебля находится слаборазвитая сердцевина. В радиальном направлении стебель пересекают сердцевинные (лубодревесинные) лучи.


3 -х летний прирост древесины стебля сосны.

Стелярная теория – это учение о строении и эволюции проводящих тканей стебля у различных отделов высших растений. Эволюция стелы сопровождалась увеличением доли паренхимы в проводящих тканях и в сердцевине, в также разделением одного цельного осевого тяжа проводящих тканей стебля на отдельные более мелкие тяжи в составе которых все большую часть занимала проводящая система листьев. Эволюция стелы включает три направления: протостела, сифоностела и полистелия. Протостела шла по пути увеличения поверхности соприкосновения между ксилемой и флоэмой, встречается у микрофилльных растений (плауновидных). Протостела имеет три модификации: гаплостела, актиностела и плектостела. Гаплостела представляет собой центральный округлый тяж ксилемы, который снаружи кольцом окружает флоэма; актиностела – центральная лопастная ксилема, окруженная флоэмой и плектостела – центральная ксилема раздроблена на отдельные тяжи вокруг которых находится флоэма.
, образованных при внедрении")
Сифоностела характеризуется возникновением в стебле сердцевины, и листовых прорывов (лакун), образованных при внедрении проводящих тканей листьев в проводящую систему стебля и заполненных паренхимой; встречается у папоротниковидных. Выделяют три модификации сифоностелы: эктофлойная сифоностела – в центре располагается сердцевина, снаружи от которой находятся слои флоэмы и ксилемы; амфифлойная сифоностела – отличается от эктофлойной наличием не только наружной, но и внутренней флоэмы, которая граничит с сердцевиной и диктиостела – состоит из отдельных концентрических амфикрибральных пучков (меристел), расположенных по кругу.
 внутри которых и между которыми")
Полистелия связана с появлением множества отдельных проводящих пучков (стел) внутри которых и между которыми находится паренхима, а также постепенной редукцией осевой проводящей системы стебля и заменой ее проводящей системой листьев. Эвстела представлена открытыми коллатеральными и биколлатеральными пучками, располагающимися вокруг сердцевины в один ряд или сплошными слоями флоэмы, камбия и ксилемы (голосеменные и двудольные покрытосеменные растения). Атактостела – многочисленные закрытые коллатеральные пучки листового происхождения располагаются диффузно на поперечном срезе стебля (однодольные растения). Артростела представлена закрытыми коллатеральными пучками, расположенными вокруг центральной полости, в узлах каждый из пучков расщепляется на три (хвощи).
. 1 – протостела")
Схема эволюции стелы (ксилема заштрихована продольными штрихами, а флоэма – точками). 1 – протостела или гаплостела; 2 – актиностела; 3 – плектостела; 4, 5 – эктофлойная и амфифлойная сифоностела; 6 – диктиостела; 7 – артростела; 8 – эустела; 9 – атактостела.

Типы стелы и их эволюция: 1 – протостела; 2 – актиностела; 3 – стела корня; 4 – сифоностела; 5 – диктиостела; 6 – эустела; 7 – атактостела. Сопоставлены поперечные разрезы и трёхмерные изображения. Ксилема – чёрная.

Презентация стебель 2.ppt