Взаимодействия.ppt
- Количество слайдов: 63


 взаимодействия Итог взаимодействия: + - 0")
Межвидовые (биотические) взаимодействия Итог взаимодействия: + - 0

Типы взаимодействий + + Мутуализм + - Хищничество - - Конкуренция + 0 Комменсализм (нахлебничество) - 0 Аменсализм (ассиметричная конкуренция) 00 Нейтрализм

Мутуализм — любые взаимовыгодные отношения между видами но их нужно доказать. . .

Даже если они очевидны. . .

Протокооперация Медоед Медоуказчик

Актиния и амфиприон

Симбиоз — частный случай мутуализма

Термиты способны питаться чистой целлюлозой Жгутиковые из кишечника термитов Hypermastigina

Микориза увеличивает площадь питания проростка лиственницы в 100 000 раз.


Муравьи - «скотоводы»

Кальмар Euprymna scolopes использует светящихся симбиотических бактерий Vibrio fischeri для отпугивания хищников

Эволюционное возникновение мутуализма Несбалансированные фитопатогены Сбалансированные фитопатогены Микориза Фитофаги Локализация нектаром Опылители Фитопатогенные бактерии Образование галлов Клубеньковые азотфиксаторы
 В эукариотическом геноме смесь")
Симбиогенетическое возникновение эукариот (надцарство Эукариот, отдел Лишайники и пр. ) В эукариотическом геноме смесь генов архейного и бактериального происхождения, которые объединились на ранних этапах становления эукариотической клетки

Конкуренция Взаимодействие между видами, использующими один и тот же ограниченный ресурс Классификация конкурентных отношений: 1. Внутривидовая и межвидовая 2. Прямая (интерференция) и косвенная (эксплуатация) Основное различие между внутривидовой и межвидовой конкуренцией Можно ли вымереть в результате внутривидовой конкуренции? Нет (кроме, возможно, каких-то экзотических случаев) Можно ли вымереть в результате межвидовой конкуренции? Да

Классическая модель конкуренции Лотки-Вольтерры: Логика уравнений Экспоненциальный рост численности первого вида – экспоненциальное уравнение описывает рост при полном отсутствии конкуренции (популяция ничем не лимитирована) Логистический рост численности первого вида – логистическое уравнение описывает рост при наличии внутривидовой конкуренции. Это происходит в силу наличия «замедляющего» члена (K-N)/K; с ростом N величина (K-N)/K уменьшается и скорость роста замедляется. Рост численности первого вида при наличии второго вида – это уравнение описывает рост как с учетом внутривидовой конкуренции, так и с учетом межвидовой конкуренции Коэффициент конкуренции α 12 показывает, во сколько раз одна особь 2 -го вида потребляет больше ресурсов (или занимает больше пространства), чем одна особь 1 -го вида. Т. е. это коэффициент воздействия 2 -го вида на 1 -й вид.
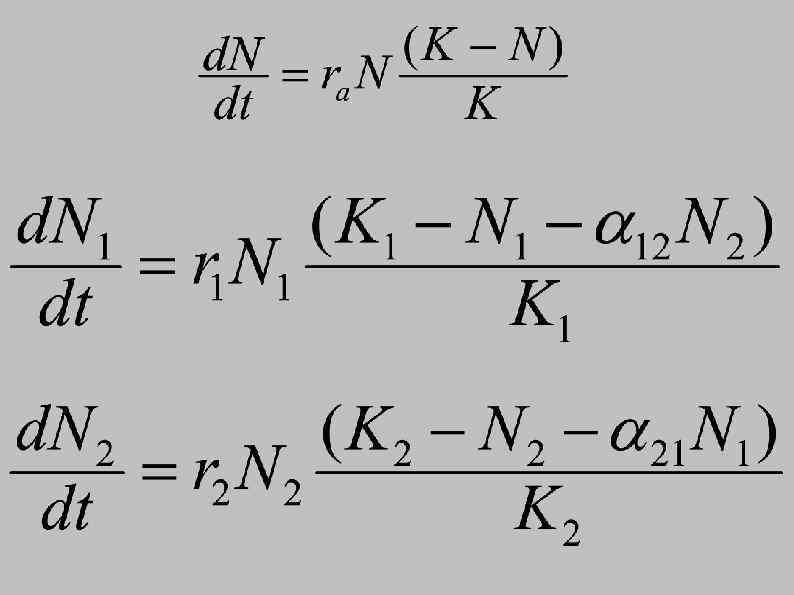
 в современной")
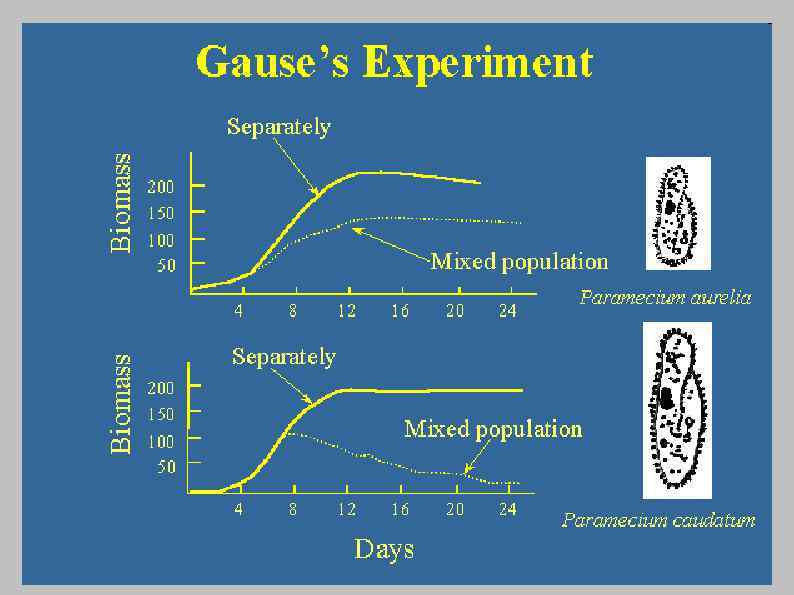
Принцип конкурентного исключения ( «закон Гаузе» , «принцип Вольтерры – Гаузе» ) в современной трактовке: Число видов, неограниченно долго сосуществующих в постоянных условиях гомогенного местообитания с постоянными численностями, не может превышать числа плотностно-зависимых факторов, лимитирующих развитие их популяций А. М. Гиляров

Кто победит? Tribolium confusum Tribolium castaneum

Исход конкуренции между мучными хрущаками в зависимости от температуры и влажности среды
 и T. weissflogii (Tw) при совместном росте на")
Динамика биомассы водорослей P. delicatissima (Pd) и T. weissflogii (Tw) при совместном росте на средах с мочевиной (а, в) и нитратами (б, г). Верхний ряд (а, б) - низкая освещенность, нижний ряд (в, г) - высокая освещенность. Л. В. Ильяш, Е. В. Запара. 2006. Конкуренция двух морских диатомовых водорослей за азот мочевины и нитратов при трех уровнях освещенности // Журнал общей биологии, т. 67

Два способа достичь сосуществования: 1. Виды расходятся по разным экологическим нишам: соответственно, конкуренция между ними отсутствует или же резко ослаблена.
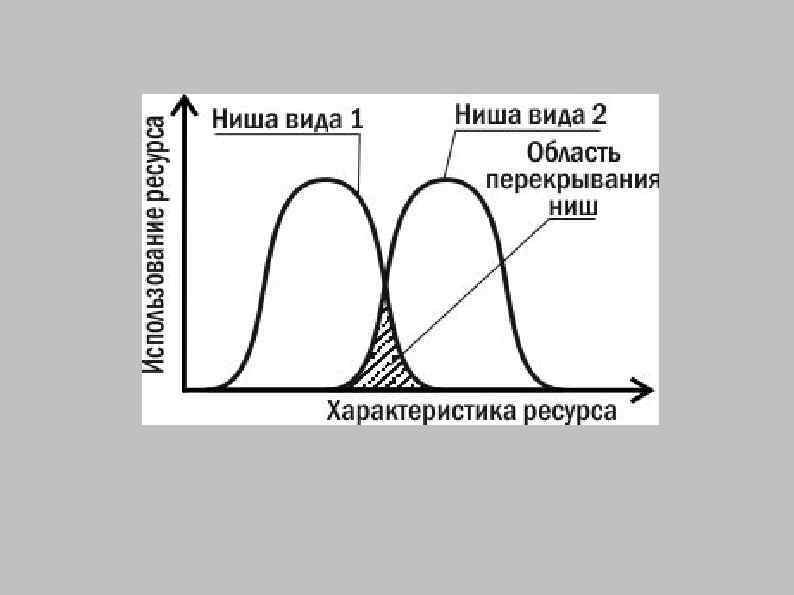


Два способа достичь сосуществования: 2. Виды занимают одну экологическую нишу, конкурируют за общие ресурсы, но вытеснение одного вида другим откладывается на долгое время, прерывается извне или же не допускается путем интерференции (ингибирования одних видов другими).

Способы сосуществования видов, занимающих одну и ту же экологическую нишу: 1. Ограничение разными ресурсами. 2. Смена конкурентного преимущества по мере расходования лимитирующего ресурса или же, наоборот, дополнительного его поступления в среду. 3. Прерывание процесса вытеснения нарушениями извне за счет абиотических (пожары, осыпи и т. п. ) или биотических факторов (выедание, вытаптывание). 4. Интерференция – ингибирование одних видов другими за счет специально выделяемых веществ. 5. Конвергенция разных видов, направленная на возрастание экологического сходства (сосуществование за счет «отложенного вытесенения» ). 6. Мутуалистические взаимоотношения между видами, иногда опосредованные (например, через микоризу).
 (resource-based competition theory) Успех вида в конкуренции определяется")
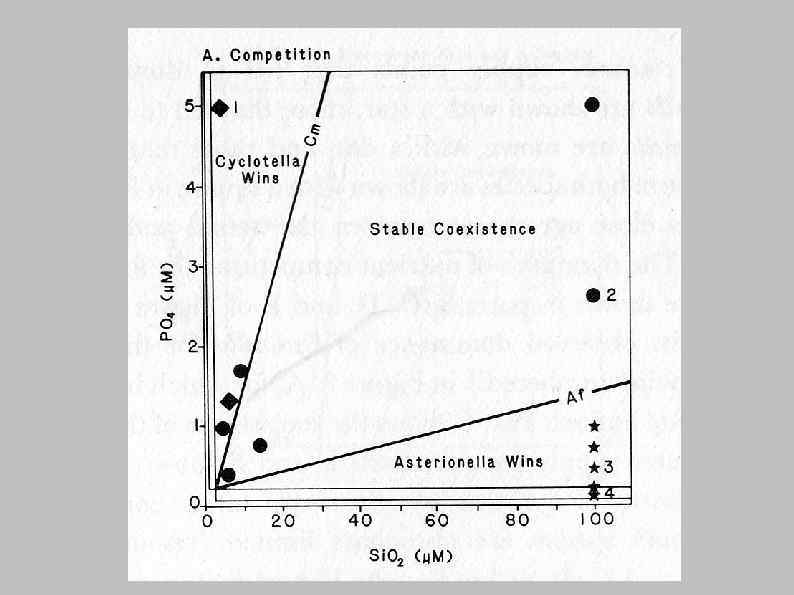
Ресурсная теория конкуренции Тильмана (David Tilman) (resource-based competition theory) Успех вида в конкуренции определяется пороговой концентрацией ресурса для этого вида, а возможность сосуществования с др. видами – скоростью потребления лимитирующего ресурса

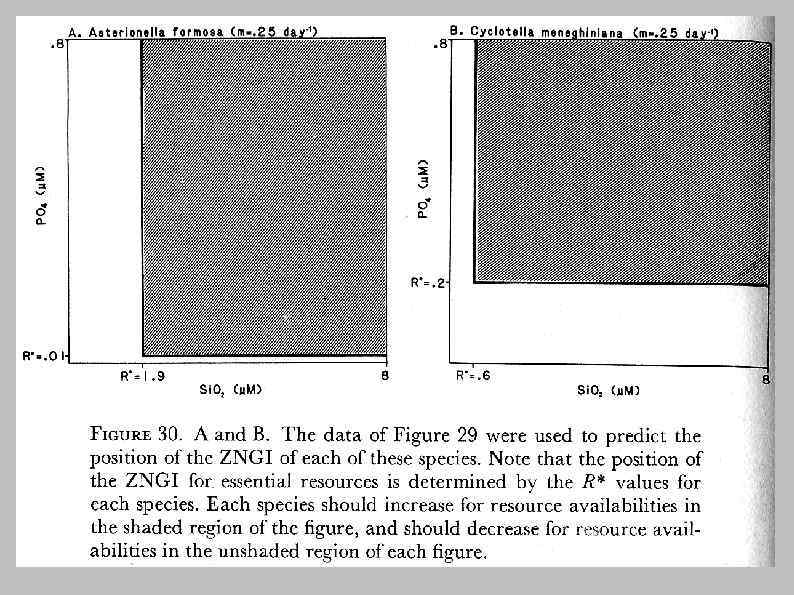
Лимитирование фосфором Лимитирование кремнием Asterionella formosa и Cyclotella meneghiniana в изолированных культурах

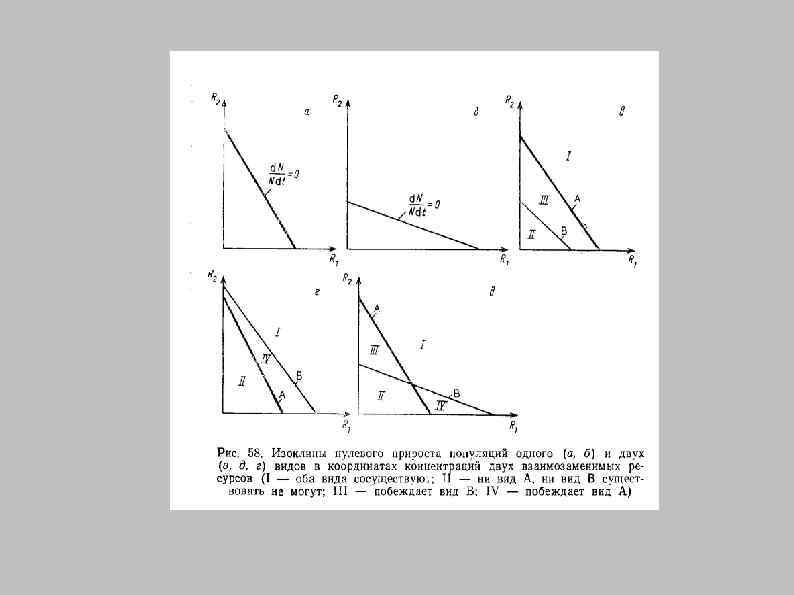
Два ресурса: Изоклины нулевого роста Незаменимые ресурсы R 2 r=0 Полностью взаимозаменяемые ресурсы R 2 R*2 r>0 r= 0 Концентрация 2 -го ресурса Изоклины нулевого роста численности r = 0 в пространстве двух ресурсов r=0 R*2 r<0 R*1 R 1 Концентрация 1 -го ресурса

Побеждает A Побеждает B Выживает A Выживает B R A R 1 B R 2 A Изоклины пересекаются Изоклины не пересекаются R 2 Конкуренция двух видов (А и В) за два ресурса (R 1 и R 2) R 2 B A B R 1
 обычный (sensitive) 2) убийца (killer) 3) устойчивый")
Ингибирование продуктами обмена: Три клона бактерий: 1) обычный (sensitive) 2) убийца (killer) 3) устойчивый (resistant) завоевывают поверхность питательной среды (один ресурс)
 обычного (S) устойчивому (R) Обычный (S) устойчивого (R)")
Штамм: Выигрывает у: Проигрывает: Киллер (K) обычного (S) устойчивому (R) Обычный (S) устойчивого (R) киллеру (K) Устойчивый (R) киллера (K) обычному (S)

Мутуализм и конкуренция Зеленые кружки — виды (P — растения, A — животные, например насекомые-опылители). Сплошные синие стрелки — мутуалистические связи, сплошные красные стрелки — конкуренция, пунктирные стрелки — косвенные эффекты мутуализма. Слева (a) — структура сообщества является «вложенной» : видыспециалисты, имеющие только по одному партнеру (растение P 1 и животное A 2), сотрудничают с видамигенералистами, имеющими по два партнера. В этом случае мутуализм ведет к ослаблению конкуренции (например, вид А 1 является не только конкурентом вида A 2, но и «другом его друга» — растения P 2). Справа (b) — структура не является «вложенной» , поскольку виды-специалисты A 2 и P 3 сотрудничают не с генералистами, а друг с другом. В этом случае мутуализм приводит к обострению конкуренции. Например, вид A 1 не только конкурирует с видом А 2 напрямую, но еще и косвенно вредит ему, помогая растениям P 1 и P 2 — конкурентам растения P 3. George Sugihara, Hao Ye. Complex systems: Cooperative network dynamics // Nature. 2009. V. 458. P. 979– 980.

Планктонный парадокс В верхнем освещенном и перемешиваемом слое водной толщи обитает множество организмов фитопланктона, ограниченных в своем развитии одними и теми же немногими ресурсами. Данная ситуация противоречит принципу конкурентного исключения Вольтерры – Гаузе.

Гипотеза нейтральности Стивена Хаббелл и его сторонники утверждают, что виды, входящие в одно сообщество и как правило конкурирующие за общие ресурсы, могут обитать вместе, не вопреки, а благодаря тому, что очень сходны по своим экологическим характеристикам. Они способны сосуществовать, если экологически идентичны, если в расчёте на одну особь ( «на душу населения» ) имеют равный шанс размножиться (оставить определенное количество жизнеспособных потомков), погибнуть, заселить освободившийся участок и даже эволюционировать. Как неоднократно подчеркивает в своих работах Хаббел, под нейтральностью следует понимать вовсе не отсутствие взаимодействий между особями разных видов, а их одинаковость.
 и степени разделения ниш (ось Х) может")
Комбинация сходства демографических характеристик видов (ось Y) и степени разделения ниш (ось Х) может обеспечивать сосуществование видов. Разделение ниш характеризуется «степенью стабилизации» . Чем более схожи между собой виды по скорости популяционного роста (значения по оси Y ближе к 1), тем меньше потребность в стабилизирующем механизме для достижения их сосуществования. Если же виды по скорости популяционного роста сильно различаются, то их сосуществование возможно только за счет различия ниш (большей стабилизации). Область сосуществования — правая верхняя часть поля графика, а область конкурентного исключения — нижняя левая. Механизму нейтральности соответствуют точки, лежащие в левой верхней части графика, а механизму разделения ниш — в правой нижней Peter B. Adler, Janneke Hille. Ris. Lambers, Jonathan M. Levine. A niche for neutrality // Ecology Letters. 2007. V. 10 (2). P. 95– 104

Экологическая ниша – «место» , занимаемое видом в сообществе • место можно понимать буквально – как место в пространстве, т. е. пространственное распределение вида (Grinnell) • место можно понимать как положение вида в цепях питания, т. е. как набор необходимых виду пищевых ресурсов (пищевых организмов, элементов минерального питания у растений и т. д. ) (Elton) • положение вида в многомерном пространстве условий и ресурсов (многомерная ниша Хатчинсона, Hutchinson) Экологическая ниша: 1. Фундаментальная — набор требований вида к условиям среды 2. Реализованная — ниша занимаемая в сообществе, результат взаимодействия с другими видами
")
Многомерная экологическая ниша (ниша Хатчинсона)

Модель ниши Хатчинсона

Реализованная экологическая ниша Положение вида, которое занимает в общей системе биоценоза в зависимости от его требований к абиотическим факторам (потенциальной ниши) и в условиях биотических ограничений (конкуренции) Chthamalus Balanus Реали. Потен- зованная циальная Океан Потенциальная и реализиванная экологические ниши для двух видов усоногих рачков в зоне прилива

Конкурентное высвобождение — расширение реализованной ниши до фундаментальной при удалении конкурента Эксперименты Ж. И. Резниковой в сообществе степных муравьев Formica cunicularia Formica pratensis
")
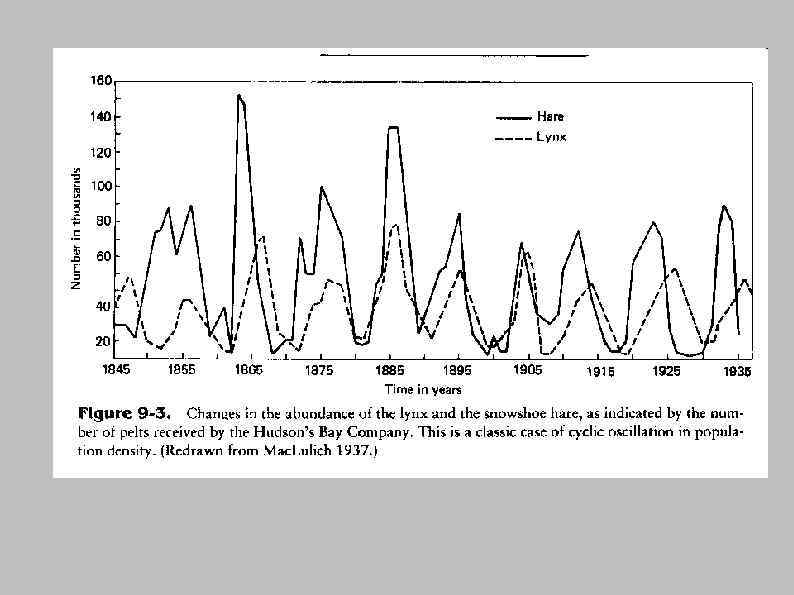
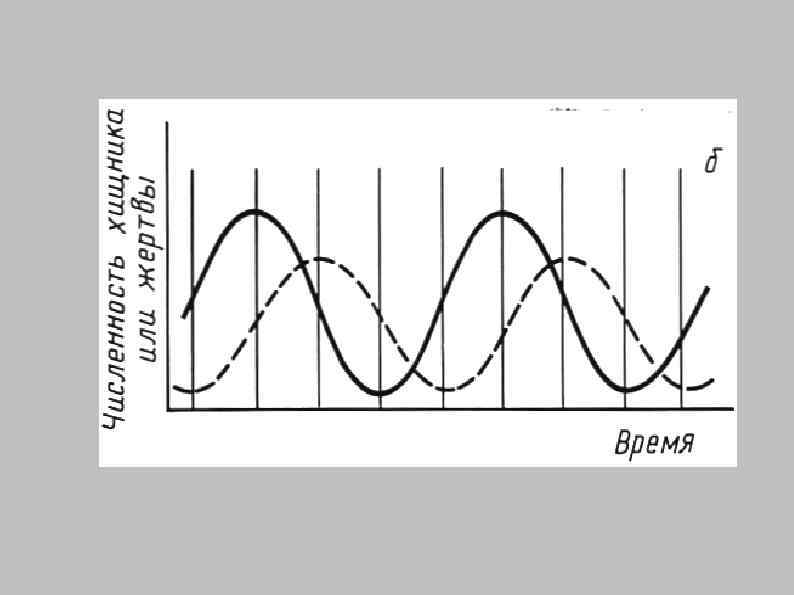
Хищник — жертва (ресурс-потребитель)
 2. Пастьба")
Классификация типов отношений ресурс потребитель 1. Истинное хищничество (1 жертва, жертва погибает) 2. Пастьба (Много жертв, жертвы не погибают) 3. Паразитизм (1 жертва, жертва не погибает) 4. Фильтрация (Много жертв, жертвы погибают) Классификация по спектру питания: 1. Монофаги 2. Олигофаги 3. Полифаги


Уравнение для жертвы N 1 – численность жертвы N 2 – численность хищника r 1 – скорость популяционного роста жертвы в отсутствие хищника p 1 – коэффициент, переводящий встречу хищника и жертвы в смертность жертв

Уравнение для хищника N 1 – численность жертвы N 2 – численность хищника p 2 – коэффициент, переводящий встречу хищника и жертвы в рождаемость хищника m 2 – смертность хищника

A. Неэффективный хищник не способен успешно эксплуатировать свою жертву до тех пор, пока плотность ее популяции не приблизится к предельной плотности насыщения. Векторы образуют спираль, закрученную внутрь, при этом колебания плотности хищника и жертвы затухают и система стремится к точке совместного устойчивого равновесия (в которой пересекаются изоклины). Б. Высокоэффективный хищник способен эксплуатировать очень немногочисленные популяции жертвы, имеющие предельно низкую плотность. На этот раз векторы образуют раскрученную спираль и амплитуда колебаний популяций непрерывно возрастает вплоть до предельного цикла, после чего часто либо только хищник, либо хищник вместе с жертвой погибают. В. Хищник, обладающий умеренной эффективностью, начинает эксплуатировать популяцию жертвы при средних значениях ее плотности. Здесь векторы образуют замкнутый круг, а колебания популяций хищника и жертвы соответствуют колебаниям нейтральной устойчивости. (По Mac. Arthur, Connell, 1966. ) I — жертва; II — хищник.

Система хищник — жертва у инфузорий: опыты Г. Ф. Гаузе Paramecium caudatum Didinium nasutum

Huffaker, C. B. 1958. Experimental studies on predation: dispersion factors and predator-prey oscillations. Hilgardia 27(14): 343 -383 Predator Typhlodromus occidentalis http: //www. goodbugs. org. au/BCAs%20 available/occidentalis. htm Prey Eotetranychus sexmaculatus http: //www. uae. gov. ae/uaeagricent/qgcc/application/insects/A 1/IA 1 -52. htm
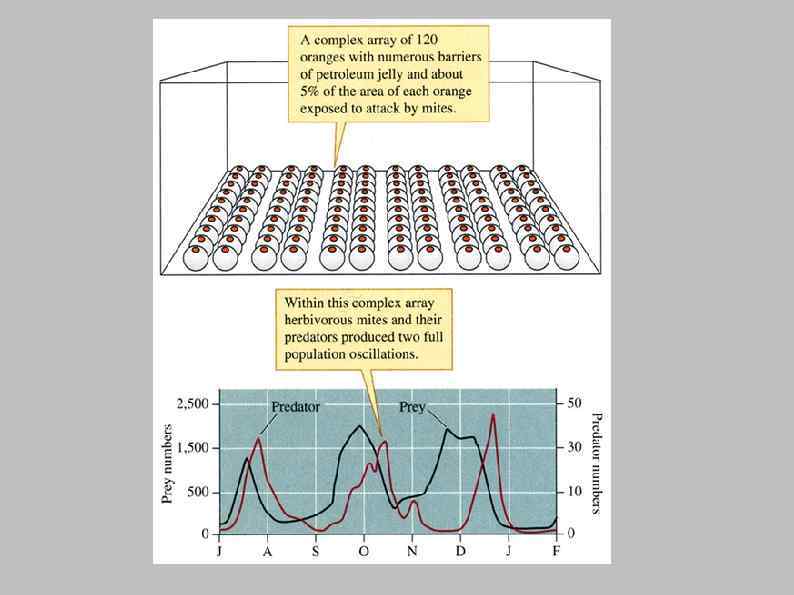

Конкуренция и хищничество Изменения размера популяций жука-хозяина и двух паразитических ос за четырехлетний период (Utida, 1957) Neocatolaccus mamezophagus Callosobruchus chinensis Heterospilus prosopidis

На контрольных участках, где присутствовал Pisaster, было зарегистрировано 15 видов беспозвоночных, а на участках, лишенных морской звезды, — только 8 видов. Pisaster ochraceus Mytilus californianus
 и численная реакция (numerical response) хищника в ответ на увеличение")
Функциональная реакция (functional response) и численная реакция (numerical response) хищника в ответ на увеличение численности жертв
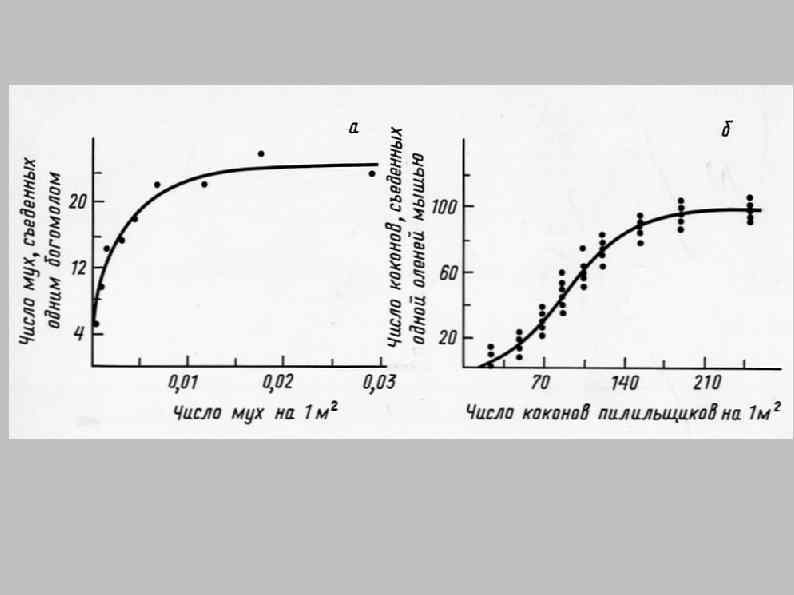

Gilg, O. et al. Functional and numerical responses of four lemming predators in high arctic Greenland. Oikos 113, 193– 216 (2006)

Теория оптимального фуражирования: Почему не все хищники генералисты? Схема, показывающая, как Холдинг, используя геометрию и исходя из анатомии передней ноги богомола, рассчитал оптимальный размер жертвы. Оптимальный диаметр жертвы D равен Т sin (β — α), где Т — расстояние А С. (Из Holling, 1964. ) Процент жертв, атакованных голодными богомолами, в зависимости от размеров жертвы. (Из Holling, 1964. )

Пищевое предпочтение — избирательное потребление какого-либо вида пищи Пищевое переключение — смена предпочитаемого вида пищи
Взаимодействия.ppt