

Межклеточные взаимодействия.ppt
- Количество слайдов: 77
 Межклеточные взаимодействия 1. Гликокаликс, внеклеточный матрикс 2. Адгезивность 3. Онкогены и онкобелки мембран 4. Сигнальные молекулы 5. Типы межклеточной сигнализации 6. Классификация первичных мессенджеров 7. Рецепторы клеточной поверхности 8. Рецепторные тирозинкиназы
Межклеточные взаимодействия 1. Гликокаликс, внеклеточный матрикс 2. Адгезивность 3. Онкогены и онкобелки мембран 4. Сигнальные молекулы 5. Типы межклеточной сигнализации 6. Классификация первичных мессенджеров 7. Рецепторы клеточной поверхности 8. Рецепторные тирозинкиназы
 Гликокаликс – поверхностный матрикс клетки • В мембранах гликозилировано около 10% всех белков и от 5 до 26% липидов • Углеводные цепи белков колеблются по составу от двухзвенных структур до разветвленных 18 -членных полисахаридов • Наиболее часто в мембранных гликопротеинах и гликолипидах встречаются галактоза, манноза, фукоза, галактозамин, глюкоза и сиаловая кислота. • Остатки сиаловой кислоты находятся обычно на концах углеводных боковых цепей, они ответственны за отрицательный заряд клеточной поверхности, характерный для всех эукариотических клеток.
Гликокаликс – поверхностный матрикс клетки • В мембранах гликозилировано около 10% всех белков и от 5 до 26% липидов • Углеводные цепи белков колеблются по составу от двухзвенных структур до разветвленных 18 -членных полисахаридов • Наиболее часто в мембранных гликопротеинах и гликолипидах встречаются галактоза, манноза, фукоза, галактозамин, глюкоза и сиаловая кислота. • Остатки сиаловой кислоты находятся обычно на концах углеводных боковых цепей, они ответственны за отрицательный заряд клеточной поверхности, характерный для всех эукариотических клеток.
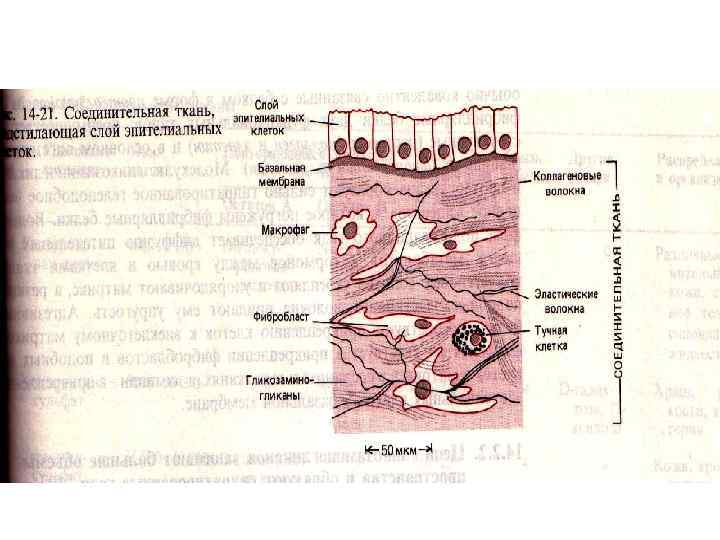
 Гликокаликс
Гликокаликс
 Внеклеточный матрикс В образовании матрикса участвуют два основных класса макромолекул: • коллагены - фибриллярные белки, образующие коллагеновые волокна; • гликозамингликаны, обычно связанные с белком в протеогликаны
Внеклеточный матрикс В образовании матрикса участвуют два основных класса макромолекул: • коллагены - фибриллярные белки, образующие коллагеновые волокна; • гликозамингликаны, обычно связанные с белком в протеогликаны
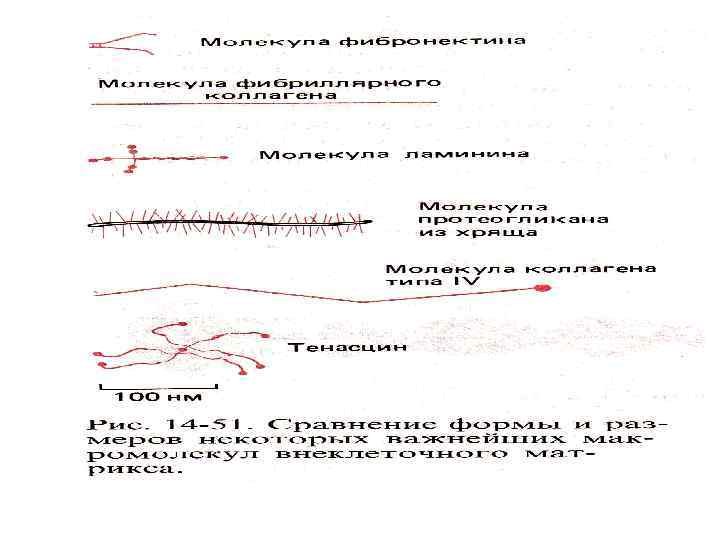
 Во внеклеточных взаимодействиях участвуют: • Фибронектин образует на поверхности многих клеток фибриллярные структуры • Ламинин – неколлагеновый гликопротеин базальных мембран
Во внеклеточных взаимодействиях участвуют: • Фибронектин образует на поверхности многих клеток фибриллярные структуры • Ламинин – неколлагеновый гликопротеин базальных мембран
 Фибронектин
Фибронектин
 Ламинин
Ламинин
 Эластин
Эластин
 Механизм опухолевой инвазии 1. взаимодействие опухолевой клетки с поверхностными ламининами, фибронектином; 2. секреция гидролитических ферментов и локальное нарушение внеклеточного матрикса; 3. движение опухолевых клеток по модифицированному протеолизом матриксу. Ферменты, участвующие в разрушении коллагенов I и IY типов, являются наиболее активными участниками процесса инвазии. Коллагеназы I и IY усиливают миграцию опухолевых клеток. Коллагеназа IY типа является маркером опухолей при лимфопролиферативных заболеваниях
Механизм опухолевой инвазии 1. взаимодействие опухолевой клетки с поверхностными ламининами, фибронектином; 2. секреция гидролитических ферментов и локальное нарушение внеклеточного матрикса; 3. движение опухолевых клеток по модифицированному протеолизом матриксу. Ферменты, участвующие в разрушении коллагенов I и IY типов, являются наиболее активными участниками процесса инвазии. Коллагеназы I и IY усиливают миграцию опухолевых клеток. Коллагеназа IY типа является маркером опухолей при лимфопролиферативных заболеваниях
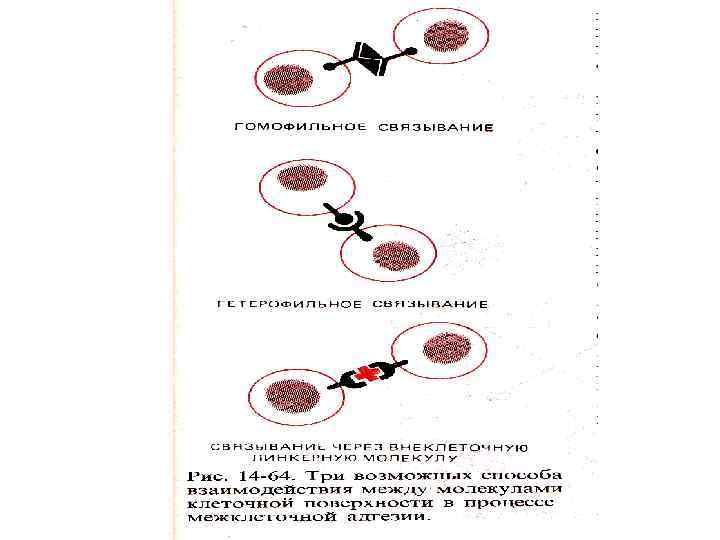
 Рецепторы клеточной поверхности • Интегрины - рецепторы клеточной поверхности для молекул межклеточного матрикса: коллагена, фибронектина, ламинина • Интегрины тромбоцитов участвуют в агрегации тромбоцитов • Лейкоцитарные белки адгезии участвуют в во взаимодействии лейкоцитов с эндотелиальными клетками сосудов при воспалении
Рецепторы клеточной поверхности • Интегрины - рецепторы клеточной поверхности для молекул межклеточного матрикса: коллагена, фибронектина, ламинина • Интегрины тромбоцитов участвуют в агрегации тромбоцитов • Лейкоцитарные белки адгезии участвуют в во взаимодействии лейкоцитов с эндотелиальными клетками сосудов при воспалении
 Интегрины
Интегрины
 Кадгерины • Е-кадгерин находится на поверхности клеток эпителиальных и эмбриональных тканей • N-кадгерин локализован на поверхности нервных клеток, хрусталика, сердца • Р-кадгерин расположен на клетках плаценты и эпидермиса Кадгерины участвуют в начальных стадиях адгезии, обеспечивают структурную целостность и полярность тканей, особенно эпителиального монослоя
Кадгерины • Е-кадгерин находится на поверхности клеток эпителиальных и эмбриональных тканей • N-кадгерин локализован на поверхности нервных клеток, хрусталика, сердца • Р-кадгерин расположен на клетках плаценты и эпидермиса Кадгерины участвуют в начальных стадиях адгезии, обеспечивают структурную целостность и полярность тканей, особенно эпителиального монослоя
 Селектины
Селектины
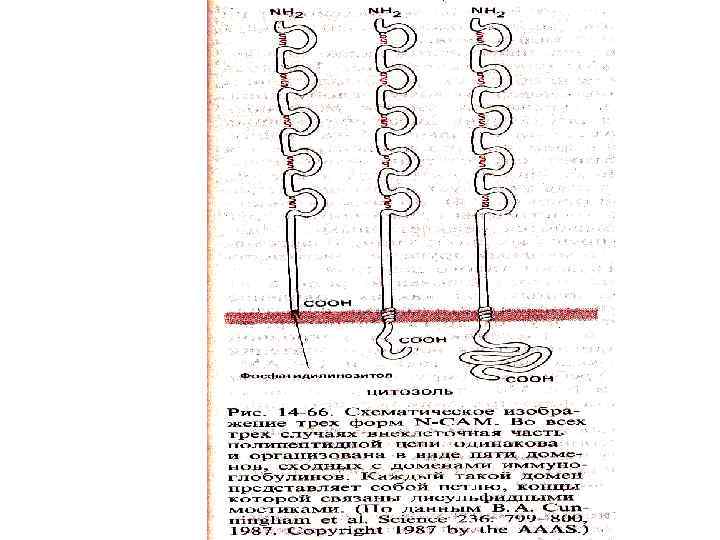
 Молекулы клеточной адгезии • Cell adhesion molecules – структура напоминает иммуноглобулины • Участвуют в передаче сигнала и связывании с белками цитоскелета • N- CAM - нейрональная
Молекулы клеточной адгезии • Cell adhesion molecules – структура напоминает иммуноглобулины • Участвуют в передаче сигнала и связывании с белками цитоскелета • N- CAM - нейрональная
 Лектины • Семейство неиммунных белков, специфически взаимодействующих с углеводными остатками гликопротеинов, протеогликанов и гликолипидов фитогемагглютинин (ФГА, из семян фасоли) конканавалин А (Кон А, из канавалии мечевидной), связывается с D-глюкозой и D-маннозой рицин (из семян клещевины) - с D-галактозой и N-ацетилгалактозамином
Лектины • Семейство неиммунных белков, специфически взаимодействующих с углеводными остатками гликопротеинов, протеогликанов и гликолипидов фитогемагглютинин (ФГА, из семян фасоли) конканавалин А (Кон А, из канавалии мечевидной), связывается с D-глюкозой и D-маннозой рицин (из семян клещевины) - с D-галактозой и N-ацетилгалактозамином
 Онкогены и онкобелки • Онкогены функционируют на ранних стадиях эмбриогенеза. Они реализуют программу нормального развития, участвуя в регуляции пролиферации Онкогены, участвующие в продукции мембранных онкобелков: • sis – поверхностный мембранный белок • erb B – белок, пронизывающий мембрану насквозь • sac и ras – находятся на внутренней поверхности мембраны
Онкогены и онкобелки • Онкогены функционируют на ранних стадиях эмбриогенеза. Они реализуют программу нормального развития, участвуя в регуляции пролиферации Онкогены, участвующие в продукции мембранных онкобелков: • sis – поверхностный мембранный белок • erb B – белок, пронизывающий мембрану насквозь • sac и ras – находятся на внутренней поверхности мембраны
 Межклеточные коммуникации
Межклеточные коммуникации
 продолжение
продолжение
 продолжение
продолжение
 Типы межклеточной сигнализации
Типы межклеточной сигнализации
 Эндокринная сигнализация • • 1. 2. 3. Гормоны секретируются в интерстициальное пространство Диффундируют к капиллярам Разносятся с кровью по организму Вновь проникают в интерстициальное пространство Взаимодействуют с рецепторами Скорость распространения гормона определяется скоростью диффузии и кровотока Для эндокринной сигнализации характерно: Низкая скорость передачи сигнала (минуты) Низкие действующие концентрации и высокое сродство к рецептору (Кδ 10 -8 - 10 -11 М) Специфичность сигнала определяется химической природой гормона и рецептора
Эндокринная сигнализация • • 1. 2. 3. Гормоны секретируются в интерстициальное пространство Диффундируют к капиллярам Разносятся с кровью по организму Вновь проникают в интерстициальное пространство Взаимодействуют с рецепторами Скорость распространения гормона определяется скоростью диффузии и кровотока Для эндокринной сигнализации характерно: Низкая скорость передачи сигнала (минуты) Низкие действующие концентрации и высокое сродство к рецептору (Кδ 10 -8 - 10 -11 М) Специфичность сигнала определяется химической природой гормона и рецептора
 • химический (<1 мс) этапы • Высокая") Синаптическая сигнализация Этапы: • Электрический (>100 м/с) • химический (<1 мс) этапы • Высокая скорость передачи сигнала • Высокая специфичность сигнала • Высокие действующие концентрации • Низкое сродство нейромедиатора к рецептору (Кδ ~ 10 -4 М)
Синаптическая сигнализация Этапы: • Электрический (>100 м/с) • химический (<1 мс) этапы • Высокая скорость передачи сигнала • Высокая специфичность сигнала • Высокие действующие концентрации • Низкое сродство нейромедиатора к рецептору (Кδ ~ 10 -4 М)
 Специфичность сигналов Сигнал – рецептор – клетка • Связывание сигнальной молекулы приводит к изменению свойств рецептора изменению функций клетки Ацетилхолин: • Уменьшает частоту и силу сердечных сокращений • Стимулирует сокращение скелетной мускулатуры • вызывает секрецию в секреторных клетках
Специфичность сигналов Сигнал – рецептор – клетка • Связывание сигнальной молекулы приводит к изменению свойств рецептора изменению функций клетки Ацетилхолин: • Уменьшает частоту и силу сердечных сокращений • Стимулирует сокращение скелетной мускулатуры • вызывает секрецию в секреторных клетках
") Вторичные мессенджеры 6. оксид азота (NO)
Вторичные мессенджеры 6. оксид азота (NO)
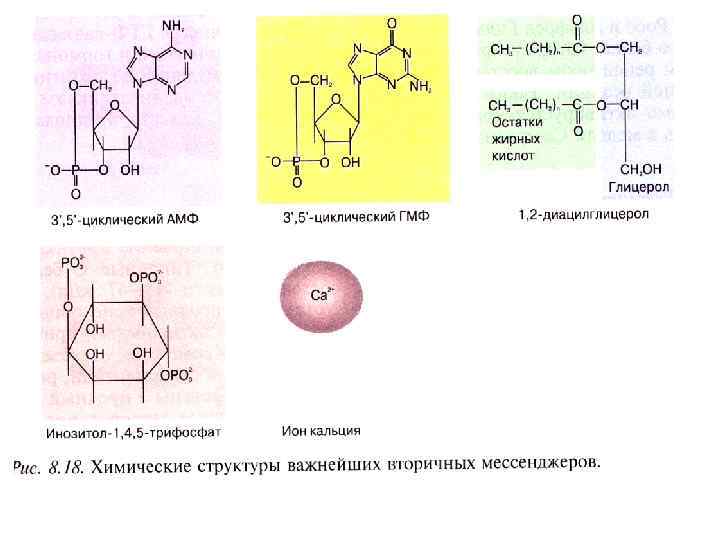


 Циклические нуклеотиды
Циклические нуклеотиды
 Фосфатидилинозитол
Фосфатидилинозитол
 Образование вторичных мессенджеров
Образование вторичных мессенджеров
 кальций
кальций

 Классификация первичных мессенджеров • Небольшие липофильные молекулы, диффундируют через плазматическую мембрану и взаимодействуют с внутриклеточными рецепторами • Липофильные молекулы, взаимодействуют с рецепторами клеточной поверхности. • Гидрофильные молекулы, взаимодействуют с рецепторами клеточной поверхности
Классификация первичных мессенджеров • Небольшие липофильные молекулы, диффундируют через плазматическую мембрану и взаимодействуют с внутриклеточными рецепторами • Липофильные молекулы, взаимодействуют с рецепторами клеточной поверхности. • Гидрофильные молекулы, взаимодействуют с рецепторами клеточной поверхности
 Активация рецепторов
Активация рецепторов
 Глюкокортикоиды
Глюкокортикоиды
 Стероидные гормоны • В крови стероидные гормоны находятся в виде комплекса со специфическими плазматическими белкамипереносчиками. • В непосредственной близости от плазматической мембраны клетки-мишени гормон отделяется от транспортного белка и диффундирует внутрь клетки. • В типичной клетке-мишени присутствует от 1000 до 10 000 рецепторов стероидных гормонов, обладающих высоким сродством к лиганду (Кd 10 -8— 10 -10 М). • Рецепторы стероидных гормонов имеют высокую избирательность в отношении их лигандов. • В зависимости от типа рецептора местом их локализации в отсутствие гормона может быть цитоплазма или ядро.
Стероидные гормоны • В крови стероидные гормоны находятся в виде комплекса со специфическими плазматическими белкамипереносчиками. • В непосредственной близости от плазматической мембраны клетки-мишени гормон отделяется от транспортного белка и диффундирует внутрь клетки. • В типичной клетке-мишени присутствует от 1000 до 10 000 рецепторов стероидных гормонов, обладающих высоким сродством к лиганду (Кd 10 -8— 10 -10 М). • Рецепторы стероидных гормонов имеют высокую избирательность в отношении их лигандов. • В зависимости от типа рецептора местом их локализации в отсутствие гормона может быть цитоплазма или ядро.
, включают ряд доменов: • Гормон-связывающий") Рецепторы стероидных гормонов являются белками (50— 120 к. Да), включают ряд доменов: • Гормон-связывающий домен содержит участок связывания гормона; • ДНК-связывающий и сайт-специфический домены участвуют в узнавании специфических нуклеотидных гормонреспонсивных элементов (HRE) • Регуляторный домен А/В, содержит участки связывания с компонентами клеточного ядра, обеспечивает транслокацию и внутриядерную компартментализацию рецептора. • Гормон-респонсивные элементы представляют собой короткие симметричные участки ДНК, которые служат энхансерами транскрипции.
Рецепторы стероидных гормонов являются белками (50— 120 к. Да), включают ряд доменов: • Гормон-связывающий домен содержит участок связывания гормона; • ДНК-связывающий и сайт-специфический домены участвуют в узнавании специфических нуклеотидных гормонреспонсивных элементов (HRE) • Регуляторный домен А/В, содержит участки связывания с компонентами клеточного ядра, обеспечивает транслокацию и внутриядерную компартментализацию рецептора. • Гормон-респонсивные элементы представляют собой короткие симметричные участки ДНК, которые служат энхансерами транскрипции.
 • Энхансер транскрипции, характерный для глюкокортикоидов, отличается по нуклеотидной последовательности от энхансеров других стероидных гормонов • Ген, регулируемый гормоном, может содержать несколько (2— 5) энхансерных участков • они могут располагаться до начала кодирующей последовательности или непосредственно в области гена
• Энхансер транскрипции, характерный для глюкокортикоидов, отличается по нуклеотидной последовательности от энхансеров других стероидных гормонов • Ген, регулируемый гормоном, может содержать несколько (2— 5) энхансерных участков • они могут располагаться до начала кодирующей последовательности или непосредственно в области гена
 • В неактивном состоянии рецептор стероидного гормона присутствует в клетке в виде комплекса с белкомингибитором. • Для эстрогенов и кортизола белкомингибитором служит шаперонный белок ( -90 к. Да), или белок теплового шока HSP-90. • Связывание с гормоном вызывает конформационные изменения в молекуле рецептора, приводящие к уменьшению его сродства к белкуингибитору и диссоциации комплекса с освобождением и последующей димеризацией активных рецепторов. Димеризация вызывает повышение сродства комплекса к ДНК.
• В неактивном состоянии рецептор стероидного гормона присутствует в клетке в виде комплекса с белкомингибитором. • Для эстрогенов и кортизола белкомингибитором служит шаперонный белок ( -90 к. Да), или белок теплового шока HSP-90. • Связывание с гормоном вызывает конформационные изменения в молекуле рецептора, приводящие к уменьшению его сродства к белкуингибитору и диссоциации комплекса с освобождением и последующей димеризацией активных рецепторов. Димеризация вызывает повышение сродства комплекса к ДНК.
 Что дальше? • • • Связывание рецепторного комплекса с энхансерным участком вызывает стимуляцию транскрипции соседних генов. Стероидный гормон может стимулировать экспрессию сразу нескольких белков путем двухстадийной стимуляции экспрессии генов. На первой стадии инициируется экспрессия нескольких белков в результате непосредственной индукции транскрипции димерными лиганд-рецепторными комплексами (первичный ответ). На второй стадии белки первичного ответа активируют ряд других генов и вызывают экспрессию белков, выполняющих другие физиологические функции (вторичный ответ). Один и тот же гормон может вызывать различные физиологические эффекты в разных тканях. Физиологические эффекты стероидных гормонов определяются природой гормона и типом клетки-мишени.
Что дальше? • • • Связывание рецепторного комплекса с энхансерным участком вызывает стимуляцию транскрипции соседних генов. Стероидный гормон может стимулировать экспрессию сразу нескольких белков путем двухстадийной стимуляции экспрессии генов. На первой стадии инициируется экспрессия нескольких белков в результате непосредственной индукции транскрипции димерными лиганд-рецепторными комплексами (первичный ответ). На второй стадии белки первичного ответа активируют ряд других генов и вызывают экспрессию белков, выполняющих другие физиологические функции (вторичный ответ). Один и тот же гормон может вызывать различные физиологические эффекты в разных тканях. Физиологические эффекты стероидных гормонов определяются природой гормона и типом клетки-мишени.
 Глюкокортикоиды
Глюкокортикоиды
 Эйкозаноиды
Эйкозаноиды
") Активация рецепторов гидрофильными молекулами ( пептидные гормоны, факторы роста, цитокины, нейротрансмиттеры)
Активация рецепторов гидрофильными молекулами ( пептидные гормоны, факторы роста, цитокины, нейротрансмиттеры)
 • В отличие от внутриклеточных рецепторов стероидных и тиреоидных гормонов рецепторы клеточной поверхности не участвуют непосредственно в регуляции экспрессии генов, они передают сигнал первичного мессенджера через плазматическую мембрану.
• В отличие от внутриклеточных рецепторов стероидных и тиреоидных гормонов рецепторы клеточной поверхности не участвуют непосредственно в регуляции экспрессии генов, они передают сигнал первичного мессенджера через плазматическую мембрану.
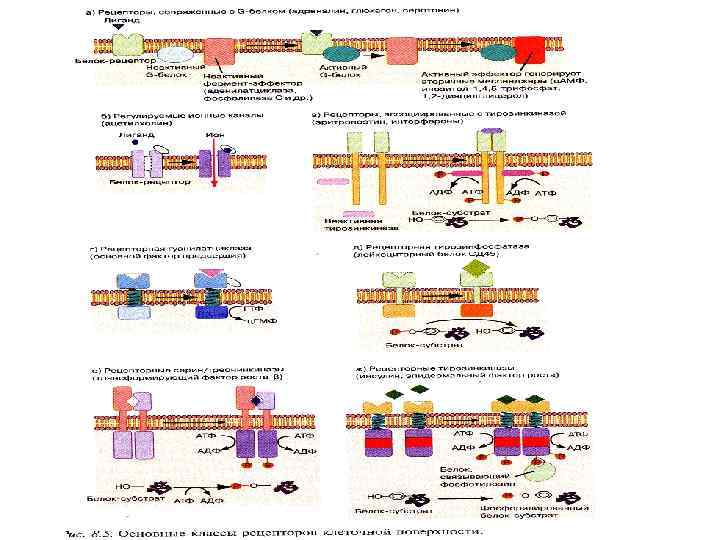
 Рецепторы клеточной поверхности • Рецепторы, сопряженные с Gбелками • Регулируемые ионные каналы • Рецепторы, ассоциированные с цитозольными тирозиновыми протеинкиназами • Каталитические рецепторы Гуанилатциклаза Тирозинфосфатаза Серин/треонинкиназа Тирозинкиназа
Рецепторы клеточной поверхности • Рецепторы, сопряженные с Gбелками • Регулируемые ионные каналы • Рецепторы, ассоциированные с цитозольными тирозиновыми протеинкиназами • Каталитические рецепторы Гуанилатциклаза Тирозинфосфатаза Серин/треонинкиназа Тирозинкиназа
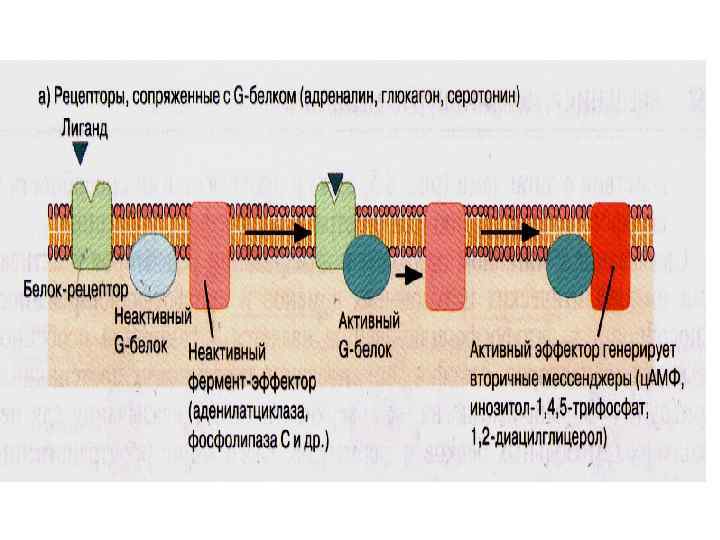
 Рецепторы, сопряженные с G-белками • Связывание лиганда с рецептором приводит к активации G-белков Активированный G-белок изменяет активность молекулярных мишеней – аденилатциклазы фосфолипазы ионных каналов с. GMP- фосфодиэстеразы • Рецепторы этого типа реагируют на небольшие молекулы (катехоламины, пептиды, хемокины) Лиганды: Адреналин, глюкагон, серотонин • Имеют широкую субстратную специфичность
Рецепторы, сопряженные с G-белками • Связывание лиганда с рецептором приводит к активации G-белков Активированный G-белок изменяет активность молекулярных мишеней – аденилатциклазы фосфолипазы ионных каналов с. GMP- фосфодиэстеразы • Рецепторы этого типа реагируют на небольшие молекулы (катехоламины, пептиды, хемокины) Лиганды: Адреналин, глюкагон, серотонин • Имеют широкую субстратную специфичность

, либо с ингибиторным (Gi)") • Различные типы рецепторов взаимодействуют либо со стимуляторным (Gs), либо с ингибиторным (Gi) белком. • Адреналин связывается с четырьмя различными типами адренэргических рецепторов: α 1, α 2, β 1, β 2. • Связывание адреналина с α 1 -рецепторами - не влияет на аденилатциклазу • С β 1 и β 2 –рецепторами – стимулирует • С α 2 – рецептором ингибирует аденилатциклазу
• Различные типы рецепторов взаимодействуют либо со стимуляторным (Gs), либо с ингибиторным (Gi) белком. • Адреналин связывается с четырьмя различными типами адренэргических рецепторов: α 1, α 2, β 1, β 2. • Связывание адреналина с α 1 -рецепторами - не влияет на аденилатциклазу • С β 1 и β 2 –рецепторами – стимулирует • С α 2 – рецептором ингибирует аденилатциклазу
 Эффект зависит от типа рецептора
Эффект зависит от типа рецептора
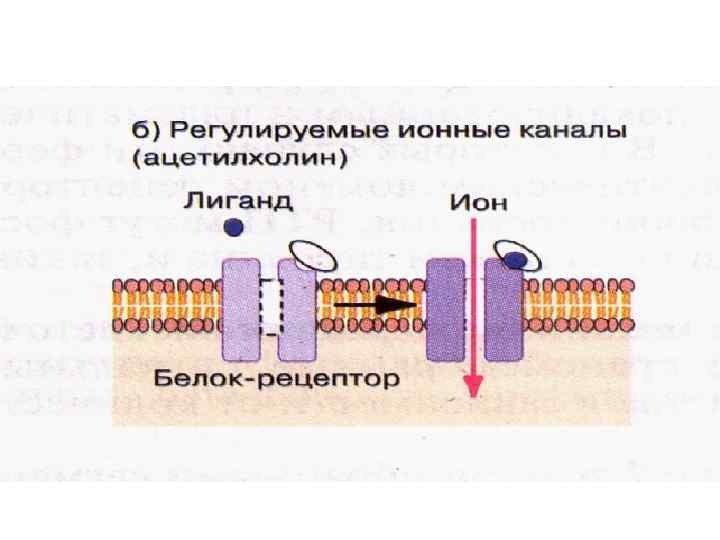
 • Связывание лиганда с рецепторным белком канала вызывает открытие канала,") Регулируемые ионные каналы (ацетилхолин) • Связывание лиганда с рецепторным белком канала вызывает открытие канала, приводящее к входу или выходу необходимых ионов • Результатом активации канала является изменение мембранного потенциала или внутриклеточного уровня вторичных мессенджеров
Регулируемые ионные каналы (ацетилхолин) • Связывание лиганда с рецепторным белком канала вызывает открытие канала, приводящее к входу или выходу необходимых ионов • Результатом активации канала является изменение мембранного потенциала или внутриклеточного уровня вторичных мессенджеров
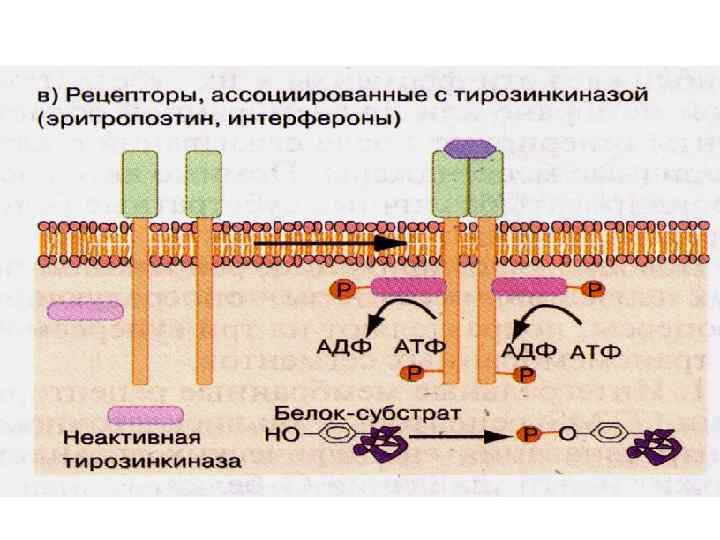
 • Присоединение лиганда приводит к димеризации рецепторов, активации") Рецепторы, ассоциированные с тирозинкиназой (эритропоэтин, интерфероны) • Присоединение лиганда приводит к димеризации рецепторов, активации киназного каталитического домена, который фосфорилирует цитоплазматические субстраты Рецепторами могут быть: тирозинкиназы, треонинкиназы • Часто разделены на сигнал-передающие субъединицы • Оказывают дублирующее и плейотропное действие • Суперсемейство цитокиновых рецепторов
Рецепторы, ассоциированные с тирозинкиназой (эритропоэтин, интерфероны) • Присоединение лиганда приводит к димеризации рецепторов, активации киназного каталитического домена, который фосфорилирует цитоплазматические субстраты Рецепторами могут быть: тирозинкиназы, треонинкиназы • Часто разделены на сигнал-передающие субъединицы • Оказывают дублирующее и плейотропное действие • Суперсемейство цитокиновых рецепторов
 Каталитические рецепторы Гуанилатциклаза Тирозинфосфатаза Серин/треонинкиназа Тирозинкиназа
Каталитические рецепторы Гуанилатциклаза Тирозинфосфатаза Серин/треонинкиназа Тирозинкиназа
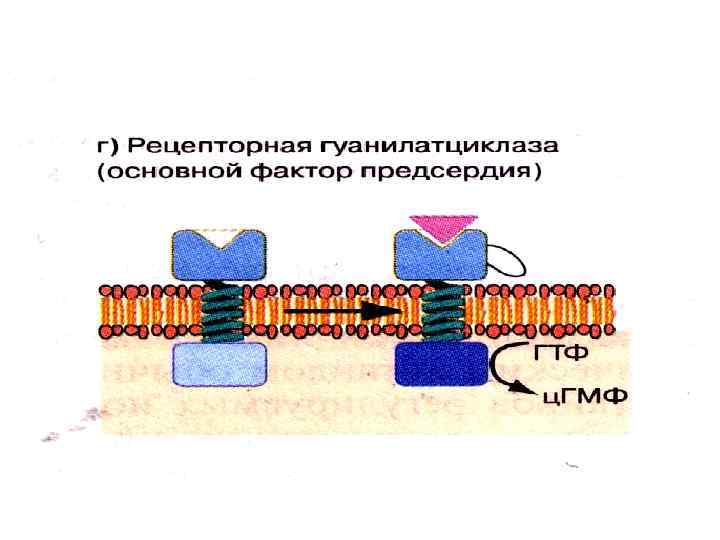
 • Связывание ПНФ вызывает активацию каталитического домена") Рецепторная гуанилатциклаза Рецептор предсердного натрийуретического фактора (ПНФ) • Связывание ПНФ вызывает активацию каталитического домена гуанилатциклазы • Активация этого рецептора приводит к входу ионов натрия в клетку
Рецепторная гуанилатциклаза Рецептор предсердного натрийуретического фактора (ПНФ) • Связывание ПНФ вызывает активацию каталитического домена гуанилатциклазы • Активация этого рецептора приводит к входу ионов натрия в клетку
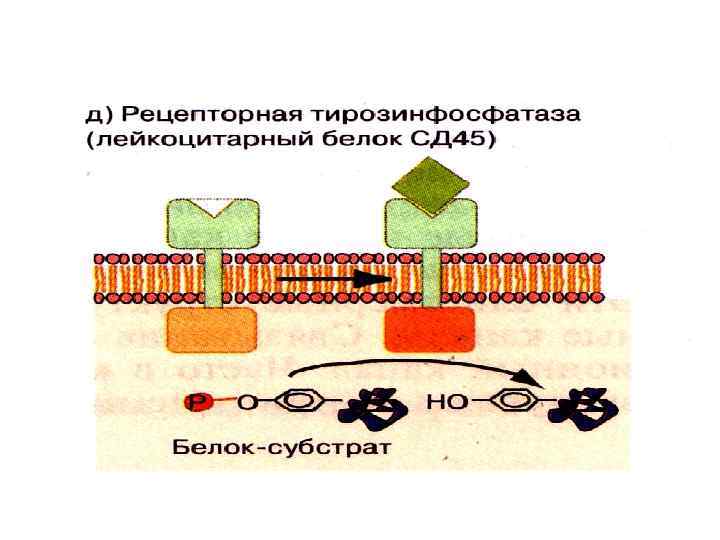
 Рецепторы с фосфатазной активностью • Лиганд активирует цитоплазматический фосфатазный домен
Рецепторы с фосфатазной активностью • Лиганд активирует цитоплазматический фосфатазный домен
 Характеристика каталитических рецепторов • Являются аллостерическими ферментами • Осуществляют автофосфорилирование • Димеризация рецепторов способствует каталитической тирозинкиназной активности • • Автофосфорилирование является сигналом к рецептор-опосредованному эндоцитозу лигандрецепторных комплексов • Рецептор-опосредованный эндоцитоз инактивирует сигнальные молекулы и снижает плотность рецепторов на клеточной поверхности
Характеристика каталитических рецепторов • Являются аллостерическими ферментами • Осуществляют автофосфорилирование • Димеризация рецепторов способствует каталитической тирозинкиназной активности • • Автофосфорилирование является сигналом к рецептор-опосредованному эндоцитозу лигандрецепторных комплексов • Рецептор-опосредованный эндоцитоз инактивирует сигнальные молекулы и снижает плотность рецепторов на клеточной поверхности
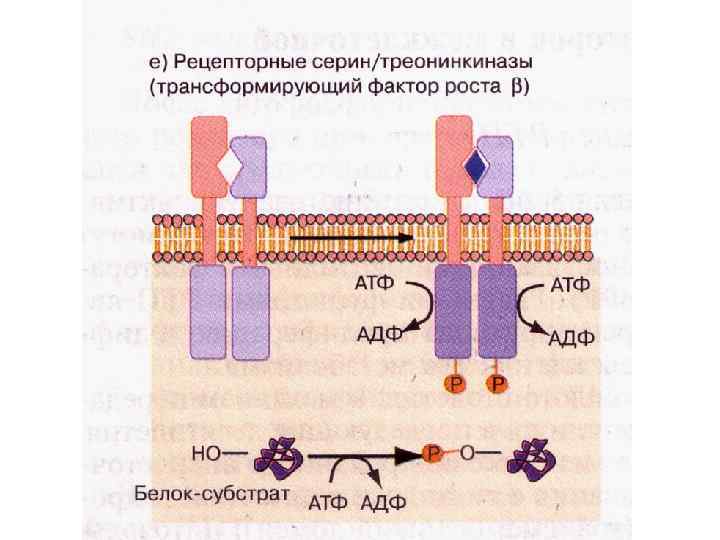
 • При взаимодействии с лигандом происходит автофосфорилирование каталитических") Рецепторные серин/треонинкиназы (трансформирующий фактор роста β) • При взаимодействии с лигандом происходит автофосфорилирование каталитических доменов • Фосфорилирование и активация цитозольных факторов транскрипции Smad • В ядре Smad индуцирует транскрипцию генов, ответственных за действие ТФРβ
Рецепторные серин/треонинкиназы (трансформирующий фактор роста β) • При взаимодействии с лигандом происходит автофосфорилирование каталитических доменов • Фосфорилирование и активация цитозольных факторов транскрипции Smad • В ядре Smad индуцирует транскрипцию генов, ответственных за действие ТФРβ
 • При связывании лиганда с рецептором активируются тирозинкиназы,") Рецепторные тирозинкиназы (инсулин, эпидермальный фактор роста) • При связывании лиганда с рецептором активируются тирозинкиназы, нековалентно связанные с поверхностным рецептором • Происходит димеризация рецепторов с последующим трансфосфорилированием
Рецепторные тирозинкиназы (инсулин, эпидермальный фактор роста) • При связывании лиганда с рецептором активируются тирозинкиназы, нековалентно связанные с поверхностным рецептором • Происходит димеризация рецепторов с последующим трансфосфорилированием
") Рецепторные тирозинкиназы (РТП)
Рецепторные тирозинкиназы (РТП)
 SH 2 -белки • • Src homology domain связывает: Адапторные белки GRB 2 и Shc Эстафетный белок IRS 1(insulin reseptor substrate) GAP (белок, активирующий ГТФазу) Протеинфосфатазы SYP и Ship (фосфатаза ИФ 5 Фосфатидилинозитол-3 -киназу и фосфолипазу Сγ Факторы транскрипции STAT 1 -5 Cтруктурный белок тензин (фокальная адгезия)
SH 2 -белки • • Src homology domain связывает: Адапторные белки GRB 2 и Shc Эстафетный белок IRS 1(insulin reseptor substrate) GAP (белок, активирующий ГТФазу) Протеинфосфатазы SYP и Ship (фосфатаза ИФ 5 Фосфатидилинозитол-3 -киназу и фосфолипазу Сγ Факторы транскрипции STAT 1 -5 Cтруктурный белок тензин (фокальная адгезия)
 RAS - белок • Переключатель, способен запускать и отменять внутриклеточный каскад пердачи сигнала • Неактивное состояние RAS –ГДФ • Связывание факторов роста индуцирует образование RAS -ГТФ
RAS - белок • Переключатель, способен запускать и отменять внутриклеточный каскад пердачи сигнала • Неактивное состояние RAS –ГДФ • Связывание факторов роста индуцирует образование RAS -ГТФ
 Ras-белок
Ras-белок
 Взаимодействие Ras-белка с РТП
Взаимодействие Ras-белка с РТП
 • Связывание") Передача регуляторного сигнала • Связывание Ras с N-концевым доменом Raf (серин/треониновая протеинкиназа) • Связывание и фосфорилирование Мек (тирозин/сериновая протеинкиназа) • Фосфорилирование и активация Маркиназы (серин/треониновая протеинкиназа) • Изменение метаболизма, секреторной, пролиферативной, миграционной активности клетки
Передача регуляторного сигнала • Связывание Ras с N-концевым доменом Raf (серин/треониновая протеинкиназа) • Связывание и фосфорилирование Мек (тирозин/сериновая протеинкиназа) • Фосфорилирование и активация Маркиназы (серин/треониновая протеинкиназа) • Изменение метаболизма, секреторной, пролиферативной, миграционной активности клетки
 Передача сигнала Ras
Передача сигнала Ras
 Типы РТП
Типы РТП
 РТП – аллостерический фермент
РТП – аллостерический фермент