Ткани 61-139.ppt.pptx
- Количество слайдов: 80



МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ Общепринятой теорией является модель мышечного сокращения, предложенная Х. Хаксли (в 1954 г. ). Это теория скольжения нитей. Суть этой теории в следующем: • нервный импульс проходит по нервному волокну и передаётся на плазмолемму мышечного волокна; • электрический импульс идёт по Т-трубочке вглубь мышечного волокна и передаётся на лежащие рядом Т-цистерны СПР; • мембраны СПР после деполяризации становятся проницаемыми для ионов; • ионы кальция выходят из Т-цистерны, связываются с молекулами Тn. C;

• при связывании ионов кальция изменяется конфигурация тропонинтропомиозинового комплекса и на актиновых филаментах и открываются активные центры для связывания головок миозина (которые в состоянии расслабления были закрыты Tn. I); • головка миозина связывается с актином, а АТФ расщепляется до АДФ, давая энергию для движения миозиновой головки; • головки изгибаются в шарнирных областях и создают тянущиеся усилия; • это вызывает скольжение тонких нитей между толстыми → тонкие нити более глубоко вдвигаются в А-диск.

Скользящие тонкие нити тянут за собой Z- линии, вызывая сокращение длины саркомера

Длина филаментов при сокращении не изменяется, но поскольку актиновые филаменты заходят глубже между миозиновыми филаментами: • длина И-дисков уменьшается; • длина Н-полоски уменьшается, хотя длина А-диска остается неизменной; • Z-линии сближаются и уменьшается протяженность саркомера в целом

• Отдельное мышечное сокращение является результатом сотен циклов образования и разъединения мостиков. Активное сокращение, которое приводит к полному взаимному перекрыванию между тонкими и толстыми филаментами, продолжается до тех пор, пока не будут удалены ионы Са. • При отсутствии нервных импульсов ионы кальция вновь откачиваются в СПР, и тропонин-тропомиозиновый комплекс вновь закрывает участки связывания миозина на актине.

ВОЗМОЖНОСТИ РЕГЕНЕРАЦИИ СКЕЛЕТНОЙ МЫШЕЧНОЙ ТКАНИ • Скелетная мышечная ткани способна регенерировать как на внутриклеточном уровне (образование в миофибриллах новых саркомеров, новых органелл), так и на клеточном уровнях. • Клеточная регенерация происходит благодаря активации миосателлитоцитов вблизи зоны травмы мышечных волокон. • Миосателлитоциты образуют миобласты, которые быстро делятся, сливаются друг с другом и формируют мышечные трубочки. Мышечные трубочки постепенно превращаются в полноценные мышечные волокна.

СТРОЕНИЕ СКЕЛЕТНОЙ МЫШЦЫ КАК ОРГАНА • • Мышца состоит из множества мышечных волокон, связанных в единое целое соединительной тканью. Между мышечными волокнами лежат тонкие прослойки рыхлой волокнистой соединительной ткани, которая называется эндомизием. Несколько мышечных волокон (от 10 до 100) окружены более толстыми прослойками рыхлой волокнистой соединительной ткани – перимизием. В эндомизии и перимизии проходят кровеносные сосуды, питающие мышцу и нервные волокна. Снаружи мышца покрыта оболочкой из плотной волокнистой соединительной ткани - эпимизием.

ТИПЫ МЫШЕЧНЫХ ВОЛОКОН Выделяют три основных типа мышечных волокон: • I тип – красные мышечные волокна. Это медленные тонические мышечные волокна: они могут сокращаться в течение длительного времени, но медленно. Красные мышечные волокна имеют небольшой диаметр, содержат относительно тонкие миофибриллы, много митохондрий, имеют высокую активность окислительно-восстановительных ферментов и много липидных включений. Красный цвет волокна связан с тем, что в саркоплазме этих волокон очень много белка миоглобина. Содержат много миосателлитоцитов и кровеносных сосудов. • II В тип – белые мышечные волокна. Это быстрые тетанические мышечные волокна: они способны вызывать сокращения большой силы, но быстро утомляются. Характеризуются большим диаметром, сильным развитием миофибрилл, меньшим количеством митохондрий и запасами питательных веществ в виде гликогена. Миоглобина в саркоплазме мало. В волокнах низкая активность окислительных ферментов и, напротив, высокая активность гликолитических ферментов. Кровоснабжение относительно слабое. • II А тип – промежуточный тип мышечных волокон. Занимают и в структурном, и в функциональном отношении среднее положение между красными и белыми мышечными волокнами.

СЕРДЕЧНАЯ МЫШЕЧНАЯ ТКАНЬ

ТИПЫ КАРДИОМИОЦИТОВ В результате дифференцировки кардиомиоциты превращаются в клетки трёх типов: • рабочие, или типичные, сократительные; • импульс-генерирующие и проводящие; • секреторные (эндокринные).

Строение кардиомиоцита. А — схема; Б — электронная микрофотография вставочного диска: 1 — миофибриллы; 2 — митохондрии; 3 — саркотубулярная сеть; 4 — Т-трубочки 5 — базальная мембрана; 6 — лизосома; 7 — вставочный диск; 8 — десмосома; 9 — зона прикрепления миофибрилл; 10 — щелевые контакты; 11 — гликоген.

• • Сердечная мышечная ткань образована клетками кардиомиоцитами. Сердечное мышечное волокно состоит из цепочки кардиомиоцитов, соединенных конец вставочными дисками. Кардиомиоциты одного волокна имеют ответвления и их боковые поверхности образуют анастомозы с соседними волокнами. В результате межклеточных контактов миокард представляет собой сложную трёхмерную сеть, которая ведёт себя как функциональный синцитий.

• Сократительный кардиомиоцит имеет вытянутую цилиндрическую слабоотростчатую форму. Крупное светлое ядро кардиомиоцита находится в центре клетки. Многие клетки имеют два ядра и являются полиплоидными. • Цитоплазма (саркоплазма) кардиомиоцита окрашивается оксифильно. Периферическую часть саркоплазмы занимают расположенные продольно исчерченные миофибриллы, построенные так же, как и в скелетной мышечной ткани.

• Между миофибриллами цепочками лежат очень крупные митохондрии с высокой плотностью крист. Митохондрии занимают до 40% объёма цитоплазмы, что связано с огромной энергетической нагрузкой. В цитоплазме кардиомиоцитов содержится большое количество включений, особенно липидных капель.

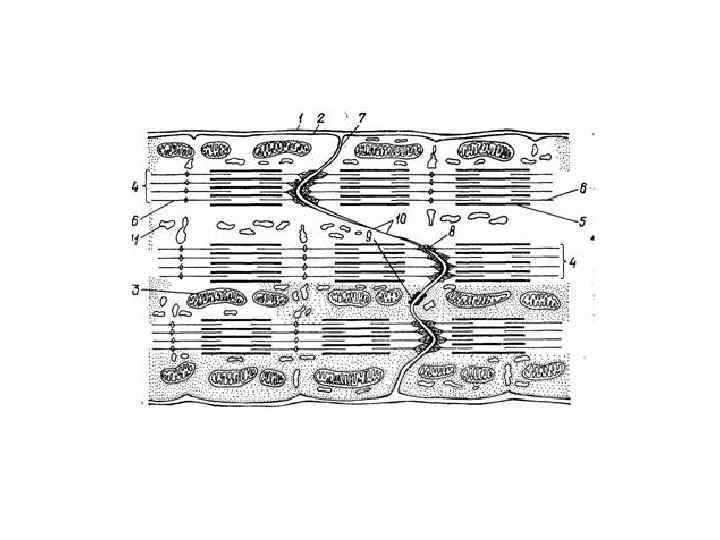
• Вставочный диск – это место контактов двух кардиомиоцитов. • Вставочный диск - комплекс межклеточный контактов, которые обеспечивают как механическую, так и химическую, функциональную коммуникацию кардиомиоцитов. • В световом микроскопе вставочные диски имеют вид тёмных поперечных полосок, более толстых и интенсивно окрашенных в сравнении с дисками А миофибрилл. • В электронном микроскопе вставочные диски имеют вид зубчатой линии (интердигитации) или ступенек.

В этой зубчатой линии можно выделить горизонтальные и вертикальные участки и три зоны: • Зоны десмосом и полосок слипания (fascia adherens). Находятся на вертикальных (поперечных) участках диска. Обеспечивают механическое соединение кардиомиоцитов, препятствуют их отделению во время циклов сокращения. • Зоны прикрепления миофибрилл. Находятся на вертикальных (поперечных) участках диска. Служат местами прикрепления актиновых филаментов конечных саркомеров к плазмолемме кардиомиоцитов и аналогичны Zлиниям. • Зоны нексусов (щелевых контактов) – места передачи возбуждения с одной клетки на другую, обеспечивают распространение деполяризации, вызывающей сокращение. Находятся на горизонтальных (продольных) участках диска.

Особенности строения секреторных кардиомиоцитов • • • Секреторные кардиомиоциты локализуются в основном в правом предсердии. В отличии от сократительных кардиомиоцитов в цитоплазме этих клеток хорошо развит секреторный аппарат: гранулярная эндоплазматическая сеть и комплекс Гольджи, и обнаруживаются многочисленные электронно-плотные секреторные гранулы. Эти гранулы содержат пептидный гормон – натрийуретический фактор (кардиодилатин). Этот гормон оказывает различные эффекты: усиливает секрецию натрия почками, расслабляет гладкие миоциты стенки артерий, подавляет секрецию гормонов, вызывающих гипертензию (альдостерона и вазопрессина). Всё это ведёт к увеличению диуреза и просвета артерий, снижению объёма циркулирующей жидкости и в результате – к снижению артериального давления.

РЕГЕНЕРАЦИЯ СЕРДЕЧНОЙ МЫШЕЧНОЙ ТКАНИ • Поскольку во взрослом организме в сердечной мышечной ткани нет камбиальных клеток, регенерация протекает на внутриклеточном уровне. • При повышенной нагрузке на сердце происходит гипертрофия (увеличение размеров) и гиперплазия (увеличение количества) органелл, в том числе миофибрилл. • При ранениях сердечной мышцы, инфарктах миокардах на месте погибших клеток образуется рубец из соединительной ткани.

ГЛАДКАЯ МЫШЕЧНАЯ ТКАНЬ

• Структурно-функциональным тканевым элементом является гладкий миоцит. Гладкий миоцит – клетка веретеновидной формы, то есть толщина максимальна в среднем участке, а к концам конически сужается. • Длина гладкого миоцита может колебаться от 20 мкм (в мелких кровеносных сосудов) до 500 мкм (в матке при беременности). • В каждой клетке имеется одно ядро палочковидную или эллипсоидную форму, с плотным хроматином, расположенное в центре наиболее широкой части клетки. Плазмолемма клеток покрыта тонкой базальной мембраной.

• В цитоплазме у полюсов ядра концентрируются элементы секреторного аппарата: эндоплазматическая сеть – место синтеза белков межклеточного вещества, комплекс Гольджи, а также мелкие митохондрии.

• Плотные тельца состоят из белка αактинина; аналоги Zлиний саркомеров. Две разновидности: • связанные с внутренней поверхностью сарколеммы; • свободно лежащие в цитоплазме в виде правильной цепочки. • К плотным тельцам прикрепляются актиновые и промежуточные десминовые филаменты.

• В гладких миоцитах нет исчерченных миофибрилл, сократительный аппарат представлен тонкими актиновыми филаментами, которые располагаются под углом крест-накрест, образуя сеть. Филаменты закреплены в плотных тельцах. • Миозиновые нити в ГМК лабильны – происходит постоянная сборка и разборка при сокращении и расслаблении соответственно.

• Механизм сокращения гладких миоцитов принципиально сходен с сокращением исчерченных мышечных тканей: взаимодействие актиновых и миозиновых филаментов. • Под действием нервного импульса из пиноцитозных пузырьков высвобождается кальций, который образует комплекс с белком кальмодулином, связывающим Са → комплекс «Са-кальмодулин» активирует фермент киназу лёгких цепей миозина → фосфорилирование миозина придаёт ему способность взаимодействовать с актиновыми филаментами → в результате движений миозиновых филамент вдоль актиновых сближаются плотные тельца, и гладкий миоцит сокращается.

• При сокращении гладкой мышечной ткани границы клеток становятся фестончатыми, а ядро – складчатым или штопорообразным. Плотные тельца передают сократительное усилие на соседние ГМК и окружающую их сеть ретикулярных волокон.

• Промежуточные десминовые филаменты препятствуют сильной деформации клетки при её сокращении. • Гидролиз АТФ происходит в ГМК медленно, что отражается на скорости сокращения. • Прекращает сокращение фермент фосфатаза миозина. При этом особенность гладких мышц заключается в том, что не все миозиновые мостики после дефосфорилирования разрушаются. Это обеспечивает длительное поддержание тонуса гладких мышц без дополнительных энергетических затрат.

• Гладкие миоциты функционируют не изолированно, а образуют миоцитарные комплексы из 10 -12 ГМК. • Нервные окончания подходят не ко всем миоцитам, а только к одному из комплекса. Миоциты тесно взаимодействуют друг с другом при помощи нексусов. В области нексусов базальные мембраны прерываются. • Через нексусы происходит передача возбуждения, и сокращение охватывает весь комплекс.
 1 — эндотелиоциты, 1. 1 — ядро, 1. 2")
Эндотелий магистрального сосуда (плоскостной препарат) 1 — эндотелиоциты, 1. 1 — ядро, 1. 2 — цитоплазма, 2 — межклеточные границы.

Регенерация гладкой мышечной ткани • Гладкая мышечная ткань способна к активной регенерации: • клеточная регенерация - за счёт делений малодифференцированных клеток; • внутриклеточная регенерация – гипертрофия, восстановление органелл.

НЕРВНАЯ ТКАНЬ
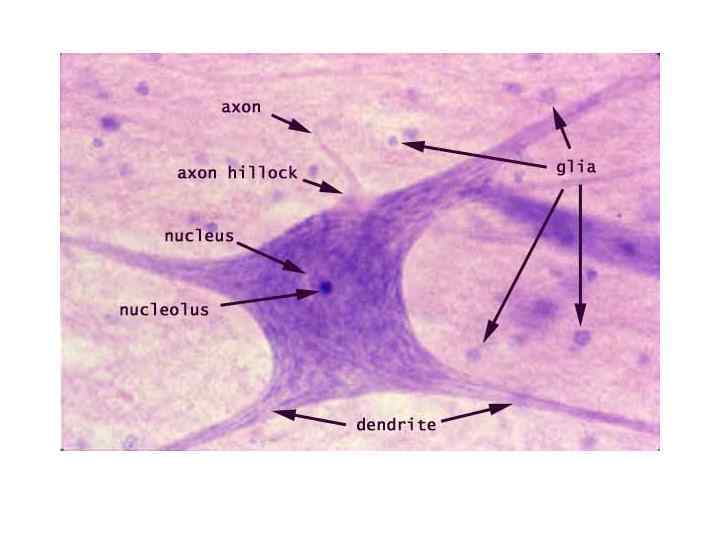
 1 — тела мультиполярных двигательных нейронов; 2")
Спинной мозг. Участок серого вещества (передние рога) 1 — тела мультиполярных двигательных нейронов; 2 — глиоциты; 3 — нейропиль; 4 — кровеносные сосуды.
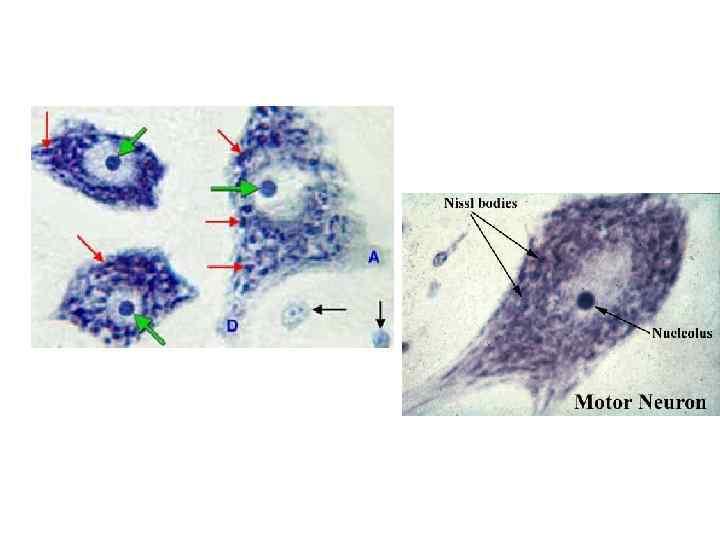
 в мультиполярных нейронах спинного мозга. Окраска по Нисслю • •")
Хроматофильная субстанция (субстанция Ниссля) в мультиполярных нейронах спинного мозга. Окраска по Нисслю • • • Метод Ниссля основан на окраске рибонуклеопротеидов (рибосомы и Гр. ЭС) основными красителями (толуидиновым синим, крезилвиолетом). На малом увеличении найти крупный мультиполярный (моторный) нейрон в переднем роге серого вещества спинного мозга. При большом увеличении обратить внимание на светлое ядро, хорошо заметное ядрышко, на наличие глыбок хроматофильной субстанции в теле и дендритах нейрона и на отсутствие их в аксоне и аксональном холмике.

Нейрофибриллы в нейронах передних рогов спинного мозга. Окраска – импрегнация серебром. • Соли серебра окрашивают ядрышко и нейрофибриллы в коричневый или чёрный цвет. • При малом увеличении найти крупный нейрон в передних рогах спинного мозга. Нейрофибриллы в теле нейрона образуют сеть, а в отростках идут параллельно другу. Ядро светлое.

Разновидности клеток нейроглии. а — протоплазматические астроциты; б — фиброзные астроциты; в — микроглия; г — олигодендроглия. Клетки а и б образуют «ножки» на кровеносных сосудах мозга, участвуя в формировании гематоэнцефалического барьера.
 в спинно-мозговом узле. Окр. : Г. -Э. • • На")
Глиоциты ганглия (мантийные глиоциты) в спинно-мозговом узле. Окр. : Г. -Э. • • На малом увеличении найти крупные округлые клетки со светлым ядром, расположенные группами по периферии узла. Это псевдоуниполярные нейроны. Отростки нейронов при данной окраске не выявляются. При большом увеличении заметно, что нейроны окружены оболочкой из мелких мантийных глиоцитов. Цитоплазма глиоцитов практически не заметна, но хорошо видны их мелкие округлые плотные ядра.

Мозжечок. Участок коры 1— молекулярный слой: 1. 1— дендриты клеток Пуркинье; 1. 2 — нейроны молекулярного слоя; 2— ганглионарный слой; 2. 1 — тела грушевидных нейронов (клеток Пуркинье), 2. 2 — «корзинки» , образованные коллатералями аксонов корзинчатых клеток; 3 —зернистый слой. 3. 1 —тела клетокзёрен, 3. 2 — нейриты клеток Пуркинье; 4 — белое вещество.

Полушарие большого мозга. Цитоархитектоника 1 — мягкая мозговая оболонка; 2 — серое вещество. 2. 1 — молекулярный слой, 2. 2 — наружный зернистый слой, 2. 3 — наружный пирамидный cлой, 2. 4 — внутренний зернистый слой. 2. 5 — внутренний пирамидный (ганглионарный) слой, 2. 6 — слой полиморфных клеток; 3 — белое вещество.
. Окраска осмиевой кислотой. Осмиевая кислота окрашивает миелиновую")
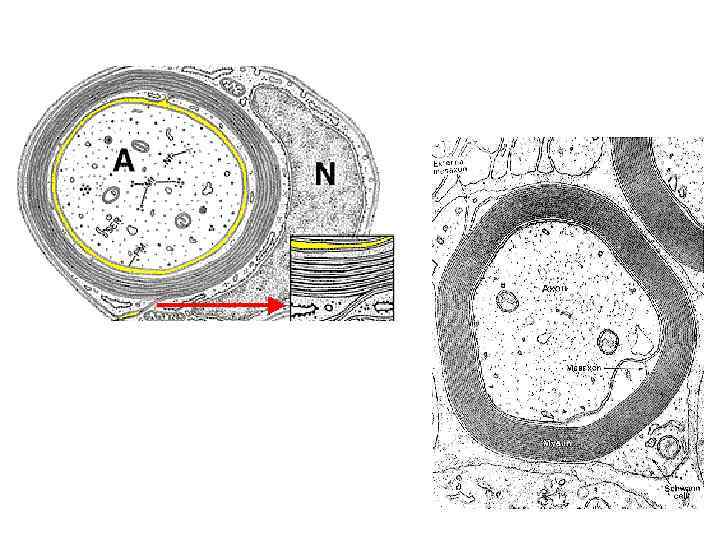
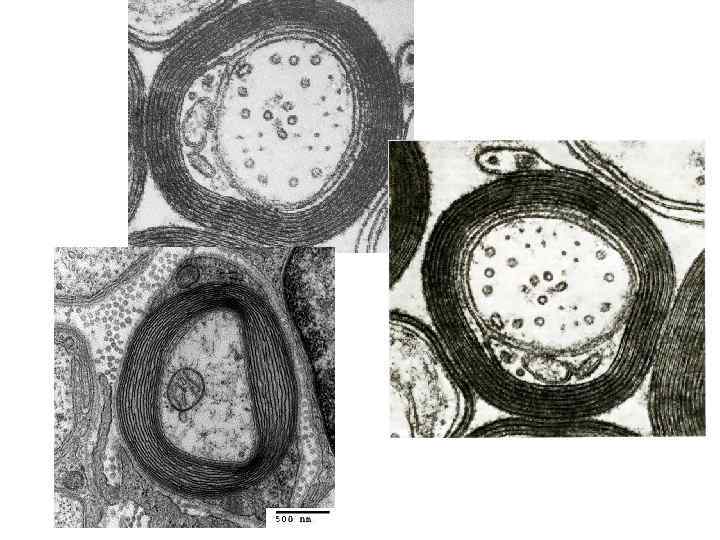
Миелиновые нервные волокна (расщепленный препарат седалищного нерва). Окраска осмиевой кислотой. Осмиевая кислота окрашивает миелиновую оболочку в чёрный цвет из-за наличия в ней липидов. На малом увеличении найти изолированное миелиновое волокно. При большом увеличении в каждом волокне виден бледно окрашенный осевой цилиндр, по бокам которого располагается тёмный миелиновый слой с узловыми перехватами и насечками, имеющих вид узких светлых косых щелей. Неврилемма при слегка опущенном конденсоре видна как блестящая полоса на периферии волокна, она особенно заметна в области узлового перехвата.

Миелиновые нервные волокна
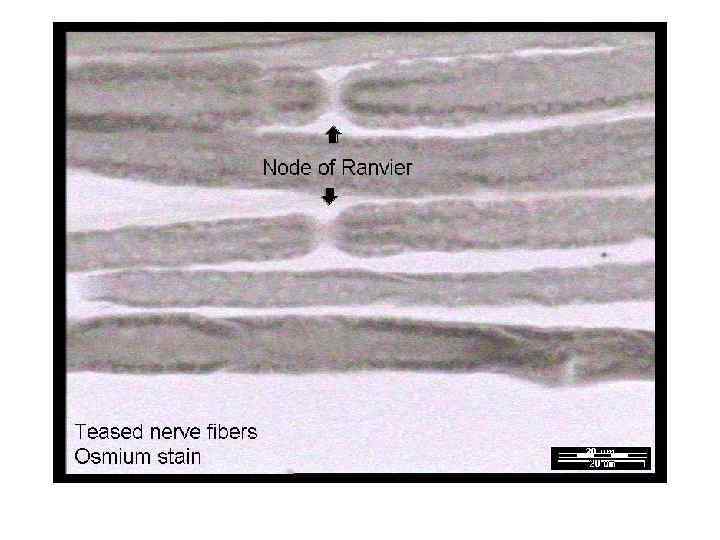
. Окр. : гем. -эозин. На малом увеличении")
Безмиелиновое нервное волокно (расщепленный препарат селезеночного нерва). Окр. : гем. -эозин. На малом увеличении найти изолированные нервные волокна. При большом увеличении они имеют вид тонких розовых тяжей, по ходу которых расположены овальной формы ядра нейролеммоцитов синефиолетового цвета. На препарате не видны оболочки нейролеммоцитов, мезаксон и осевые цилиндры, так как они очень тонкие.

Безмиелиновое нервное волокно

СКЕЛЕТНЫЕ ТКАНИ ХРЯЩЕВЫЕ ТКАНИ КОСТНЫЕ ТКАНИ

Классификация хрящевых тканей Классификация • гиалиновый, • эластический, • волокнистый – основана на структурных особенностях матрикса

• Благодаря особому строению межклеточного вещества, хрящевые ткани обладают повышенной прочностью и упругостью; эластический хрящ, кроме того, и эластичностью.

Хрящевые ткани • Главная функция – опорная: поддержка мягких тканей, формирование соединений кости (суставный гиалиновый хрящ) • Формообразующая: в раннем онтогенезе большая часть скелета первоначально формируется из хрящевой ткани. • Рост длинных костей: эпифизарные хрящевые пластинки существуют продолжительное время (до полного завершения постнатального роста костей в длину)

Клетки. • Хондроциты поддерживают нормальное состояние матрикса. • Хондроциты лежат в лакунах – полостях матрикса.

• Кровеносные и лимфатические сосуды, нервные элементы в хрящевых тканях отсутствуют. • Питание – путем диффузии питательных веществ из капилляров надхрящницы – соединительной ткани окружающей хрящевую или из синовиальной жидкости суставных сумок.

Матрикс • 75 -80% воды; • аморфное вещество, богатое протеогликанами (образуют макромолекулярные агрегаты, связывающие воду) • коллагеновые волокна

Надхрящница • За исключением суставных хрящей, гиалиновый хрящ покрыт слоем соединительной ткани - надрящницей. • Наружный волокнистый слой содержит коллаген I типа, фибробласты и многочисленные капилляры; • Внутренний клеточный (хондрогенный) слой, содержащий хондрогенные клетки и прехондробласты.

Дегенеративные изменения Гиалиновый хрящ подвержен возрастным дегенеративным изменениям. Наиболее часто – обызвествление матрикса (коллаген Х типа), гипертрофия и гибель хондроцитов. Поврежденный суставный хрящ у взрослого человека не восстанавливается, если повреждение не затрагивает кость, лежащую под ним.

Эластический хрящ Локализация: ушная раковина, стенка наружного слухового канала, евстахиева труба, надгортанник, хрящи гортани. Отличие от гиалинового хряща: Кроме коллагена II типа, матрикс содержит сеть многочисленных ветвящихся эластических волокон.

эластический хрящ • Эластический хрящ не обызвествляется, и в меньшей степени подвержен возрастным дегенеративным изменениям.

Волокнистый хрящ Занимает пограничное положение между плотной соединительной тканью и гиалиновым хрящом; сочетает в своём строении черты обеих структур. Матрикс содержит большое количество параллельно уложенных коллагеновых волокон (тип I). Хондроциты зажаты между пучками коллагеновых волокон и уложены в цепочки (изогенные группы).

Локализация волокнистого хряща • межпозвоночные диски • лобковый симфиз • мениск коленного сустава • соединения между ключицей и грудиной; между височной и нижнечелюстной костями • места перехода связок и сухожилий в кость или гиалиновый хрящ

КОСТНЫЕ ТКАНИ

Функции: • Опорно-механическая: роль опоры и рычага для прикрепленных к ним скелетных мышц • Защитная: костный скелет защищает и жизненно важные органы черепной и грудной полостей, создает нормальное окружение для кроветворения; • Гомеостатическая: мобильный резерв кальция (до 99% всего кальция в организме), фосфора и др.

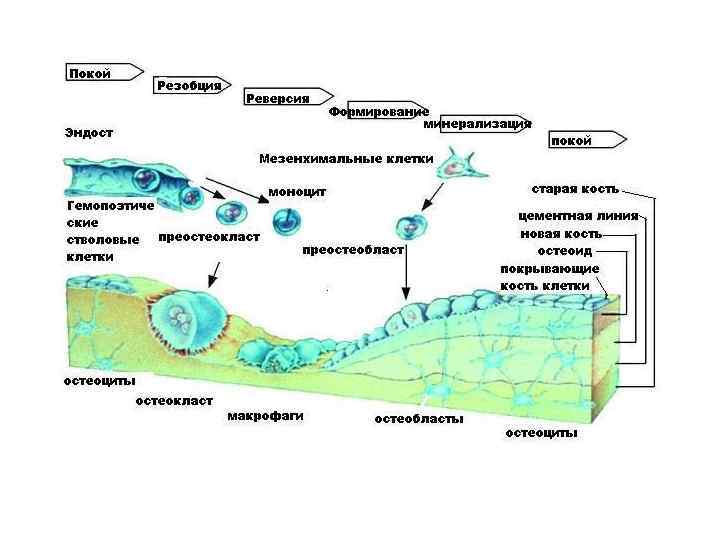
Клетки костной ткани • Клетки остеогенной дифференцировки, создающие кость: остеогенные клетки → остеобласты → остеоциты • Клетки гематогенной дифференцировки, разрушающие вещество кости (симпласты) остеокласты

остеобласты • активно синтезируют и секретируют все компоненты костного матрикса: - коллаген I типа - гликопротеины - гликозаминогликаны • органеллы белкового синтеза: - объёмная Гр. ЭС; - Гольджи; - мембранные пузырьки; - митохондрии
 многоядерные (до 50 ядер) клетки (симпласты) – образуются")
остеокласты • Гигантские (до 90 мкм) многоядерные (до 50 ядер) клетки (симпласты) – образуются в результате слияния моноцитов. Функция: • резорбция костной ткани в процессе перестройки и новообразования кости.
")
Локализация остеокластов: • На поверхности кости в небольших углублениях – лакунах Хоушипа (ниши резорбции)

Остеокласты секретируют: • Н+ ионы → кислая среда, оптимальная для растворения солей Са • гидролитические ферменты, включая коллагеназу → разрушение органического матрикса

Остеокласты чувствительны к действию гормонов: • паратиреоидный гормон → увеличивает число остеокластов и стимулирует их активность; • кальцитонин (щитовидная железа) → замедляет резорбцию кости (уменьшает протяженность гофрированной каёмки)
 • Органические вещества (25%) • Вода (25%)")
Костный матрикс • Неорганические вещества (50%) • Органические вещества (25%) • Вода (25%)

Неорганические вещества матрикса Соединения кальция и фосфора Основной компонент – гидроксиапатит. Его кристаллы при помощи белка остеонектина прикрепляются к молекулам коллагена. Ионы и молекулы воды образуют гидратированную оболочку вокруг каждого кристалла. До 30 химических элементов (Cu, F, Mg, K, Na, Zn и др. )
 Неколлагеновые")
Органический компонент • Коллаген I и Y типов (до 90% всех органических веществ) Неколлагеновые белки: • остеонектин • остеокальцин обеспечивают связывание • остеопонтин костных клеток с компонентами матрикса • Протеогликаны, ГАГ
 – первичная, или незрелая –")
Существует два основных типа костной ткани: • грубоволокнистая (ретикулофиброзная) – первичная, или незрелая – неупорядоченное расположение коллагеновых волокон в матриксе; • пластинчатая, вторичная, или зрелая, замещает грубоволокнистую в ходе нормального развития и при регенерации кости.

• Надкостница покрывает снаружи всю кость, за исключением суставных поверхностей: • наружный волокнистый слой • внутренний остеогенный • Надкостница прочно прикреплена к подлежащей костной ткани при помощи пучков коллагеновых волокон – шарпеевские волокна

• Эндост – тонкая оболочка, выстилающая кость со стороны костного мозга.

Губчатое костное вещество • Формирует внутреннюю часть эпифизов трубчатых костей и внутреннюю часть плоских костей • Состоит из переплетающихся костных пластин, между которыми остаётся значительное количество полостей, заполненных красным костным мозгом. • Пластины ориентированы в направлении наибольших нагрузок

• Формирование кости происходит благодаря слиянию трабекул друг с другом в единую сеть, промежутки которой заполнены волокнистой соединительной тканью с высоким содержанием сосудов. • Мезенхима вокруг кости даёт начало надкостнице, которая обеспечивает её питание и регенерацию.
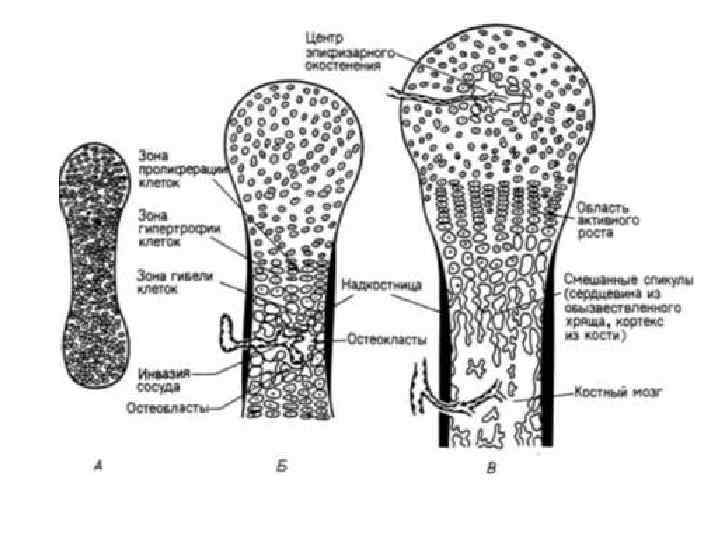

• Стадия минерализации метаэпифизарной пластинки роста В возрасте 20 -25 лет в метаэпифизарную пластинку врастают кровеносные сосуды, а с ними остеобласты, которые образуют межклеточное вещество кости. Оно минерализуется. Рост кости в длину прекращается (половые гормоны способствуют подавлению митозов в пластинке роста). Рост кости в толщину происходит за счёт надкостницы. • Стадия функциональной и возрастной перестройки кости Продолжается в течение всей жизни. Происходит постоянное разрушение старых и формирование новых остеонов, нарастание их количества и размеров при физической нагрузке и уменьшении при гипокинезии.

Ткани 61-139.ppt.pptx