

Механика и энергетика мышц.pptx
- Количество слайдов: 14
 МЕХАНИКА И ЭНЕРГЕТИКА МЫШЦ
МЕХАНИКА И ЭНЕРГЕТИКА МЫШЦ
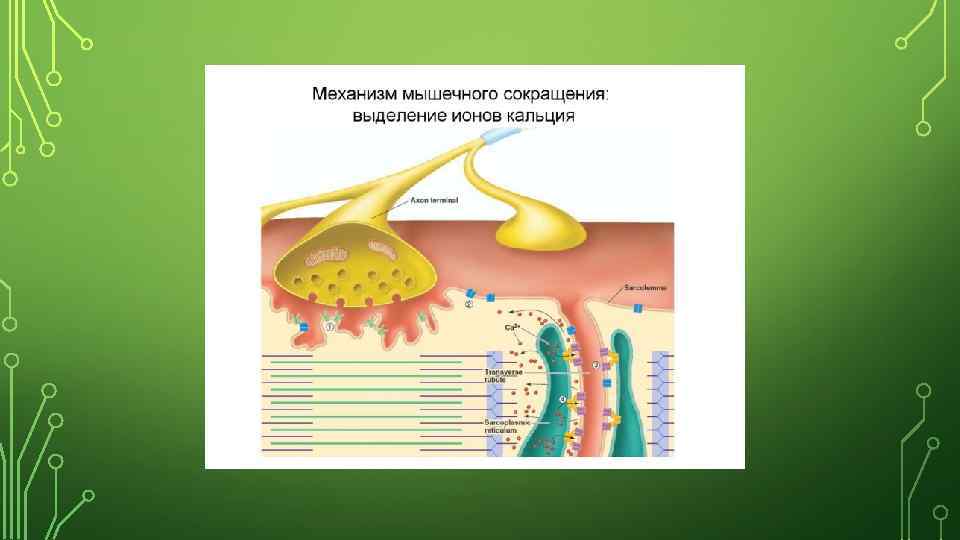
 МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ. МЫШЦА СОКРАЩАЕТСЯ В ЕСТЕСТВЕННЫХ УСЛОВИЯХ ТОЛЬКО ПРИ ПОСТУПЛЕНИИ К НЕЙ НЕРВНЫХ ИМПУЛЬСОВ. НЕРВНОЕ ВЛИЯНИЕ НА МЫШЕЧНОЕ ВОЛОКНО ПЕРЕДАЕТСЯ С ПОМОЩЬЮ НЕРВНО-МЫШЕЧНОГО СИНАПСА
МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ. МЫШЦА СОКРАЩАЕТСЯ В ЕСТЕСТВЕННЫХ УСЛОВИЯХ ТОЛЬКО ПРИ ПОСТУПЛЕНИИ К НЕЙ НЕРВНЫХ ИМПУЛЬСОВ. НЕРВНОЕ ВЛИЯНИЕ НА МЫШЕЧНОЕ ВОЛОКНО ПЕРЕДАЕТСЯ С ПОМОЩЬЮ НЕРВНО-МЫШЕЧНОГО СИНАПСА
 МЫШЕЧНОЕ СОКРАЩЕНИЕ ЯВЛЯЕТСЯ СЛОЖНЫМ МЕХАНО-ХИМИЧЕСКИМ ПРОЦЕССОМ, В ХОДЕ КОТОРОГО ПРОИСХОДИТ ПРЕОБРАЗОВАНИЕ ХИМИЧЕСКОЙ ЭНЕРГИИ ГИДРОЛИТИЧЕСКОГО РАСЩЕПЛЕНИЯ АТФ В МЕХАНИЧЕСКУЮ РАБОТУ, СОВЕРШАЕМУЮ МЫШЦЕЙ. В НАСТОЯЩЕЕ ВРЕМЯ ЭТОТ МЕХАНИЗМ ЕЩЕ ПОЛНОСТЬЮ НЕ РАСКРЫТ. НО ДОСТОВЕРНО ИЗВЕСТНО СЛЕДУЮЩЕЕ: 1. ИСТОЧНИКОМ ЭНЕРГИИ, НЕОБХОДИМОЙ ДЛЯ МЫШЕЧНОЙ РАБОТЫ ЯВЛЯЕТСЯ АТФ; 2. ГИДРОЛИЗ АТФ, СОПРОВОЖДАЮЩИЙСЯ ВЫДЕЛЕНИЕМ ЭНЕРГИИ, КАТАЛИЗИРУЕТСЯ МИОЗИНОМ, КОТОРЫЙ КАК УЖЕ ОТМЕЧАЛОСЬ, ОБЛАДАЕТ ФЕРМЕНТАТИВНОЙ АКТИВНОСТЬЮ; 3. ПУСКОВЫМ МЕХАНИЗМОМ МЫШЕЧНОГО СОКРАЩЕНИЯ ЯВЛЯЕТСЯ ПОВЫШЕНИЕ КОНЦЕНТРАЦИИ ИОНОВ СА 2+ В САРКОПЛАЗМЕ МИОЦИТОВ, ВЫЗЫВАЕМОЕ ДВИГАТЕЛЬНЫМ НЕРВНЫМ ИМПУЛЬСОМ; 4. ВО ВРЕМЯ МЫШЕЧНОГО СОКРАЩЕНИЯ МЕЖДУ ТОЛСТЫМИ И ТОНКИМИ НИТЯМИ МИОФИБРИЛЛ ВОЗНИКАЮТ ПОПЕРЕЧНЫЕ МОСТИКИ ИЛИ СПАЙКИ; 5. ВО ВРЕМЯ МЫШЕЧНОГО СОКРАЩЕНИЯ ПРОИСХОДИТ СКОЛЬЖЕНИЕ ТОНКИХ НИТЕЙ ВДОЛЬ ТОЛСТЫХ, ЧТО ПРИВОДИТ К УКОРОЧЕНИЮ МИОФИБРИЛЛ И ВСЕГО МЫШЕЧНОГО ВОЛОКНА В ЦЕЛОМ.
МЫШЕЧНОЕ СОКРАЩЕНИЕ ЯВЛЯЕТСЯ СЛОЖНЫМ МЕХАНО-ХИМИЧЕСКИМ ПРОЦЕССОМ, В ХОДЕ КОТОРОГО ПРОИСХОДИТ ПРЕОБРАЗОВАНИЕ ХИМИЧЕСКОЙ ЭНЕРГИИ ГИДРОЛИТИЧЕСКОГО РАСЩЕПЛЕНИЯ АТФ В МЕХАНИЧЕСКУЮ РАБОТУ, СОВЕРШАЕМУЮ МЫШЦЕЙ. В НАСТОЯЩЕЕ ВРЕМЯ ЭТОТ МЕХАНИЗМ ЕЩЕ ПОЛНОСТЬЮ НЕ РАСКРЫТ. НО ДОСТОВЕРНО ИЗВЕСТНО СЛЕДУЮЩЕЕ: 1. ИСТОЧНИКОМ ЭНЕРГИИ, НЕОБХОДИМОЙ ДЛЯ МЫШЕЧНОЙ РАБОТЫ ЯВЛЯЕТСЯ АТФ; 2. ГИДРОЛИЗ АТФ, СОПРОВОЖДАЮЩИЙСЯ ВЫДЕЛЕНИЕМ ЭНЕРГИИ, КАТАЛИЗИРУЕТСЯ МИОЗИНОМ, КОТОРЫЙ КАК УЖЕ ОТМЕЧАЛОСЬ, ОБЛАДАЕТ ФЕРМЕНТАТИВНОЙ АКТИВНОСТЬЮ; 3. ПУСКОВЫМ МЕХАНИЗМОМ МЫШЕЧНОГО СОКРАЩЕНИЯ ЯВЛЯЕТСЯ ПОВЫШЕНИЕ КОНЦЕНТРАЦИИ ИОНОВ СА 2+ В САРКОПЛАЗМЕ МИОЦИТОВ, ВЫЗЫВАЕМОЕ ДВИГАТЕЛЬНЫМ НЕРВНЫМ ИМПУЛЬСОМ; 4. ВО ВРЕМЯ МЫШЕЧНОГО СОКРАЩЕНИЯ МЕЖДУ ТОЛСТЫМИ И ТОНКИМИ НИТЯМИ МИОФИБРИЛЛ ВОЗНИКАЮТ ПОПЕРЕЧНЫЕ МОСТИКИ ИЛИ СПАЙКИ; 5. ВО ВРЕМЯ МЫШЕЧНОГО СОКРАЩЕНИЯ ПРОИСХОДИТ СКОЛЬЖЕНИЕ ТОНКИХ НИТЕЙ ВДОЛЬ ТОЛСТЫХ, ЧТО ПРИВОДИТ К УКОРОЧЕНИЮ МИОФИБРИЛЛ И ВСЕГО МЫШЕЧНОГО ВОЛОКНА В ЦЕЛОМ.
 МЕДИАТОРОМ В НЕРВНО-МЫШЕЧНОЙ СИНАПСЕ ЯВЛЯЕТСЯ АЦЕТИЛХОЛИН. НА ОДИН ПД ИЗ ПРЕСИНАПТИЧЕСКОГО ОКОНЧАНИЯ НЕРВНОМЫШЕЧНОГО СИНАПСА ВЫДЕЛЯЕТСЯ 200 -300 КВАНТОВ МЕДИАТОРА. В СОСТОЯНИИ ПОКОЯ, Т. Е. В ПРОМЕЖУТКАХ МЕЖДУ ПЕРЕДАЧЕЙ НЕРВНОГО ИМПУЛЬСА ПРОИСХОДИТ СПОНТАННОЕ ВЫДЕЛЕНИЕ 1 -2 КВАНТОВ МЕДИАТОРА В СИНАПТИЧЕСКУЮ ЩЕЛЬ В СРЕДНЕМ 1 РАЗ В СЕКУНДУ. ПРИ ЭТОМ НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ ФОРМИРУЕТСЯ ДЕПОЛЯРИЗАЦИЯ С АМПЛИТУДОЙ 0, 12 -0, 24 МВ. ЭТО – МИНИАТЮРНЫЕ ПОТЕНЦИАЛЫ КОНЦЕВОЙ ПЛАСТИНКИ. ПРИШЕДШИЙ ПО НЕРВНОМУ ВОЛОКНУ ИМПУЛЬС (ПД) ОБЕСПЕЧИВАЕТ ВЫДЕЛЕНИЯ В СИНАПТИЧЕСКУЮ ЩЕЛЬ АЦЕТИЛХОЛИНА, КОТОРЫЙ ВЫЗЫВАЕТ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА КОНЦЕВОЙ ПЛАСТИНКИ (ПКП) ВОЗБУЖДАЮЩЕГО ПОСТСИНАПТИЧЕСКОГО ПОТЕНЦИАЛА (ВПСП). ЭТОТ ПКП ОБЕСПЕЧИВАЕТ ПД В МЫШЕЧНОМ ВОЛОКНЕ, РАСПРОСТРАНЯЕТСЯ ВГЛУБЬ ВОЛОКНА, ЧТО ОБЕСПЕЧИВАЕТ ВЫДЕЛЕНИЕ ИОНОВ СА 2+ ИЗ САРКОПЛАЗМАТИЧЕСКОГО РЕТИКУЛУМА.
МЕДИАТОРОМ В НЕРВНО-МЫШЕЧНОЙ СИНАПСЕ ЯВЛЯЕТСЯ АЦЕТИЛХОЛИН. НА ОДИН ПД ИЗ ПРЕСИНАПТИЧЕСКОГО ОКОНЧАНИЯ НЕРВНОМЫШЕЧНОГО СИНАПСА ВЫДЕЛЯЕТСЯ 200 -300 КВАНТОВ МЕДИАТОРА. В СОСТОЯНИИ ПОКОЯ, Т. Е. В ПРОМЕЖУТКАХ МЕЖДУ ПЕРЕДАЧЕЙ НЕРВНОГО ИМПУЛЬСА ПРОИСХОДИТ СПОНТАННОЕ ВЫДЕЛЕНИЕ 1 -2 КВАНТОВ МЕДИАТОРА В СИНАПТИЧЕСКУЮ ЩЕЛЬ В СРЕДНЕМ 1 РАЗ В СЕКУНДУ. ПРИ ЭТОМ НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ ФОРМИРУЕТСЯ ДЕПОЛЯРИЗАЦИЯ С АМПЛИТУДОЙ 0, 12 -0, 24 МВ. ЭТО – МИНИАТЮРНЫЕ ПОТЕНЦИАЛЫ КОНЦЕВОЙ ПЛАСТИНКИ. ПРИШЕДШИЙ ПО НЕРВНОМУ ВОЛОКНУ ИМПУЛЬС (ПД) ОБЕСПЕЧИВАЕТ ВЫДЕЛЕНИЯ В СИНАПТИЧЕСКУЮ ЩЕЛЬ АЦЕТИЛХОЛИНА, КОТОРЫЙ ВЫЗЫВАЕТ ВОЗНИКНОВЕНИЯ ПОТЕНЦИАЛА КОНЦЕВОЙ ПЛАСТИНКИ (ПКП) ВОЗБУЖДАЮЩЕГО ПОСТСИНАПТИЧЕСКОГО ПОТЕНЦИАЛА (ВПСП). ЭТОТ ПКП ОБЕСПЕЧИВАЕТ ПД В МЫШЕЧНОМ ВОЛОКНЕ, РАСПРОСТРАНЯЕТСЯ ВГЛУБЬ ВОЛОКНА, ЧТО ОБЕСПЕЧИВАЕТ ВЫДЕЛЕНИЕ ИОНОВ СА 2+ ИЗ САРКОПЛАЗМАТИЧЕСКОГО РЕТИКУЛУМА.
 МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ. В ПРОЦЕССЕ СОКРАЩЕНИЯ МЫ ЕЧНОГО ВОЛОКНА В НЕМ ПРОИСХОДЯТ СЛЕДУЮЩИЕ ШЕЧНОГО Ш ПРЕОБРАЗОВАНИЯ: А. ЭЛЕКТРОХИМИЧЕСКОЕ ПРЕОБРАЗОВАНИЕ: 2. РАСПРОСТРАНЕНИЕ ПД ПО Т-СИСТЕМЕ. 3. ЭЛЕКТРИЧЕСКАЯ СТИМУЛЯЦИЯ ЗОНЫ КОНТАКТА Т-СИСТЕМЫ И САРКОПЛАЗМАТИЧЕСКОГО РЕТИКУЛУМА, АКТИВАЦИЯ ФЕРМЕНТОВ, ОБРАЗОВАНИЕ ИНОЗИТОЛТРИФОСФАТА, ПОВЫШЕНИЕ ВНУТРИКЛЕТОЧНОЙ КОНЦЕНТРАЦИИ ИОНОВ СА 2+. Б. ХЕМОМЕХАНИЧЕСКОЕ ПРЕОБРАЗОВАНИЕ: 4. ВЗАИМОДЕЙСТВИЕ ИОНОВ СА 2+ С ТРОПОНИНОМ, ОСВОБОЖДЕНИЕ АК ИВНЫХ ЦЕНТРОВ ТИВНЫХ Т НА АКТИНОВЫХ ФИЛАМЕНТАХ. 5. ВЗАИМОДЕЙСТВИЕ МИОЗИНОВОЙ ГОЛОВКИ С АКТИНОМ, ВРАЩЕНИЕ ГО ОВКИ И РАЗВИТИЕ ЛОВКИ Л ЭЛАСТИЧЕСКОЙ ТЯГИ. 6. СКОЛЬЖЕНИЕ НИТЕЙ АКТИНА И МИОЗИНА ОТНОСИТЕЛЬНО ДРУГА, УМЕНЬШЕНИЕ РАЗМЕРА САРКОМЕРА, РАЗВИТИЕ НАПРЯЖЕНИЯ ИЛИ УКОРО ЕНИЕ МЫШЕЧНОГО ВОЛОКНА. ЧЕНИЕ Ч
МЕХАНИЗМ МЫШЕЧНОГО СОКРАЩЕНИЯ. В ПРОЦЕССЕ СОКРАЩЕНИЯ МЫ ЕЧНОГО ВОЛОКНА В НЕМ ПРОИСХОДЯТ СЛЕДУЮЩИЕ ШЕЧНОГО Ш ПРЕОБРАЗОВАНИЯ: А. ЭЛЕКТРОХИМИЧЕСКОЕ ПРЕОБРАЗОВАНИЕ: 2. РАСПРОСТРАНЕНИЕ ПД ПО Т-СИСТЕМЕ. 3. ЭЛЕКТРИЧЕСКАЯ СТИМУЛЯЦИЯ ЗОНЫ КОНТАКТА Т-СИСТЕМЫ И САРКОПЛАЗМАТИЧЕСКОГО РЕТИКУЛУМА, АКТИВАЦИЯ ФЕРМЕНТОВ, ОБРАЗОВАНИЕ ИНОЗИТОЛТРИФОСФАТА, ПОВЫШЕНИЕ ВНУТРИКЛЕТОЧНОЙ КОНЦЕНТРАЦИИ ИОНОВ СА 2+. Б. ХЕМОМЕХАНИЧЕСКОЕ ПРЕОБРАЗОВАНИЕ: 4. ВЗАИМОДЕЙСТВИЕ ИОНОВ СА 2+ С ТРОПОНИНОМ, ОСВОБОЖДЕНИЕ АК ИВНЫХ ЦЕНТРОВ ТИВНЫХ Т НА АКТИНОВЫХ ФИЛАМЕНТАХ. 5. ВЗАИМОДЕЙСТВИЕ МИОЗИНОВОЙ ГОЛОВКИ С АКТИНОМ, ВРАЩЕНИЕ ГО ОВКИ И РАЗВИТИЕ ЛОВКИ Л ЭЛАСТИЧЕСКОЙ ТЯГИ. 6. СКОЛЬЖЕНИЕ НИТЕЙ АКТИНА И МИОЗИНА ОТНОСИТЕЛЬНО ДРУГА, УМЕНЬШЕНИЕ РАЗМЕРА САРКОМЕРА, РАЗВИТИЕ НАПРЯЖЕНИЯ ИЛИ УКОРО ЕНИЕ МЫШЕЧНОГО ВОЛОКНА. ЧЕНИЕ Ч
 ЭТАПЫ ВОЗБУЖДЕНИЯ МЫШЕЧНОГО ВОЛОКНА: • РАСПРОСТРАНЕНИЕ ПД ПО НЕРВНОМУ ВОЛОКНУ ОТ ТЕЛА МОТОНЕЙРОНА ДО СИНАПТИЧЕСКОГО ОКОНЧАНИЯ. ВОЗНИКШАЯ В РЕЗУЛЬТАТЕ ДЕПОЛЯРИЗАЦИЯ ОТКРЫВАЕТ ПОТЕНЦИАЛЗАВИСИМЫЕ СА++ КАНАЛЫ В ТЕРМИНАЛИ АКСОНА (ПРЕСИНАПТИЧЕСКАЯ МЕМБРАНА). ИОНЫ СА++ ПОСТУПАЮТ В КЛЕТКУ ПО ГРАДИЕНТУ КОНЦЕНТРАЦИИ. • ИОНЫ СА++ , ВЗАИМОДЕЙСТВУЯ СО СПЕЦИАЛЬНЫМИ БЕЛКАМИ – СИНАПСИНАМИ, АКТИВИРУЮТ ЭКЗОЦИТОЗ ГРАНУЛ С МЕДИАТОРОМ. • АЦЕТИЛХОЛИН ВЫДЕЛЯЕТСЯ В СИНАПТИЧЕСКУЮ ЩЕЛЬ И ВЗАИМОДЕЙСТВУЕТ С НИКОТИНОВЫМИ ХОЛИНОРЕЦЕПТОРАМИ (Н) НА МЕМБРАНЕ МЫШЕЧНОГО ВОЛОКНА (ПОСТСИНАПТИЧЕСКАЯ МЕМБРАНА). ИЗБЫТОК МЕДИАТОРА РАЗРУШАЕТСЯ АХ ХОЛИНЭСТЕРАЗОЙ ДО УКСУСНОЙ КИСЛОТЫ И ХОЛИНА. СВЯЗЫВАНИЕ АХ С РЕЦЕПТОРАМИ ПРИВОДИТ К ОТКРЫТИЮ КАНАЛОВ ДЛЯ ИОНОВ, В РЕЗУЛЬТАТЕ ИОН НАТРИЯ ПОСТУПАЕТ В КЛЕТКУ, ПРОИСХОДИТ ДЕПОЛЯРИЗАЦИЯ МЕМБРАНЫ И ВОЗНИКАЕТ ПОТЕНЦИАЛ КОНЦЕВОЙ ПЛАСТИНКИ (ПКП). • ПОТЕНЦИАЛ ДЕЙСТВИЯ ВОЗНИКАЕТ НА СОСЕДНЕМ УЧАСТКЕ МЕМБРАНЫ В ТОМ СЛУЧАЕ, ЕСЛИ РАЗНОСТЬ ПОТЕНЦИАЛОВ МЕЖДУ ЗАРЯДОМ МЕМБРАНЫ НА ЭТОМ УЧАСТКЕ И НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ ДОСТИГНЕТ ТАКОЙ ВЕЛИЧИНЫ, ЧТОБЫ ДЕПОЛЯРИЗОВАТЬ МЕМБРАНУ ДО КРИТИЧЕСКОГО УРОВНЯ ДЕПОЛЯРИЗАЦИИ. КАК ПРАВИЛО, В НЕРВНО-МЫШЕЧНЫХ СИНАПСАХ АМПЛИТУДА ПКП ВСЕГДА ВЫСОКА, НЕ ТРЕБУЕТ СУММАЦИИ И А ВСЕГДА ВЫЗЫВАЕТ ПД. • ПД БЫСТРО РАСПРОСТРАНЯЕТСЯ ПО ВСЕЙ МЕМБРАНЕ МЫШЕЧНОГО ВОЛОКНА.
ЭТАПЫ ВОЗБУЖДЕНИЯ МЫШЕЧНОГО ВОЛОКНА: • РАСПРОСТРАНЕНИЕ ПД ПО НЕРВНОМУ ВОЛОКНУ ОТ ТЕЛА МОТОНЕЙРОНА ДО СИНАПТИЧЕСКОГО ОКОНЧАНИЯ. ВОЗНИКШАЯ В РЕЗУЛЬТАТЕ ДЕПОЛЯРИЗАЦИЯ ОТКРЫВАЕТ ПОТЕНЦИАЛЗАВИСИМЫЕ СА++ КАНАЛЫ В ТЕРМИНАЛИ АКСОНА (ПРЕСИНАПТИЧЕСКАЯ МЕМБРАНА). ИОНЫ СА++ ПОСТУПАЮТ В КЛЕТКУ ПО ГРАДИЕНТУ КОНЦЕНТРАЦИИ. • ИОНЫ СА++ , ВЗАИМОДЕЙСТВУЯ СО СПЕЦИАЛЬНЫМИ БЕЛКАМИ – СИНАПСИНАМИ, АКТИВИРУЮТ ЭКЗОЦИТОЗ ГРАНУЛ С МЕДИАТОРОМ. • АЦЕТИЛХОЛИН ВЫДЕЛЯЕТСЯ В СИНАПТИЧЕСКУЮ ЩЕЛЬ И ВЗАИМОДЕЙСТВУЕТ С НИКОТИНОВЫМИ ХОЛИНОРЕЦЕПТОРАМИ (Н) НА МЕМБРАНЕ МЫШЕЧНОГО ВОЛОКНА (ПОСТСИНАПТИЧЕСКАЯ МЕМБРАНА). ИЗБЫТОК МЕДИАТОРА РАЗРУШАЕТСЯ АХ ХОЛИНЭСТЕРАЗОЙ ДО УКСУСНОЙ КИСЛОТЫ И ХОЛИНА. СВЯЗЫВАНИЕ АХ С РЕЦЕПТОРАМИ ПРИВОДИТ К ОТКРЫТИЮ КАНАЛОВ ДЛЯ ИОНОВ, В РЕЗУЛЬТАТЕ ИОН НАТРИЯ ПОСТУПАЕТ В КЛЕТКУ, ПРОИСХОДИТ ДЕПОЛЯРИЗАЦИЯ МЕМБРАНЫ И ВОЗНИКАЕТ ПОТЕНЦИАЛ КОНЦЕВОЙ ПЛАСТИНКИ (ПКП). • ПОТЕНЦИАЛ ДЕЙСТВИЯ ВОЗНИКАЕТ НА СОСЕДНЕМ УЧАСТКЕ МЕМБРАНЫ В ТОМ СЛУЧАЕ, ЕСЛИ РАЗНОСТЬ ПОТЕНЦИАЛОВ МЕЖДУ ЗАРЯДОМ МЕМБРАНЫ НА ЭТОМ УЧАСТКЕ И НА ПОСТСИНАПТИЧЕСКОЙ МЕМБРАНЕ ДОСТИГНЕТ ТАКОЙ ВЕЛИЧИНЫ, ЧТОБЫ ДЕПОЛЯРИЗОВАТЬ МЕМБРАНУ ДО КРИТИЧЕСКОГО УРОВНЯ ДЕПОЛЯРИЗАЦИИ. КАК ПРАВИЛО, В НЕРВНО-МЫШЕЧНЫХ СИНАПСАХ АМПЛИТУДА ПКП ВСЕГДА ВЫСОКА, НЕ ТРЕБУЕТ СУММАЦИИ И А ВСЕГДА ВЫЗЫВАЕТ ПД. • ПД БЫСТРО РАСПРОСТРАНЯЕТСЯ ПО ВСЕЙ МЕМБРАНЕ МЫШЕЧНОГО ВОЛОКНА.
 ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ СОКРАЩЕНИЕ МЫШЦЫ - ЭТО ДВИЖЕНИЯ МОСТИКОВ, КОТОРЫМ ТРЕБУЕТСЯ ЭНЕРГИЯ. ЗАПАС ЭНЕРГИИ В МЫШЦЕ ОГРАНИЧЕН, ПОЭТОМУ ПРИ РАБОТЕ МЫШЦЫ ТРЕБУЕТСЯ ПОСТОЯННОЕ ЕЕ ВОСПОЛНЕНИЕ. ИСТОЧНИКОМ ЭНЕРГИИ ДЛЯ СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ СЛУЖИТ АТФ.
ЭНЕРГЕТИКА МЫШЕЧНОГО СОКРАЩЕНИЯ СОКРАЩЕНИЕ МЫШЦЫ - ЭТО ДВИЖЕНИЯ МОСТИКОВ, КОТОРЫМ ТРЕБУЕТСЯ ЭНЕРГИЯ. ЗАПАС ЭНЕРГИИ В МЫШЦЕ ОГРАНИЧЕН, ПОЭТОМУ ПРИ РАБОТЕ МЫШЦЫ ТРЕБУЕТСЯ ПОСТОЯННОЕ ЕЕ ВОСПОЛНЕНИЕ. ИСТОЧНИКОМ ЭНЕРГИИ ДЛЯ СОКРАЩЕНИЯ И РАССЛАБЛЕНИЯ СЛУЖИТ АТФ.
 ОДНАКО ЗАПАСЫ АТФ В КЛЕТКЕ ОГРАНИЧЕНЫ. ПОЭТОМУ ДЛЯ ВОСПОЛНЕНИЯ ЗАПАСОВ АТФ ПРОИСХОДИТ ЕГО ВОССТАНОВЛЕНИЕ – РЕСИНТЕЗ.
ОДНАКО ЗАПАСЫ АТФ В КЛЕТКЕ ОГРАНИЧЕНЫ. ПОЭТОМУ ДЛЯ ВОСПОЛНЕНИЯ ЗАПАСОВ АТФ ПРОИСХОДИТ ЕГО ВОССТАНОВЛЕНИЕ – РЕСИНТЕЗ.
 МЫШЦА ИМЕЕТ ТРИ ИСТОЧНИКА ВОСПРОИЗВОДСТВА ЭНЕРГИИ: 1. РАСЩЕПЛЕНИЕ КРЕАТИНФОСФАТА. 2. АНАЭРОБНЫЙ ГЛИКОЛИЗ. 3. АЭРОБНЫЙ ГЛИКОЛИЗ.
МЫШЦА ИМЕЕТ ТРИ ИСТОЧНИКА ВОСПРОИЗВОДСТВА ЭНЕРГИИ: 1. РАСЩЕПЛЕНИЕ КРЕАТИНФОСФАТА. 2. АНАЭРОБНЫЙ ГЛИКОЛИЗ. 3. АЭРОБНЫЙ ГЛИКОЛИЗ.
 РАСЩЕПЛЕНИЕ КРЕАТИНФОСФАТА. В ХОДЕ ТАКОЙ РЕАКЦИИ, МОЛЕКУЛА КРЕАТИНФОСФАТА ОТДАЕТ СВОЮ ФОСФАТНУЮ ГРУППУ МОЛЕКУЛЕ АДЕНЕЗИНДИФОСФАТА (АДФ), В СЛЕДСТВИЕ ЧЕГО АДФ СНОВА ПРЕВРАЩАЕТСЯ В АТФ, А КРЕАТИНФОСФАТ – В КРЕАТИН. АДФ + КРЕАТИНФОСФАТ = АТФ + КРЕАТИН. ВАЖНАЯ ОСОБЕННОСТЬ ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО НА ВОССТАНОВЛЕНИЕ ЗАПАСОВ КРЕАТИНФОСФАТА НУЖНО НЕСКОЛЬКО МИНУТ И ТО, ЧТО ЭТА РЕАКЦИЯ МОЖЕТ ОСУЩЕСТВЛЯТЬСЯ ТОЛЬКО ПОСЛЕ ПРЕКРАЩЕНИЯ РАБОТЫ. ЕСЛИ БЫ КРЕАТИНФОСФАТ МОГ ВОССТАНАВЛИВАТЬСЯ ВО ВРЕМЯ РАБОТЫ, МЫ БЫ МОГЛИ ДОЛГО РАБОТАТЬ ТЯЖЕЛЫМИ ВЕСАМИ В ОЧЕНЬ БОЛЬШОМ КОЛИЧЕСТВЕ ПОВТОРЕНИЙ. ПОЭТОМУ ПОДОБНАЯ ПОДПИТКА ДЛИТСЯ ВЕСЬМА ОГРАНИЧЕННОЕ ВРЕМЯ, ПОДДЕРЖИВАЯ ЭНЕРГЕТИЧЕСКИЙ БАЛАНС МЫШЦ ЛИШЬ В САМОМ НАЧАЛЕ ИХ РАБОТЫ. СВЯЗАНО ЭТО С МАЛЫМ ЗАПАСОМ КРЕАТИНФОСФАТА В МЫШЕЧНЫХ КЛЕТКАХ. ДАЛЕЕ В РАБОТУ ВКЛЮЧАЮТСЯ АЭРОБНЫЙ И АНАЭРОБНЫЙ ГЛИКОЛИЗЫ.
РАСЩЕПЛЕНИЕ КРЕАТИНФОСФАТА. В ХОДЕ ТАКОЙ РЕАКЦИИ, МОЛЕКУЛА КРЕАТИНФОСФАТА ОТДАЕТ СВОЮ ФОСФАТНУЮ ГРУППУ МОЛЕКУЛЕ АДЕНЕЗИНДИФОСФАТА (АДФ), В СЛЕДСТВИЕ ЧЕГО АДФ СНОВА ПРЕВРАЩАЕТСЯ В АТФ, А КРЕАТИНФОСФАТ – В КРЕАТИН. АДФ + КРЕАТИНФОСФАТ = АТФ + КРЕАТИН. ВАЖНАЯ ОСОБЕННОСТЬ ЗАКЛЮЧАЕТСЯ В ТОМ, ЧТО НА ВОССТАНОВЛЕНИЕ ЗАПАСОВ КРЕАТИНФОСФАТА НУЖНО НЕСКОЛЬКО МИНУТ И ТО, ЧТО ЭТА РЕАКЦИЯ МОЖЕТ ОСУЩЕСТВЛЯТЬСЯ ТОЛЬКО ПОСЛЕ ПРЕКРАЩЕНИЯ РАБОТЫ. ЕСЛИ БЫ КРЕАТИНФОСФАТ МОГ ВОССТАНАВЛИВАТЬСЯ ВО ВРЕМЯ РАБОТЫ, МЫ БЫ МОГЛИ ДОЛГО РАБОТАТЬ ТЯЖЕЛЫМИ ВЕСАМИ В ОЧЕНЬ БОЛЬШОМ КОЛИЧЕСТВЕ ПОВТОРЕНИЙ. ПОЭТОМУ ПОДОБНАЯ ПОДПИТКА ДЛИТСЯ ВЕСЬМА ОГРАНИЧЕННОЕ ВРЕМЯ, ПОДДЕРЖИВАЯ ЭНЕРГЕТИЧЕСКИЙ БАЛАНС МЫШЦ ЛИШЬ В САМОМ НАЧАЛЕ ИХ РАБОТЫ. СВЯЗАНО ЭТО С МАЛЫМ ЗАПАСОМ КРЕАТИНФОСФАТА В МЫШЕЧНЫХ КЛЕТКАХ. ДАЛЕЕ В РАБОТУ ВКЛЮЧАЮТСЯ АЭРОБНЫЙ И АНАЭРОБНЫЙ ГЛИКОЛИЗЫ.
 НА") ГЛИКОЛИЗ - ПРОЦЕСС РАСПАДА ОДНОЙ МОЛЕКУЛЫ ГЛЮКОЗЫ (C 6 H 12 O 6) НА ДВЕ МОЛЕКУЛЫ МОЛОЧНОЙ КИСЛОТЫ (C 3 H 6 O 3) С ВЫДЕЛЕНИЕМ ЭНЕРГИИ, ДОСТАТОЧНОЙ ДЛЯ "ЗАРЯДКИ" ДВУХ МОЛЕКУЛ АТФ, ПРОТЕКАЕТ В САРКОПЛАЗМЕ ( ЦИТОПЛАЗМА МЫШЕЧНОЙ КЛЕТКИ ) ПОД ВОЗДЕЙСТВИЕМ 10 СПЕЦИАЛЬНЫХ ФЕРМЕНТОВ.
ГЛИКОЛИЗ - ПРОЦЕСС РАСПАДА ОДНОЙ МОЛЕКУЛЫ ГЛЮКОЗЫ (C 6 H 12 O 6) НА ДВЕ МОЛЕКУЛЫ МОЛОЧНОЙ КИСЛОТЫ (C 3 H 6 O 3) С ВЫДЕЛЕНИЕМ ЭНЕРГИИ, ДОСТАТОЧНОЙ ДЛЯ "ЗАРЯДКИ" ДВУХ МОЛЕКУЛ АТФ, ПРОТЕКАЕТ В САРКОПЛАЗМЕ ( ЦИТОПЛАЗМА МЫШЕЧНОЙ КЛЕТКИ ) ПОД ВОЗДЕЙСТВИЕМ 10 СПЕЦИАЛЬНЫХ ФЕРМЕНТОВ.
 И С ПОТРЕБЛЕНИЕМ КИСЛОРОДА") ГЛИКОЛИЗ МОЖЕТ ПРОТЕКАТЬ БЕЗ ПОТРЕБЛЕНИЯ КИСЛОРОДА (ТАКИЕ ПРОЦЕССЫ НАЗЫВАЮТСЯ АНАЭРОБНЫМИ) И С ПОТРЕБЛЕНИЕМ КИСЛОРОДА (АЭРОБНЫЙ ГЛИКОЛИЗ) СПОСОБЕН БЫСТРО ВОССТАНАВЛИВАТЬ ЗАПАСЫ АТФ В МЫШЦЕ.
ГЛИКОЛИЗ МОЖЕТ ПРОТЕКАТЬ БЕЗ ПОТРЕБЛЕНИЯ КИСЛОРОДА (ТАКИЕ ПРОЦЕССЫ НАЗЫВАЮТСЯ АНАЭРОБНЫМИ) И С ПОТРЕБЛЕНИЕМ КИСЛОРОДА (АЭРОБНЫЙ ГЛИКОЛИЗ) СПОСОБЕН БЫСТРО ВОССТАНАВЛИВАТЬ ЗАПАСЫ АТФ В МЫШЦЕ.
 СХЕМА РЕСИНТЕЗА АТФ
СХЕМА РЕСИНТЕЗА АТФ