07_Получение энергии из глюкозы.ppt
- Количество слайдов: 62



Метаболизм Получение энергии из пищи I. Образование энергии из глюкозы

Основные этапы окисления глюкозы

Окисление глюкозы • Суммарное уравнение С 6 Н 12 О 6 + 6 О 2 → 6 СО 2 + 6 Н 2 О G = – 2820 к. Дж/моль • 3 этапа: 1. Гликолиз (в цитоплазме) 2. Цикл Кребса (в митохондриях) 3. Дыхательная цепь (на внутренней мембране митохондрий)

Дихотомический путь распада глюкозы. Гликолиз

Гликолиз… • сложный ферментативный процесс превращения глюкозы, протекающий в тканях человека и животных без потребления кислорода • На промежуточных стадиях образуются трехуглеродные фрагменты, используемые для биосинтеза ряда веществ • У ряда микроорганизмов процессами, аналогичными гликолизу, являются различные виды брожения • 11 ферментов • В гиалоплазме клеток

1. Фосфорилирование G° = – 16, 74 к. Дж/моль • Гексокиназа не обладает абсолютной специфичностью • В печени – глюкокиназа, катализирующая фосфорилирование только D-глюкозы • Первая пусковая реакция гликолиза

2. Изомеризация Г-6 -Ф во Ф-6 -Ф G° = 1, 67 к. Дж/моль • Гексозофосфатизомераза (Г-6 -Ф-изомераза) • Гексозофосфатизомераза активируется субстратом

3. Фосфорилирование Ф-6 -Ф G° = – 14, 23 к. Дж/моль • • 6 -фосфофруктокиназа Вторая пусковая реакция гликолиза Лимитирующая стадия Фосфофруктокиназа ингибируется АТФ и стимулируется АДФ и АМФ

4. Распад Ф-1, 6 -д. Ф на 2 фосфотриозы G° = 23, 85 к. Дж/моль • Альдолаза

5. Изомеризация триозофосфатов G° = 7, 53 к. Дж/моль • Триозофосфатизомераза • Равновесие сдвинуто в сторону ДОАФ (его 95%) • НО в последующие реакции может включаться только 3 -ФГА, поэтому по мере потребления 3 -ФГА ДОАФ превращается в 3 -ФГА • Образованием 3 -ФГА завершается первая стадия гликолиза

Биологическое окисление – … совокупность окислительных реакций, происходящих в биологических объектах и обеспечивающих их энергией и метаболитами для осуществления процессов жизнедеятельности

Типы биологического окисления 1. Свободное окисление 2. Окисление, сопряженное с фосфорилированием АДФ Ø субстратное фосфорилирование Ø окислительное

Субстратное фосфорилирование – … вид биологического окисления, при котором: ü макроэргическая связь возникает в момент непосредственного окисления субстрата ü затем передается на фосфатный остаток, ü который используется для фосфорилирования АДФ – синтеза АТФ Окисление, сопряженное с фосфорилированием АДФ на уровне субстрата

6. Окисление 3 -ФГА до 1, 3 -д. ФГК G° = 6, 28 к. Дж/моль • Глицеральдегид-3 -фосфатдегидрогеназа (кофермент – НАД)
–СН")
6. Окисление 3 -ФГА до 1, 3 -д. ФГК • Механизм: Е–SН + ОНС–СН(ОН)–СН 2–О–РО(ОН)2 + НАД ⇄ ⇄ Е–S СО–СН(ОН)–СН 2–О–РО(ОН)2 + НАДН 2; I Е–S СО–СН(ОН)–СН 2–О–РО(ОН)2 + Н 3 РО 4 ⇄ ⇄ Е–SН + + (ОН)2 РО О–СО–СН(ОН)–СН 2–О–РО(ОН)2 II

7. Субстратное фосфорилирование III G° = – 18, 83 к. Дж/моль • Фосфоглицераткиназа • Таким образом, энергия, высвобождающаяся при окислении альдегидной группы 3 -ФГА, запасается в форме энергии АТФ

8. Изомеризация 3 -ФГК в 2 -ФГК G° = 4, 6 к. Дж/моль • фосфоглицерат-фосфомутаза • легко обратима, протекает в присутствии Mg 2+

9. Дегидратация 2 -ФГК G° = 1, 67 к. Дж/моль • Енолаза, активируется Mg 2+ или Mn 2+ и ингибируется F–

10. Субстратное фосфорилирование G° = – 31, 38 к. Дж/моль • пируваткиназа; необходимы Mg 2+ и Mn 2+, а также ионы щелочных металлов (К+ или др. )

Схема гликолиза

Суммарное уравнение гликолиза С 6 Н 12 О 6 + 2 НАД + 2 АДФ + 2 Н 3 РО 4 → → 2 СН 3–СО–СООН + 2 НАДН 2 + 2 АТФ 6 АТФ 8 АТФ

Биологическое значение гликолиза • образование макроэргических соединений • Энергетическая эффективность гликолиза – 2 молекулы АТФ на 1 молекулу глюкозы • Если гликолиз начинается не с глюкозы, а с гликогена или крахмала (гликогенолиз), то накапливаются не 2, а 3 молекулы АТФ (? ) • Однако в энергетическом плане гликолиз и гликогенолиз практически равноценны, т. к. на синтез гликогена в тканях расходуется АТФ

Обмен пировиноградной кислоты

1. В анаэробных условиях – восстановление до молочной кислоты • Лактатдегидрогеназа • Другие названия процесса – – молочнокислое брожение – окончание гликолиза • (в зависимости от локализации)

• Молочная кислота накапливается при активной мышечной работе и вызывает утомление мышц • Часть ее (1/5 – 1/6) в аэробных условиях окисляется до СО 2 и Н 2 О или идет на ресинтез гликогена • G° 310 расщепления глюкозы до молочной кислоты ~210 к. Дж/моль • ~126 к. Дж – в виде тепла, а ~84 к. Дж – в форме АТФ • Концевая макроэргическая связь АТФ соответствует ~33, 6– 42, 0 к. Дж/моль • Таким образом, КПД гликолиза (до молочной кислоты) составляет около 0, 4

2. В анаэробных условиях – спиртовое брожение • Дрожжеподобные организмы и некоторые плесневые грибы С 6 Н 12 О 6 → 2 СО 2 + 2 С 2 Н 5 ОН • По механизму близко к гликолизу • пируватдекарбоксилаза (Mg 2+ и кофермент тиаминпирофосфат) СН 3–СО–СООН → СН 3–СНО + СО 2 • алкогольдегидрогеназа СН 3–СНО + НАДН 2 → СН 3–СН 2–ОН + НАД • Другие виды брожения отличаются конечными продуктами
 • спиртовое брожение")
Анаэробные условия • восстановление до молочной кислоты (молочнокислое брожение, окончание гликолиза) • спиртовое брожение • другие виды брожения
 • Окислительное декарбоксилирование с образованием ацетил-Ко.")
3. В аэробных условиях – окислительное декарбокислирование (дыхание) • Окислительное декарбоксилирование с образованием ацетил-Ко. А, который затем может окисляться до СО 2 и Н 2 О СН 3–СО–СООН + НАД + НSКо. А → → СО 2 + СН 3–СО SКо. А + НАДН 2 • Пируватдегидрогеназный комплекс – мультиэнзимный комплекс

Модель дегидрогеназного полиферментного комплекса 12 димерных молекул пируватдекарбоксилазы 6 димерных молекул дигидролипоилдегидрогеназы 24 молекулы липоатацетилтрансферазы

Механизм окислительного декарбоксилирования ПВК
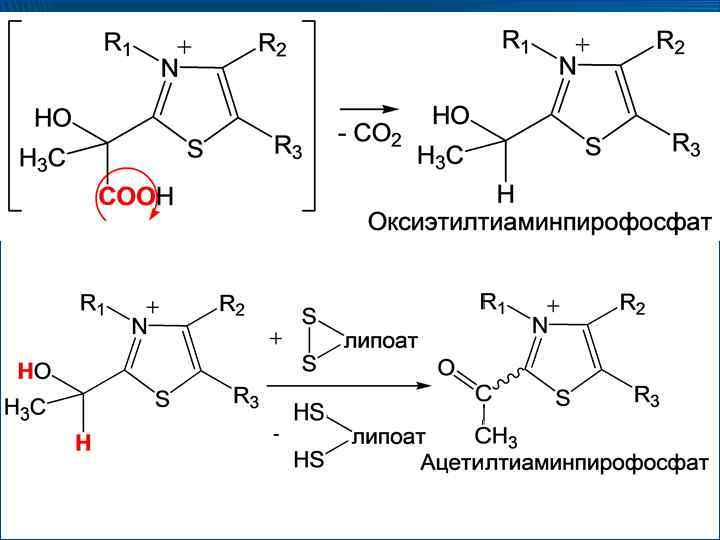
2–липоат + ФАД → S 2–липоат + ФАДН 2; ФАДН 2 + НАД →")
(HS)2–липоат + ФАД → S 2–липоат + ФАДН 2; ФАДН 2 + НАД → НАДН 2 + ФАД

Механизм действия пируватдегидрогеназного комплекса ПВК ТПФ СО 2 Оксиэтил. ТПФ Липоат ФАДН 2 НАД ФАД НАДН 2 Дигидролипоат Ацетилгидролипоат Ко. А Ацетил. Ко. А
")
Цикл трии дикарбоновых кислот (цикл Кребса)

• Х. А. Кребс, 1937 г. • впервые постулировал значение цикла для полного «сгорания» ПВК, главным источником которой является гликолиз • В дальнейшем было показано, что цикл Кребса – «фокус» , в котором сходятся практически все метаболические пути • 8 последовательных реакций • Ферменты цикла собраны в метаболон, локализованный в кристах митохондрий Ханс Адольф Кребс • Рядом с метаболоном цикла Кребса располагаются (1900– 1981) пируватдегидрогеназный комплекс и, вероятно, метаболон -окисления жирных кислот, поставляющие ему ацетил-Ко. А

1. Конденсация ацетил-Ко. А со ЩУК • Цитратсинтаза • Несколько стадий
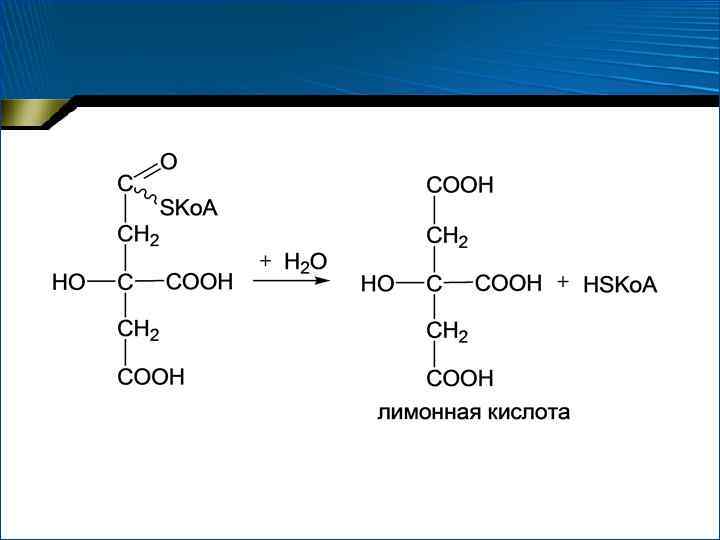

• Суммарное уравнение 1 стадии: G° = – 31, 38 к. Дж/моль

2. Изомеризация лимонной кислоты в изолимонную • Аконитатгидратаза • Две стадии – 2. 1. Дегидратация лимонной кислоты G° = 8, 37 к. Дж/моль

• Аконитатгидратаза обладает стереохимической специфичностью, обусловленной симметричностью субстрата

– 2. 2. Гидратация цис-аконитовой кислоты: G° = – 2, 09 к. Дж/моль
 • G° =")
3. Окисление изолимонной кислоты, сопровождающееся декарбоксилированием • Изоцитратдегидрогеназа (кофермент – НАД) • G° = – 8, 37 к. Дж/моль

4. Окислительное декарбоксилирование -кетоглутаровой кислоты • -Кетоглутаратдегидрогеназа Сукцинил-Ко. А G° = – 30, 12 к. Дж/моль

5. Субстратное фосфорилирование • Сукцинат: Ко. А-лигаза Название фермента сукцинаттиокиназа устарело! G° = – 3, 35 к. Дж/моль ГТФ + АДФ ⇄ ГДФ + АТФ
 • Сукцинатдегидрогеназа (кофермент – ФАД): G° =")
6. Превращение янтарной кислоты в фумаровую (дегидрирование) • Сукцинатдегидрогеназа (кофермент – ФАД): G° = 0

7. Гидратация фумаровой кислоты с образованием яблочной кислоты • Фумаратгидратаза G° = – 3, 77 к. Дж/моль

8. Окисление яблочной кислоты до кетоформы ЩУК • Малатдегирогеназа G° = 29, 71 к. Дж/моль

Суммарное уравнение цикла Кребса СН 3–СО SКо. А + 3 НАД + ФАД + АДФ + Н 3 РО 4 → → НSКо. А + 3 НАДН 2 + ФАДН 2 + АТФ + 2 СО 2

• За 1 оборот цикла Кребса – полное окисление 1 молекулы ацетил-Ко. А • Для непрерывной работы цикла необходимо – постоянное поступление в систему ацетил-Ко. А – НАДН 2 и ФАДН 2 должны снова и снова окисляться в дыхательной цепи • Освобождающаяся энергия в значительной мере сосредоточивается в макроэргических связях АТФ • При окислении 1 молекулы ацетил-Ко. А в цикле Кребса – 12 молекул АТФ

ОКИСЛИТЕЛЬНОЕ ФОСФОРИЛИРОВАНИЕ. Дыхательная цепь

Окислительное фосфорилирование… Ø сопряжение окисления с синтезом АТФ, Ø когда атомы водорода с коферментов дегидрогеназ, принимающих участие в окислении субстратов, передаются в оксидоредуктазную цепь, Ø где сопряженно с переносом ионов Н+ и электронов на молекулярный кислород происходит активирование неорганического фосфата и при его посредстве – фосфорилирование АДФ с образованием АТФ Ø Окисляемый субстрат непосредственного участия в активировании неорганического фосфата не принимает Ø Сопряжение окисления с фосфорилированием идет главным образом на внутренних мембранах митохондрий
")
ОКИСЛИТЕЛЬНОВОССТАНОВИТЕЛЬНЫЕ ФЕРМЕНТЫ (повторение)

Пиридиновые дегидрогеназы Ø Коферменты – НАД и НАДФ Ø Универсальный донор атомов Н для дыхательной цепи ферментов – НАДН 2 НАДФН 2 + НАД ⇄ НАДФ + НАДН 2

Флавиновые дегидрогеназы Ø Коферменты – ФМН и ФАД Ø являются акцепторами атомов Н и осуществляют перенос их от НАДН 2 + ФАД ⇄ НАД + ФАДН 2 Ø В некоторых случаях (? ) могут играть роль первичных дегидрогеназ
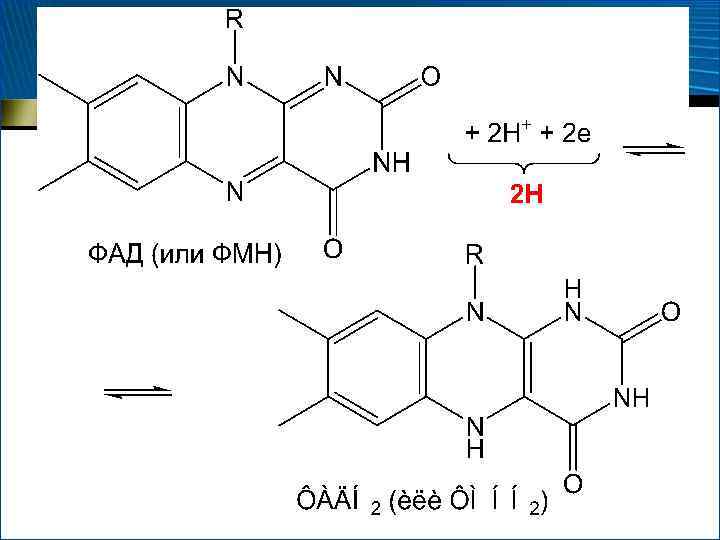
")
Кофермент Q (убихинон)

Цитохромы Ø Дальнейший перенос электронов от Ко. Q на О 2 осуществляет система цитохромов – ряд гемопротеидов, расположенных в порядке возрастания ОВП (? ) Ø Цит а и а 3 содержат в своем составе атомы Cu Ø При транспорте электронов в направлении увеличения ОВП происходят процессы: Fe 2+ ⇄ Fe 3+ (в цит. b, c 1, с) Cu+ ⇄ Cu 2+ (в цит. а, а 3)

Схема дыхательной цепи

Ø Самая примечательная особенность дыхательной цепи – наличие участков, где соседние компоненты резко отличаются значениями ОВП Ø Именно здесь происходит сопряжение окисления с фосфорилированием АДФ

Хемиосмотическая гипотеза Питер Митчелл, Владимир Петрович Скулачев Ø Реакции, сопровождающиеся расходованием или образованием Н+, протекают на внутренней мембране митохондрий так , что Н+ переносятся с внутренней мембраны на внешнюю, т. е. перенос электронов сопровождается возникновением трансмембранного градиента концентрации Н+ – совершением осмотической работы Ø Этот градиент, создающий разность химических ( ) и электрических ( ) потенциалов, – источник энергии образования АТФ Ø АТФаза – фермент, способный использовать градиент концентрации ионов Н+ для обращения процесса гидролиза АТФ

Энергетический эффект гликолитического расщепления 1 молекулы глюкозы и последующего окисления ПВК до СО 2 и Н 2 О

ГЛИКОЛИЗ Глюкоза 2 ПВК 2 НАДН 2 6 АТФ ЦИКЛ КРЕБСА 2 ацетил-Ко. А СО 2 + Н 2 О 2 НАДН 2 2 • 3 НАДН 2 2 ФАДН 2 2 АТФ 6 АТФ 18 АТФ 4 АТФ 2 АТФ лактат На самом деле – 3638 АТФ 2 АТФ тратятся на АТФ т. к. перенос НАДН 2 из цитоплазмы в митохондрию
07_Получение энергии из глюкозы.ppt