Гормоны.ppt
- Количество слайдов: 90


; •")
Мембраны: • отделяют клетки от окружающей среды и делят ее на компартменты (отсеки); • регулируют транспорт веществ в клетки и органеллы и в обратном направлении; • обеспечивают специфику межклеточных контактов; • воспринимают сигналы из внешней среды.

Биологические мембраны построены из липидов и белков, связанных друг с другом с помощью нековалентных взаимодействий. Основу мембраны составляет двойной липидный слой, в состав которого включены белковые молекулы. Липидный бислой образован двумя рядами амфифильных молекул, гидрофобные «хвосты» которых спрятаны внутрь, а гидрофильные группы - полярные «головки» обращены наружу и контактируют с водной средой. Поперечный разрез плазматической мембраны

Липиды мембран. В состав липидов мембран входят как насыщенные, так и ненасыщенные жирные кислоты. Ненасыщенные жирные кислоты встречаются в два раза чаще чем насыщенные, что определяет текучесть мембран и конформационную лабильность мембранных белков. Некоторые мембранные белки перемещаются вдоль бислоя (латеральная диффузия) или поворачиваются вокруг оси, перпендикулярно его поверхности. Например, фермент фосфолипаза А 2, связываясь с цитоплазматической поверхностью мембраны, может латерально перемещаться по поверхности бислоя и гидролизовать несколько тысяч фосфолипидов в минуту до тех пор, пока не отделится от мембраны. В мембранах присутствуют липиды трех главных типов - фосфолипиды, гликолипиды и холестерол. Чаще всего встречаются глицерофосфолипиды - производные фосфатидной кислоты.

Фосфатидная кислота - это диацилглицеролфосфат. R 1, R 2 - радикалы жирных кислот (гидрофобные «хвосты» ). Со вторым углеродным атомом глицерола связан остаток полиненасыщенной жирной кислоты. Полярной «головкой» является остаток фосфорной кислоты и присоединенная к нему гидрофильная группа серина, холина, этаноламина или инозитола Глицерофосфолипиды
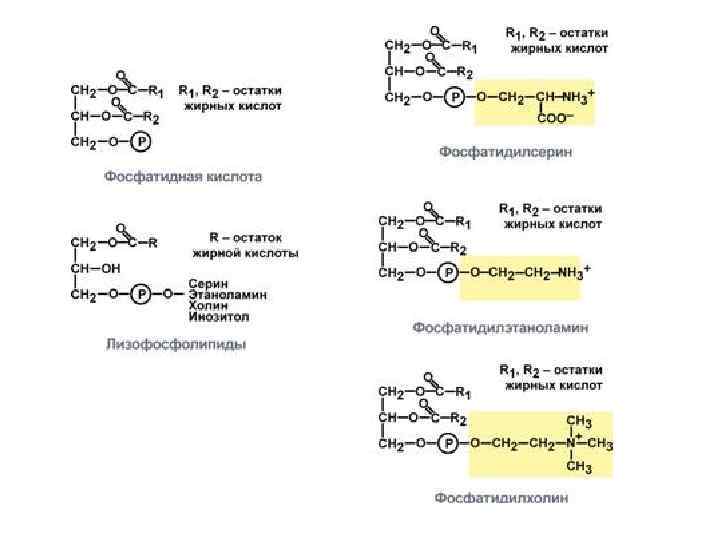

Существуют также липиды - производные аминоспирта сфингозина. Аминоспирт сфингозин при ацилировании, т. е. присоединении жирной кислоты к NH 2 -группе, превращается в церамид. Церамиды различаются по остатку жирной кислоты. С ОН-группой церамида могут быть связаны разные полярные группы. В зависимости от строения полярной «головки» эти производные разделены на две группы - фосфолипиды и гликолипиды. Строение полярной группы сфингофосфолипидов (сфингомиелинов) сходно с глицерофосфолипидами. Много сфингомиелинов содержится в составе миелиновых оболочек нервных волокон. Гликолипиды представляют собой углеводные производные церамида. В зависимости от строения углеводной составляющей различают цереброзиды и ганглиозиды (см. след слайд).
. К фосфолипидам относятся сфингомиелины,")
Церамид - ацилированный сфингозин (R 1 - радикал жирной кислоты). К фосфолипидам относятся сфингомиелины, у которых полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Гидрофильной группой (полярной «головкой» ) гликолипидов является углеводный остаток. Цереброзиды содержат моно- или олигосахаридный остаток линейного строения. В состав ганглиозидов входит разветвленный олигосахарид, одним из мономерных звеньев которого является НАНК - Nацетилнейраминовая кислота Производные аминоспирта сфингозина.
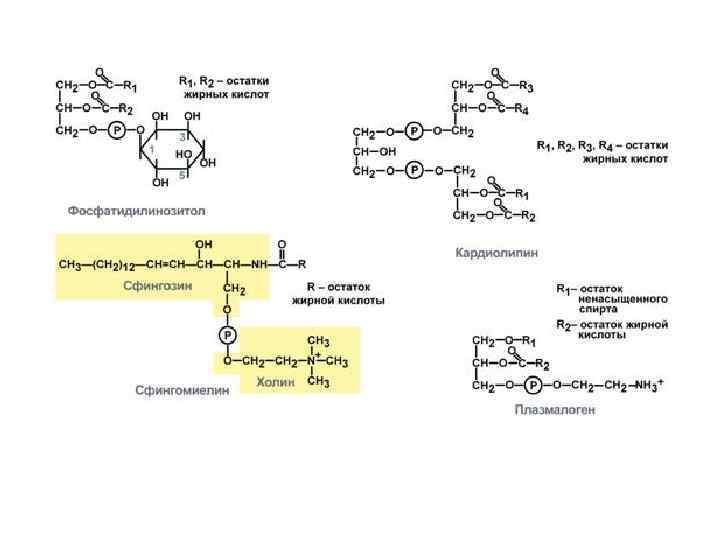

Холестерол содержится в мембранах всех животных клеток, он придает мембранам жесткость и снижает их жидкостность (текучесть). Молекула холестерола располагается в гидрофобной зоне мембраны параллельно гидрофобным «хвостам» молекул фосфо- и гликолипидов. Гидроксильная группа холестерола, как и гидрофильные «головки» фосфо- и гликолипидов, обращена к водной фазе. Молярное соотношение холестерола и других липидов в мембранах равно 0, 3 -0, 9. Самое высокое значение имеет эта величина для цитоплазматической мембраны. Увеличение содержания холестерола в мембранах уменьшает подвижность цепей жирных кислот, что влияет на конформационную лабильность мембранных белков и снижает возможность их латеральной диффузии.

Молекула холестерола состоит из жесткого гидрофобного ядра и гибкой углеводородной цепи. Полярной «головкой» является ОН-группа у 3 -го углеродного атома молекулы холестерола. Для сравнения на рисунке представлено схематическое изображение фосфолипида мембран. Полярная головка этих молекул значительно больше и имеет заряд Положение в мембране фосфолипидов и холестерола.

Главные функции липидов мембран состоят в том, что они: • формируют липидный бислой - структурную основу мембран; • обеспечивают необходимую для функционирования мембранных белков среду; • участвуют в регуляции активности ферментов; • служат «якорем» для поверхностных белков; • участвуют в передаче гормональных сигналов. Изменение структуры липидного бислоя может привести к нарушению функций мембран.
 белки; 3, 4, 5, 6 - поверхностные")
Белки мембран: 1, 2 - интегральные (трансмембранные) белки; 3, 4, 5, 6 - поверхностные белки. В интегральных белках часть полипептидной цепи погружена в липидный слой. Те участки белка, которые взаимодействуют с углеводородными цепями жирных кислот, содержат преимущественно неполярные аминокислоты. Участки белка, находящиеся в области полярных «головок» , обогащены гидрофильными аминокислотными остатками. Поверхностные белки разными способами прикрепляются к мембране: 3 - связанные с интегральными белками; 4 - присоединенные к полярным «головкам» липидного слоя; 5 - «заякоренные» в мембране с помощью короткого гидрофобного концевого домена; 6 - «заякоренные» в мембране с помощью ковалентно связанного ацильного остатка

Для нормального функционирования многоклеточного организма необходима взаимосвязь между отдельными клетками, тканями и органами. Эту взаимосвязь осуществляют 4 основные системы регуляции: • Центральная и периферическая нервная системы (через нервные импульсы и нейромедиаторы) • Эндокринная система (через гормоны, которые секретируются в кровь) • Паракринная и аутокринная системы (через гормоны, которые секретируются в межклеточное пространство) • Иммунная система (через специфические белки: цитокины и антитела)

ГОРМОНЫ

Гормоны - биологически активные органические соединения, которые вырабатываются в незначительных количествах в железах внутренней секреции, осуществляют гуморальную регуляцию обмена веществ и имеют различную химическую структуру. Признаки гормонов: - Дистантность действия – синтез в железах внутренней секреции, а регуляция отдаленных тканей - Строгая специфичность действия – даже очень близкие по химической структуре аналоги гормонов не дают нужного биологического эффекта (принцип структурной комплементарности) - Кратковременность действия - Действуют в очень низких концентрациях 10 -12 -10 -6 моль/л, под контролем ЦНС и регуляция их действия осуществляется в большинстве случаев по типу обратной связи - Действуют опосредованно через белковые рецепторы (R) и ферментативные системы, что запускает каскадный механизм усиления гормонального сигнала -Действие направлено на изменение скорости синтеза ферментов или их активности (итог действия гормона – изменение количества определенного метаболита)

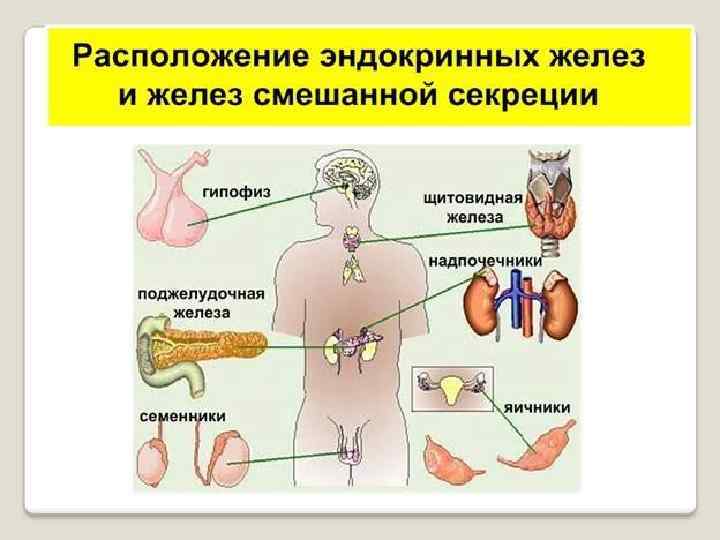
Гормоны можно классифицировать в зависимости от места их природного синтеза, в соответствии с которым различают гормоны гипоталамуса, гипофиза, щитовидной железы, надпочечников, поджелудочной железы, половых желез и др. Однако подобная анатомическая классификация недостаточно совершенна, поскольку некоторые гормоны или синтезируются не в тех железах внутренней секреции, из которых они секретируются в кровь (например, гормоны задней доли гипофиза, вазопрессин и окситоцин синтезируются в гипоталамусе, откуда переносятся в заднюю долю гипофиза), или синтезируются и в других железах (например, частичный синтез половых гормонов осуществляется в коре надпочечников, синтез простагландинов происходит не только в предстательной железе, но и в других органах, соматостатин синтезируется как в гипоталямусе, так и в поджелудочной железе) и т. д. С учетом этих обстоятельств были предприняты попытки создания современной классификации гормонов, основанной на их химической природе. В соответствии с этой классификацией различают три группы истинных гормонов:
 ГОРМОН ПЕПТИДНОЙ или БЕЛКОВОЙ ПРИРОДЫ Адренокортикотропный")
Классификация гормонов по химическому строению (не указаны эйкозаноиды) ГОРМОН ПЕПТИДНОЙ или БЕЛКОВОЙ ПРИРОДЫ Адренокортикотропный гормон (кортикотропин; АКТГ) Гормон роста (соматотропин; СТГ) Тиротропный гормон (тиротропин; ТТГ) Лактогенный гормон (пролактин; ЛТГ) Лютеинизирующий гормон (лютропин; ЛГ) Фолликул ости мул ирующий гормон (ФСГ) Мепаноцитстимулирующий гормон (МСГ) Хорионический гонадотропин (ХГ) Антидиуретический гормон (вазопрессин; АДГ) Окситоцин Паратироидный гормон (паратгормон; ПТГ) Кальцитонин Инсулин Глюкагон СТЕРОИД Альдостерон Кортизоп Кальцитриоп Тестостерон Эстрадиол Прогестерон ПРОИЗВОДНОЕ АМИНОКИСЛОТЫ Адреналин Норадреналин Трийодтиронин (Т 3) Тироксин (Т 4)

Взаимосвязь нервной и эндокринной систем регуляции Клетки центральной нервной системы воспринимают сигналы из внешней и внутренней среды (1). По нейронам нервных клеток сигналы поступают в гипоталамус, где синтезируются пептиды - рилизинг-гормоны: либерины или статины (2). Они регулируют синтез и секрецию гормонов передней доли гипофиза (тропных гормонов), причем либерины стимулируют, а статины тормозят эти процессы (3). Тропные гормоны являются гликопротеинами. Они секретируются в кровь и транспортируются к эндокринным железам, стимулируя синтез соответствующих гормонов в эндокринных периферических железах (4). Эти гормоны поступают в кровоток, взаимодействуют с белками-рецепторами клеток-мишеней и, изменяя скорость транспорта веществ в клетки, активность или количество специфических белков и ферментов, влияют на метаболизм (5). Изменение концентрации метаболитов в клеткахмишенях по механизму отрицательной обратной связи подавляет синтез гормонов эндокринных желез (6). Синтез и секреция тропных гормонов подавляется гормонами эндокринных желез (7). (+) – стимуляция синтеза и секреции гормонов, (—) – подавление синтеза и секреции гормонов (отрицательная обратная связь)
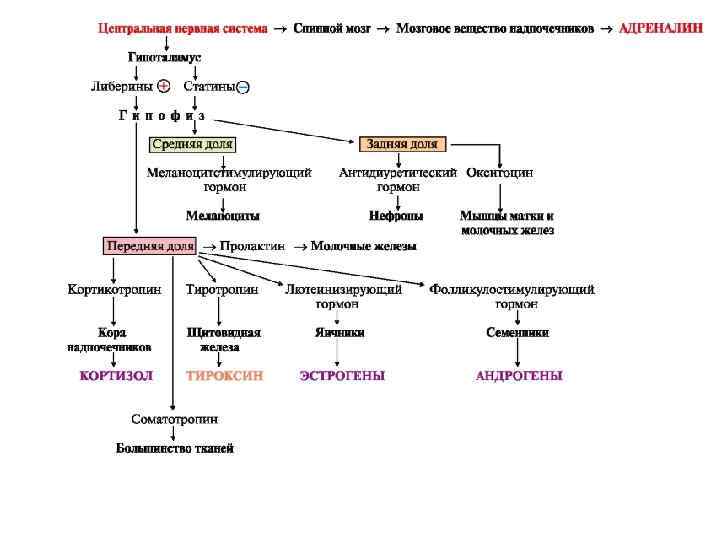
 и окситоцин - представляют собой пептиды,")
Гормоны задней доли гипофиза - антидиуретический гормон (вазопрессин) и окситоцин - представляют собой пептиды, которые синтезируются в гипоталамусе, поступают в гипофиз и оттуда секретируются в кровь. Синтез гормонов в организме регулируется по механизму отрицательной обратной связи. Под их влиянием изменяются скорости метаболических путей в тканях-мишенях и, следовательно, количество образующихся в них метаболитов. Это является сигналом для торможения синтеза гормонов гипоталамуса, гипофиза и эндокринных желез. Мозговое вещество надпочечников, в хромаффинных клетках которого синтезируется гормон адреналин, находится под непосредственным контролем центральной нервной системы. Секреция гормонов поджелудочной железы - инсулина и глюкагона - определяется уровнем глюкозы в крови.

В зависимости от биологических функций, которые регулируют гормоны, выделяют группы, контролирующие: • обмен углеводов, липидов, аминокислот (инсулин, глюкагон, адреналин, кортизол, соматотропин, тироксин); • водно-солевой обмен (альдостерон, антидиуретический гормон, предсердный натриуретический фактор); • обмен кальция и фосфатов (паратгормон, кальцитриол, кальцитонин); • репродуктивную функцию (гонадотропные гормоны, эстрогены, тестостерон); • функции эндокринных желез (тропные гормоны гипофиза).

Примеры Пептидные гормоны синтезируются, как и другие белки, в процессе трансляции из аминокислот. Некоторые пептидные гормоны - это короткие пептиды; например, гормон гипоталамуса тиреотропин-либерин — трипептид. Большинство гормонов передней доли гипофиза — гликопротеины. Некоторые пептидные гормоны являются продуктами общего гена. Большинство полипептидных гормонов синтезируется в виде неактивных предшественников - препрогормонов. Образование активных гормонов происходит путем частичного протеолиза.
 синтезируется в передней")
Образование пептидных гормонов, являющихся продуктами общего гена: А - ПОМК (проопиомеланокортин) синтезируется в передней и промежуточной долях гипофиза и в некоторых других тканях (кишечнике, плаценте). Полипептидная цепь состоит из 265 аминокислотных остатков; Б - после отщепления N-концевого сигнального пептида полипептидная цепь расщепляется на два фрагмента: АКТГ (39 а. к. ) и β-липотропин (42 -134 а. к. ); В, Г, Д - при дальнейшем протеолизе происходит образование α- и β-МСГ (меланоцитстимулирующего гормона) и эндорфинов. КППДГ - кортикотропиноподобный гормон промежуточной доли гипофиза. Процессинг ПОМК в передней и промежуточной долях гипофиза протекает по-разному, с образованием разного набора пептидов
: кортиколиберин, тиролиберин, гонадолиберин, фоллилиберин, соматолиберин,")
Гормоны гипоталамуса Открыто 7 либеринов (стимуляторов секреции гормонов гипофиза): кортиколиберин, тиролиберин, гонадолиберин, фоллилиберин, соматолиберин, пролактолиберин, меланолиберин, и 3 статина (ингибиторов): соматостатин, меланостатин, пролактостатин. По химической структуре все гормоны гипоталамуса являются низкомолекулярными пептидами.

Гормоны гипофиза Выделяют гормоны передней и задней доли гипофиза.

Гормоны задней доли гипофиза Вазопрессин и окситоцин (синтезируются в гипоталамусе, в комплексе с белками нейрофизинами транспортируются в гипофиз, где откладываются про запас; после диссоциации из комплекса с белком секретируются в кровь). Представляют собой олигопептиды. Биологический эффект окситоцина: сокращение гладких мышц матки при родах, секреция молока. Биологический эффект вазопрессина: сужение сосудов, регуляция водного обмена (антидиуретическое действие).

Гормоны промежуточной доли гипофиза Меланотропин = меланоцитстимулирующий гормон (олигопептид, стимулирует выработку меланина, у животных отвечает за окраску)

Гормоны передней доли гипофиза 1. АКТГ = адренокортикотропный гормон = кортикотропин (39 АМК, стимулирует синтез и секрецию гормонов коры надпочечников). 2. СТГ = гормон роста = соматотропный гормон = соматотропин (191 АМК, стимулирует процессы роста и развития). 3. Лактотропный гормон = пролактин (199 АМК, стимулирует рост внутренних органов, секрецию желтого тела)

4. Тиреотропный гормон = ТТГ = тиротропин (гликопротеин, состоит из 2 субъединиц, контролирует развитие щитовидной железы, синтез и секрецию в кровь тиреоидных гормонов). 5, 6. Гонадотропины: фолликулостимулирующий гормон = ФСГ и лютеинизирующий гормон = ЛГ (гликопротеины, состоят из 2 субъединиц, регулируют стероидо- и гаметогенез в половых железах). 7. Липотропные гормоны = ЛТГ (пептиды, при ограниченном протеолизе образуют биологически активные пептиды, обладающие опиоидным действием (энкефалины, эндорфины), расщепляющие липиды, оказывающие инсулиноподобное, гипокальциемическое, кортикотропное и др. действия)

Патологии при избытке и недостатке гормонов гипофиза Гормон Избыток Недостаток СТГ Акромегалия Карликовость АКТГ Синдром Иценко. Кушинга ТТГ Гипертиреоз Вторичная гипофункция коры надпочечн. Вторичный гипотиреоз Пролактин Аменорея, галакторея, бесплодие Преждевременное созревание Отсутствие лактации ЛГ То же Липотропины Истощение Ожирение ФСГ Бесплодие
")
Гормоны паращитовидных желез Гормоны паращитовидной железы = паратиреоидные гормоны = паратгормоны (гормоны белковой природы) – группа соединений белковой природы, повышает содержание в крови ионов кальция и фосфат-ионов, т. к. способствует вымыванию солей кальция (в виде цитратов и фосфатов) из костной ткани и соответственно деструкции минеральных и органических компонентов костей и увеличивает реабсорбцию кальция в почках.

Гормоны щитовидной железы В фолликулярной части щитовидной железы образуются 2 гормона – тироксин и трийодтиронин. Они запасаются в виде йодсодержащего гликопротеина– йодтиреоглобулина. В парафолликулярных клетках щитовидной железы (Склетках) синтезируется пептидный гормон кальцитонин.

Кальцитонин Гормон пептидной природы, обладает действием, противоположным эффекту паратгормона – подавляет резорбцию кальция из костной ткани.

Йодсодержащие гормоны щитовидной железы Являются производными тиронина, синтезирующегося из АМК тирозина. Биосинтез этих гормонов регулируется тиротропином. Биологическое действие: действуя, как и стероиды, на генетический аппарат, усиливают синтез ферментов, регулирующих окислительно-восстановительные процессы. Гипофункция вызывает кретинизм или микседему. Гиперфункция – гипертиреоз (зоб диффузный токсический = базедова болезнь). Не путать с эндемическим зобом (при недостатке йода)! Синтез тиреоидных гормонов тормозят антагонисты тироксина – тиомочевина, тиоурацил, метилтиоурацил.

В щитовидной железе синтезируются гормоны - йодированные производные тирозина. Они объединены общим названием йодтиронины. К ним относят 3, 5, 3'-трийодтиронин (трийодтиронин, Т 3) и 3, 5, 3', 5'тетрайодтиронин (Т 4), или тироксин. Йодтиронины участвуют в регуляции многих процессов метаболизма, развития, клеточной дифференцировки, в регуляции экспрессии генов. Заболевания, возникающие в результате нарушений синтеза, секреции и функций йодтиронинов, - наиболее распространённые заболевания эндокринной системы.

Гормоны поджелудочной железы Поджелудочная железа – железа смешанной секреции (внешняя и внутренняя – гормональная - секреция). Внутрисекреторную функцию выполняют клетки островков Лангерганса: α-клетки (А-клетки) секретируют глюкагон; β-клетки (В-клетки) – инсулин; δ-клетки (D-клетки) – соматостатин; F-клетки – малоизученный панкреатический полипептид.
, образованных из одного")
Инсулин Состоит из 2 пептидных цепей (т. е. гормон белковой природы), образованных из одного полипептида путем частичного протеолиза. Повышение глюкозы в крови вызывает повышение секреции инсулина, понижение – замедляет ее.

Этапы синтеза и посттрансляционной модификации инсулина 1 - элонгация сигнального пептида на полирибосомах ЭР с образованием препроинсулина; 2 - отщепление сигнального пептида от препроинсулина; 3 - частичный протеолиз проинсулина с образованием инсулина и С-пептида; 4 - включение инсулина и С-пептида в секреторные гранулы; 5 - секреция инсулина и С-пептида из β-клеток поджелудочной железы в кровь

Инсулин: - снижает уровень сахара в крови; - усиливает синтез из глюкозы гликогена в мышцах; - усиливает анаболические процессы; - повышает скорость утилизации глюкозы в тканях; - оказывает опосредованное влияние на водный и минеральный обмен.

Глюкагон 29 АМК. По биологическому действию – антагонист инсулина. Вызывает: - ускорение распада гликогена в печени; - торможение синтеза гликогена в печени; - повышение концентрации глюкозы в крови, в том числе за счет образования глюкозы из промежуточных продуктов обмена белков и жиров – глюконеогенеза; - торможение распада глюкозы до молочной кислоты (в отличие от адреналина); - активация обмена жиров (в отличие от адреналина). Механизм действия – через мембранные рецепторы клеток и образование ц. АМФ.
 и коркового")
Гормоны надпочечников Надпочечники состоят из мозгового вещества (вырабатывает гормоны – производные АМК) и коркового вещества (вырабатывает стероидные гормоны - кортикостероиды).

Гормоны мозгового вещества надпочечников Адреналин и норадреналин. Предшественник этих гормонов – тирозин → ДОФА → дофамин → норадреналин → адреналин Биологический эффект адреналина: -сосудосуживающее действие; - повышение давления; - ускорение распада гликогена в печени; - торможение окисления глюкозы.

Адреналин и норадреналин, как и дофамин, относятся к катехоламинам. Кроме того, все они оказывают мощное сосудосуживающее действие, вызывая повышение артериального давления, и в этом отношении действие их сходно с действием симпатической нервной системы. Известно мощное регулирующее влияние этих гормонов на обмен углеводов в организме. Адреналин вызывает резкое повышение уровня глюкозы в крови, что обусловлено ускорением распада гликогена в печени под действием фермента фосфорилазы

Гормоны коркового вещества надпочечников Корковое вещество образует кортикостероиды: глюкокортикоиды (кортикостерон, кортизон, гидрокортизон и др. ) и минералкортикоиды (дезоксикортикостерон, альдостерон и др. ). Являются стероидами, а именно производными прегнана. Образуются из холестерина, процесс образования регулируется АКТГ и, соответственно кортиколиберином.

Глюкокортикоиды -в мышечной, лимфатической, соединительной и жировой ткани оказывают катаболическое действие (за счет снижения проницаемости клеточных мембран), в печени – противоположное действие; - стимулируют глюконеогенез; - снижают синтез гликогена в мышцах; - тормозят окисление глюкозы в тканях; - усиление распада жиров, дающих энергию; - усиливают синтез некоторых белков; - вызывают гипергликемию.

Минералкортикоиды - способствуют удержанию ионов натрия и хлора в организме; - способствуют выведению с мочой ионов калия.

Половые гормоны Существует 2 группы женских половых гормонов, различающихся своей химической структурой и биологической функцией: эстрогены (главный представитель – эстрадиол) и прогестины (главный представитель – прогестерон). Основным местом синтеза женских половых гормонов – эстрогенов (от греч. oistros – страстное влечение) – являются яичники и желтое тело; доказано также образование этих гормонов в надпочечниках, семенниках и плаценте.
 осуществляется главным образом в семенниках и частично в яичниках")
Биосинтез андрогенов (мужских половых гормонов) осуществляется главным образом в семенниках и частично в яичниках и надпочечниках.

С 27 – холестерин, С 21 — гормоны коры надпочечников и прогестерон, С 19 — мужские половые гормоны (андрогены и тестостерон), С 18 — женские половые гормоны (эстрогены). С 19 С 18, ароматизация и деметилирование С 21

Гормоны, производные жирорастворимых витаминов
 стимулирует экспрессию генов многих рецепторов к факторам роста.")
Ретиноевая кислота (гормональная форма витамина А) стимулирует экспрессию генов многих рецепторов к факторам роста. Иными словами, повышает чувствительность клеток к ростовым стимулам. Благодаря этому она: ● регулирует нормальный рост и дифференцировку клеток эмбриона и молодого организма, ● регулирует деление и дифференцировку быстро делящихся тканей – хряща, костной ткани, сперматогенного эпителия, плаценты, эпителия кожи, слизистых, иммунной системы.

Синтез витаминов группы D

Витамин представлен двумя формами – эргокальциферол и холекальциферол. Химически эргокальциферол отличается от холекальциферола наличием в молекуле двойной связи между С 22 и С 23 и метильной группой при С 24. После всасывания в кишечнике или после синтеза в коже витамин попадает в печень. Здесь он гидроксилируется по С 25 и кальциферолтранспортным белком переносится к почкам, где еще раз гидроксилируется, уже по С 1. Образуется 1, 25 -дигидроксихолекальциферол или кальцитриол.
 выполняет гормональную функцию, участвуя в регуляции обмена Са 2+")
Кальцитриол 1, 25 -дигидроксихолекальциферол (кальцитриол) выполняет гормональную функцию, участвуя в регуляции обмена Са 2+ и фосфатов, стимулируя всасывание Са 2+ в кишечнике и кальцификацию костной ткани, реабсорбцию ионов кальция и фосфатов в почках. При низкой концентрации Са 2+ или высокой концентрации D 3 он стимулирует мобилизацию Са 2+ из костей.

Эйкозаноиды

Предшественником всех эйкозаноидов являются полиненасыщенные жирные кислоты, в частности арахидоновая кислота (и ряд ее производных, дигомо-γлиноленовая и пентаноевая кислоты, в свою очередь образующиеся в организме из линолевой и линоленовой кислот). Арахидоновая кислота после освобождения из фосфоглицеринов (фосфолипидов) биомембран под действием специфических фосфолипаз А (или С) в зависимости от ферментативного пути превращения дает начало простагландинам, простациклинам, тромбоксанам и лейкотриенам.

Синтез эйкозаноидов из арахидоновои кислоты. Глюкокортикоиды ингибируют синтез всех типов эйкозаноидов, так как ингибируют фосфолипазу А 2, и таким образом уменьшают количество субстрата для их синтеза. Аспирин и другие противовоспалительные препараты нестероидного действия ингибируют только циклооксигеназный путь.

Эйкозаноиды – это гормоны местного действия. Они синтезируясь в клетках регулируют многочисленные клеточные и тканевые функции, включая агрегацию тромбоцитов, воспалительные реакции и функционирование лейкоцитов, сужение и расширение сосудов, кровяное давление, бронхиальные сокращения и сокращение матки. В послеоперацинный период хорошо бы знать индекс: простациклины (ингибиторы агрегации тромбоцитов) тромбоксаны (активаторы агрегации тромбоцитов).

КАТАБОЛИЗМ ГОРМОНОВ. Белково-пептидные гормоны подвергаются протеолизу, распадаются до отдельных аминокислот. Эти аминокислоты вступают дальше в реакции дезаминирования, декарбоксилирования, трансаминирования и распадаются до конечных продуктов: NH 3, CO 2 и Н 2 О. Гормоны - производные аминокислот подвергаются окислительному дезаминированию и дальнейшему окислению до СО 2 и Н 2 О. Стероидные гормоны распадаются иначе. В организме нет ферментных систем, которые обеспечивали бы их распад. Что же происходит при их катаболизме?

В основном происходит модификация боковых радикалов. Вводятся дополнительные гидроксильные группы. Гормоны становятся более гидрофильными. Образуются молекулы, представляющие собой структуру стерана, у которого в 17 -м положении находится кетогруппа. В таком виде продукты катаболизма стероидных половых гормонов выводятся с мочой и называются 17 -КЕТОСТЕРОИДЫ. Определение их количества в моче и крови показывает содержание в организме половых гормонов. У мужчин 2/3 кетостероидов образуется за счет кортикостероидов и 1/3 - за счет тестостерона (всего 12 -17 мг в сутки). У женщин 17 -кетостероиды образуются преимущественно за счет кортикостероидов (7 -12 мг в сутки).

Биологическое действие гормонов проявляется через их взаимодействие с клетками, имеющими рецепторы к данному гормону (клетками-мишенями). Для проявления биологической активности связывание гормона с рецептором должно приводить к образованию химического сигнала внутри клетки, который вызывает специфический биологический ответ, например, изменение скорости синтеза ферментов и других белков или изменение их активности. Мишенью для гормона могут служить клетки одной или нескольких тканей. Воздействуя на клетку-мишень, гормон вызывает специфическую ответную реакцию, проявление которой зависит от того, какие метаболические пути активируются или тормозятся в этой клетке. • Рецепторы гормонов могут быть расположены или в плазматической мембране или внутри клетки (в цитозоле или ядре). • По механизму действия гормоны можно разделить на две группы: - к первой группе относятся гормоны, взаимодействующие с мембранными рецепторами (например, пептидные гормоны, адреналин, а также гормоны местного действия - цитокины, эйкозаноиды); - вторая группа включает гормоны, взаимодействующие с внутриклеточными рецепторами - стероидные гормоны и йодсодержащие гормоны.
 Внутриклеточные рецепторы (гормоны стероидной природы, тиреоидные гормоны)")
Гормональный сигнал Мембранные рецепторы (пептидные гормоны, адреналин) Внутриклеточные рецепторы (гормоны стероидной природы, тиреоидные гормоны) Изменение активности имеющихся ферментов через систему вторичных мессенджеров Синтез новых ферментов
 с рецептором приводит к изменению конформации рецептора. Это изменение улавливается")
Связывание гормона (первичного посредника) с рецептором приводит к изменению конформации рецептора. Это изменение улавливается другими макромолекулами, т. е. связывание гормона с рецептором приводит к сопряжению одних молекул с другими (трансдукция сигнала). Таким образом, генерируется сигнал, который регулирует клеточный ответ путём изменения активности или количества ферментов и других белков. В зависимости от способа передачи гормонального сигнала в клетках меняется скорость реакций метаболизма: в результате изменения активности ферментов; в результате изменения количества ферментов.

Передача сигнала с помощью мембранных рецепторов.

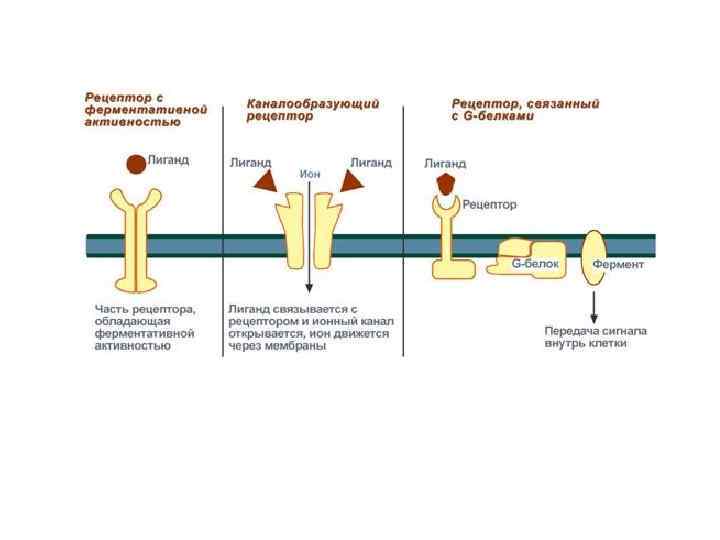
Мембранные рецепторы подразделяются на: • рецепторы, содержащие субъединицу, связывающую первичный мессенджер, и ионный канал; • рецепторы, способные проявлять каталитическую активность; • рецепторы, с помощью G-белков активирующие образование вторичных (внутриклеточных) мессенджеров, передающих сигнал специфическим белкам и ферментам цитозоля. Рецепторы: 1 - содержащие субъединицу, связывающую сигнальную молекулу и ионный канал, например рецептор ацетилхолина на постсинаптической мембране; 2 проявляющие каталитическую активность после присоединения сигнальной молекулы, например рецептор инсулина; 3, 4 - передающие сигнал на фермент аденилатциклазу (АЦ) или фосфолипазу С (ФЛС) при участии мембранных G-белков, например разные типы рецепторов адреналина, ацетилхолина и других сигнальных молекул

ПОДРОБНЕЕ: 1. Рецепторы, обладающие каталитической активностью – при взаимодействии лиганда с рецептором активируется внутриклеточная часть (домен) рецептора, имеющий тирозинкиназную или тирозинфосфатазную или гуанилатциклазную активность. По этому механизму действуют СТГ, инсулин, пролактин, интерлейкины, ростовые факторы, интерфероны α, β, γ. 2. Каналообразующие рецепторы – присоединение лиганда к рецептору вызывает открытие ионного канала на мембране. Таким образом действуют нейромедиаторы (ацетилхолин, глицин, ГАМК, серотонин, гистамин, глутамат); 3. Рецепторы, связанные с G-белками – передача сигнала от гормона происходит при посредстве G-белка. G-белок влияет на ферменты, образующие вторичные мессенджеры (посредники ). Последние передают сигнал на внутриклеточные белки. Большинство гормонов действуют по данному механизму. Роль вторичных мессенджеров выполняют молекулы и ионы: • ц. АМФ (циклический аденозин-3', 5'-монофосфат); • ц. ГМФ (циклический гуанозин-3', 5'-монофосфат); • ИФ 3 (инозитол-1, 4, 5 -трифосфат); • ДАГ (диацилглицерол); • Са 2+; • NO.

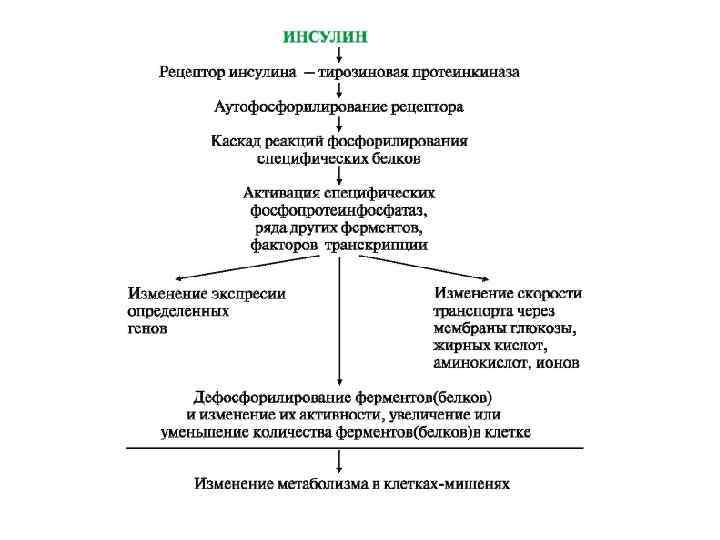
Каталитические рецепторы являются ферментами. Активаторами этих ферментов могут быть гормоны, ростовые факторы, цитокины. При участии специальных механизмов сигнал, полученный каталитическим рецептором, может быть передан в ядро, где он стимулирует или подавляет экспрессию определенных генов. Рецептор инсулина - тирозиновая протеинкиназа (Тир-ПК). Рецепторные Тир-ПК участвуют в трансмембранной передаче сигналов. В активной форме это ферменты, фосфорилирующие специфические белки по ОН-группам тирозина. Рецептор инсулина построен из 2 α- и 2 β-субъединиц. αСубъединицы расположены на наружной поверхности клеточной мембраны, β-субъединицы пронизывают мембранный бислой. Центр связывания инсулина образован N-концевыми доменами α-субъединиц. Каталитический центр рецептора находится на внутриклеточных доменах β-субъединиц. Цитозольная часть рецептора имеет несколько остатков тирозина, которые могут фосфорилироваться и дефосфорилироваться.

Присоединение инсулина в центр связывания, образованный αсубъединицами, вызывает кооперативные конформационные изменения рецептора. β-Субъединицы проявляют тирозинкиназную активность и катализируют трансаутофосфорилирование (первая β-субъединица фосфорилирует вторую β-субъединицу, и наоборот) по нескольким остаткам тирозина. Фосфорилирование приводит к изменению заряда, конформации и субстратной специфичности фермента (Тир-ПК). Тирозиновая-ПК фосфорилирует определенные клеточные белки, которые получили название субстратов рецептора инсулина. В свою очередь эти белки участвуют в активации каскада реакций фосфорилирования: • фосфопротеинфосфатазы (ФПФ), которая дефосфорилирует специфические фосфопротеины; • фосфодиэстеразы, которая превращает ц. АМФ в АМФ и ц. ГМФ в ГМФ; • ГЛЮТ 4 - переносчиков глюкозы в инсулинзависимых тканях, поэтому повышается поступление глюкозы в клетки мышц и жировой ткани; • тирозиновой протеинфосфатазы, которая дефосфорилирует βсубъединицы рецептора инсулина; • регуляторных белков ядра, факторов транскрипции, повышающих или снижающих экспрессию генов определенных ферментов.

Активация рецептора инсулина.
.")
Механизм действия инсулина Эффекты инсулина реализуются через специфические мембранные рецепторы (практически у всех клеток). Рецепторы инсулина состоят из α- и β-цепей. Инсулин связывается с α-цепями, в результате чего активируются протеинкиназные свойства β-цепей – запускается каскад фосфорилирования-дефосфорилирования протеинкиназ. Рецепторы инсулина сами обладают протеинкиназной активностью (в отличие от адреналина, глюкагона)

К каталитическим рецепторам также относятся рецепторы с гуанилатциклазной активностью. Гуанилатциклаза катализирует образование из ГТФ ц. ГМФ, который является одним из важных мессенджеров внутриклеточной передачи сигнала. Гуанилатциклазный механизм пока изучен недостаточно подробно, но известно, что по нему работает атриопептин (предсердный натрийуретический пептид), регулирующий гомеостаз жидкости в организме, а также оксид азота (NO) и другие, пока малоизученные, соединения. Гуанилатциклазный механизм передачи сигнала в общих чертах схож с аденилатциклазным: после появления какой-либо сигнальной молекулы сигнал воспринимается ферментом гуанилатциклазой и далее передается при помощи вторичного мессенджера ц. ГМФ. Он воздействует на протеинкиназу G, которая фосфорилирует определенные белки, чем изменяет активность их в клетке. Гуанилатциклаза имеет четыре разновидности, три из которых связаны с мембраной, четвертый – цитозольный: - мембраносвязанные формы гуанилатциклазы (трансмембранные гликопротеины) работают как рецепторы, обладающие каталитической активностью. - цитозольный фермент является димером и содержит в своем составе гем, он непосредственно взаимодействует в цитозоле с сигнальной молекулой (например, оксидом азота).

Регуляция активности мембранной гуанилатциклазы.

Рецепторы, сопряженные с G-белками Взаимодействие гормонов с рецепторами, сопряженными с G -белками, приводит к активации инозитолфосфатной системы трансдукции сигнала или изменению активности аденилатциклазной регуляторной системы.

G-белки Универсальные посредники при передаче сигналов от рецепторов к ферментам клеточных мембран, катализирующих образование вторичных посредников гормонального сигнала. G-белки - олигомеры, состоят из 3 субъединиц α, β, γ. Есть активаторные Gsбелки, они активируют мембранные ферменты, есть ингибиторные Gi-белки, они ингибируют мембранные ферменты. G-белок Г R R Ац β γ γ α α 1). 2). ГТФ ГДФ Г G-белок R R Ац β γ α α 3). ГТФ АТФ ц. АМФ ГДФ Фн 1). Гормон (Г), взаимодействуя с рецептором (R), изменяет его конформацию. 2). Рецептор, взаимодействуя с Gбелком, уменьшает у αсубъединицы (α) сродство к ГДФ и увеличивает сродство к ГТФ. 3). Присоединение ГТФ к αсубъединице вызывает в G-белке изменение конформации и диссоциацию его на субъединицы: α-субъединицу (α-ГТФ) и димер βγ. α-ГТФ имеет высокое сродство к аденилатциклазе (Ац), его присоединение приводит к активации последней. 4). α-субъединица катализирует распад ГТФ до ГДФ + Фн. α-ГДФ имеет низкое сродство к Ац и высокое к димеру βγ. Отделение αГДФ от Ац инактивирует последнюю.
 Гликопротеин, имеет 8 изоформ, ключевой фермент аденилатциклазной системы, катализирует образование вторичного посредника")
Аденилатциклаза (Ац) Гликопротеин, имеет 8 изоформ, ключевой фермент аденилатциклазной системы, катализирует образование вторичного посредника ц. АМФ из АТФ. Есть во всех клетках, располагается на внутренней стороне клеточной мембраны. Активность Ац контролируется: 1) внеклеточными регуляторами - гормонами, эйкозаноидами, биогенными аминами через G-белки; 2) внутриклеточным регулятором Са 2+. Протеинкиназа А (ПК А) ПК А есть во всех клетках. Участвует в аденилатциклазной системе, стимулируется ц. АМФ. Состоит из 4 субъединиц: 2 регуляторных (R) и 2 каталитических (С). Регуляторные субъединицы имеют по 2 участка связывания ц. АМФ. Тетрамер не обладает каталитической активностью. Присоединение 4 ц. АМФ к 2 субъединицам R приводит к изменению их конформации и диссоциации тетрамера. При этом высвобождаются 2 активные каталитические субъединицы С, которые катализируют реакцию фосфорилирования белков-ферментов. Фосфодиэстеразы (ФДЭ) ФДЭ превращает ц. АМФ и ц. ГМФ в АМФ и ГМФ, инактивируя аденилатциклазную и гуанилатциклазную систему. ФДЭ активируется Са 2+, 4 Са 2+-кальмодулином, ц. ГМФ.

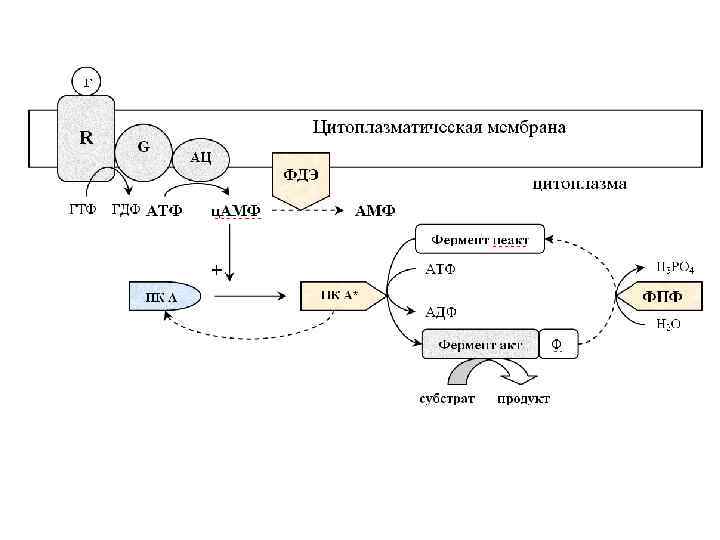
Трансмембранная передача информации с участием аденилатциклазной системы
. 1 Гормон присоединяется к Rs-рецептору с образованием гормонрецепторного комплекса, который через несколько Gs-белков")
1). 1 Гормон присоединяется к Rs-рецептору с образованием гормонрецепторного комплекса, который через несколько Gs-белков активирует несколько аденилатциклаз (комплекс гормон-Ri-рецептор через Gi-белки ингибирует аденилатциклазы); 2). Одна аденилатциклаза превращает тысячи АТФ в тысячи ц. АМФ; 3). 4 ц. АМФ обратимо присоединяясь к 2 рецепторным субъединицам R ПК А, вызывают диссоциацию протомеров ПК А с освобождением 2 активных каталитических субъединиц С и двух 2 ц. АМФ-R; 4). Одна субъединица С ПК А фосфорилирует сотни ферментов, что способствует их активации или ингибированию. Таким образом, сигнал одной молекулы гормона в аденилатциклазной системе суммарно усиливается в 106107 раз; 5). Миллионы активных ферментов превращают молекулы субстрата в продукты. Инактивация аденилатциклазной системы осуществляется через ФДЭ и фосфотазу. ФДЭ разрушает ц. АМФ до АМФ, это приводит к самосборке субъединиц ПК А и ее инактивации. Фосфотаза инактивирует (активирует) фермент в реакции гидролитического дефосфорилирования. Посредством стимуляции аденилатциклазной системы оказывают гормональное действие кортиколиберин, кальцитонин, соматолиберин, ВИП (вазоактивный интестинальный полипептид), глюкагон, вазопрессин (через V 2 рецепторы), ЛГ, ФСГ, ТТГ, хорионический гонадотропин, АКТГ, паратгормон, простагландины типа Е, D и I, адренергические катехоламины (через β 1 и β 2 рецепторы).
 Гидролизует фосфоэфирную связь")
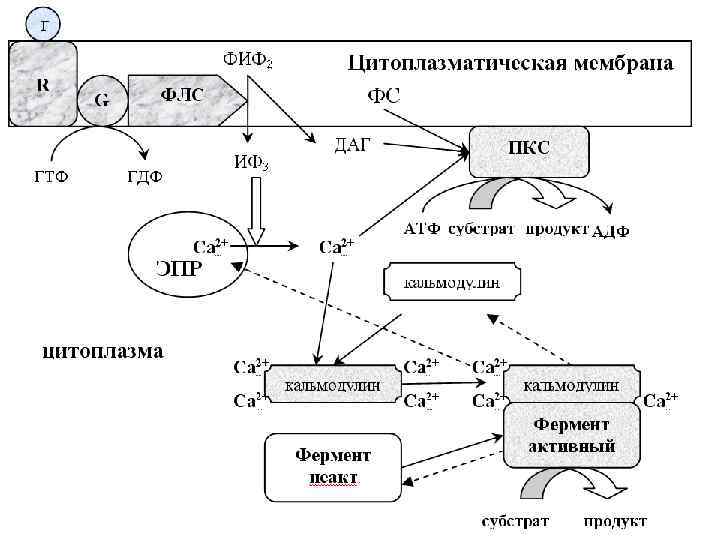
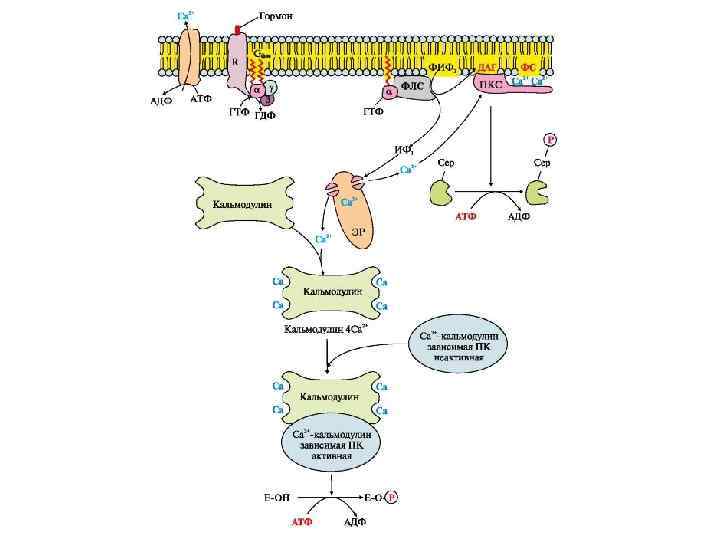
Трансмембранная передача информации с участием инозитолтрифосфатной системы Фосфолипаза С (ФЛ С) Гидролизует фосфоэфирную связь в фосфатидилинозитолах с образованием ДАГ и ИФ 3, имеет 10 изоформ. ФЛ С регулируется через G-белки и активируется Са 2+. Инозитолтрифосфат открывает кальциевые каналы в эндоплазматическом ретикулуме, что вызывает увеличение концентрации ионов Cа 2+. Диацилглицерол совместно с ионами Са 2+ активирует протеинкиназу С. Кроме этого, диацилглицерол имеет и другую сигнальную функцию: он может распадаться на 1 -моноацилглицерол и полиеновую жирную кислоту (обычно арахидоновую), из которой образуются эйкозаноиды. Протеинкиназа С фосфорилирует ряд ферментов и в целом участвует в процессах клеточной пролиферации.
. Гормон (Г) присоединяется к R-рецептору с")
Последовательность событий, приводящих к каталитической активации ферментов 1). Гормон (Г) присоединяется к R-рецептору с образованием гормонрецепторного комплекса, который через G-белок активирует фосфолипазу С; 2). Фосфолипаза С расщепляет фосфатидилинозитол-4, 5 -дифосфат (ФИФ 2) клеточной мембраны на инозитолтрифосфат (ИФ 3) и диацилглицерин (ДАГ); 3). ИФ 3 присоединяясь к рецептору на мембране ЭПР, открывает кальциевые каналы, выпуская Са 2+ из ЭПР в цитоплазму; 4). Са 2+, ДАГ активируют ПК С, которая превращает субстрат в продукт. 5). 4 Са 2+ присоединяются к кальмодулину, способствуя присоединению кальмодулина к ферменту и образованию активного комплекса 4 Са 2+кальмодулин-фермент, который превращает субстрат в продукт. Инактивация инозитолтрифосфатной системы осуществляется кальциевыми насосами, которые откачивают Са 2+ из цитоплазмы. Гидролиз ФИФ 2 продолжается некоторое время, пока α-субъединица G-белка, которая является ГТФ-азой, отщепляет фосфат от ГТФ. Как только ГТФ превратился в ГДФ, то α-субъединица инактивируется, теряет свое влияние на фосфолипазу C, обратно соединяется с β- и γ-субъединицами. Все возвращается в исходное положение. Посредством стимуляции инозитолтрифосфатной системы оказывают гормональное действие гонадолиберин, тиролиберин, дофамин, тромбоксаны А 2, эндоперекиси, лейкотриены, агниотензин II, эндотелин, паратгормон, нейропептид Y, адренергические катехоламины (через α 1 рецепторы), ацетилхолин, брадикинин, вазопрессин (через V 1 рецепторы).

Передача сигнала с помощью внутриклеточных рецепторов.
 могут диффундировать через мембраны, поэтому")
Гидрофобные по химической природе гормоны (стероидные гормоны и тироксин) могут диффундировать через мембраны, поэтому их рецепторы находятся в цитозоле или ядре клетки. Цитозольные рецепторы связаны с белком-шапероном, который предотвращает преждевременную активацию рецептора. Ядерные и цитозольные рецепторы стероидных и тиреоидных гормонов содержат ДНК-связывающий домен, обеспечивающий в ядре взаимодействие комплекса гормон-рецептор с регуляторными участками ДНК и изменение скорости транскрипции.

Последовательность событий, приводящих к изменению скорости транскрипции - Гормон проходит через двойной липидный слой клеточной мембраны. - В цитозоле или ядре гормон взаимодействует с рецептором. - Комплекс гормон-рецептор проходит в ядро и присоединяется к регуляторной нуклеотидной последовательности ДНК - энхансеру или сайленсеру. Доступность промотора для РНК-полимеразы увеличивается при взаимодействии с энхансером или уменьшается при взаимодействии с сайленсером. Соответственно увеличивается или уменьшается скорость транскрипции определенных структурных генов. - Зрелые м. РНК выходят из ядра. - Увеличивается или уменьшается скорость трансляции определенных белков. - Изменяется количество белков, которые влияют на метаболизм и функциональное состояние клетки.

Иногда, гормон самостоятельно проникает в ядро, где соединяется с ядерным рецептором, образуя гормонрецепторный комплекс. Этот комплекс также присоединяется к ДНК, запускает или блокирует процесс транскрипции м. РНК и трансляции новых ферментов. Эффекты гормонов, которые взаимодействуют с внутриклеточными рецепторами, нельзя наблюдать быстро, так как на реализацию процессов транскрипции и трансляции, которые они стимулируют, требуются часы.

Торможение гормонального сигнала: носитель сигнала должен быстро разрушаться • Разрушение гормонов под действием ферментов • Самовыключение G-белков. (Активные G-белки связаны с ГТФ. G-белки обладают ГТФ-азной активностью и со временем превращают ГТФ в ГДФ, что способствует переходу их активного центра в неактивное состояние). • ц-3’, 5’-АМФ и ц-3’, 5’-ГМФ под действием фосфодиэстеразы превращаются , соответственно, в АМФ и ГМФ. • Ионы Са 2+ выводятся из цитоплазмы. • Диацилглицерины разрушаются под действием липаз. • Фосфорилированные ферменты возвращаются в исходное состояние, подвергаясь дефосфорилированию под действием протеинфосфатаз.
Гормоны.ppt