Мейоз 2014 - 10.ppt
- Количество слайдов: 33
 Мейоз Домашнее задание: § 40, записи в тетради, материалы сайта.
Мейоз Домашнее задание: § 40, записи в тетради, материалы сайта.
 - форма ядерного деления довольно сложного типа, сопровождающаяся") Мейоз (от греч. meiosis - уменьшение) - форма ядерного деления довольно сложного типа, сопровождающаяся уменьшением числа хромосом от диплоидного (2 n) до гаплоидного (n). Его отличительной особенностью является образование из одной диплоидной клетки четырех дочерних клеток с гаплоидным набором хромосом. Второй отличительной особенностью мейоза является то, что он протекает в два деления: первое деление называется редукционным, второе - эквационным. Интерфаза между делениями очень короткая, во время которой не происходит удвоение ДНК. В механизмах мейоза есть также отличительные особенности, которые затрагивают в большей степени профазу первого деления. Необходимо отметить, что мейоз представляет собой универсальное явление, характерное для всех эукариотических организмов. При мейозе происходит не только редукция числа хромосом до гаплоидного их числа, но и осуществляется чрезвычайно важный генетический процесс обмен участками между гомологичными хромосомами, процесс, получивший название кроссинговера.
Мейоз (от греч. meiosis - уменьшение) - форма ядерного деления довольно сложного типа, сопровождающаяся уменьшением числа хромосом от диплоидного (2 n) до гаплоидного (n). Его отличительной особенностью является образование из одной диплоидной клетки четырех дочерних клеток с гаплоидным набором хромосом. Второй отличительной особенностью мейоза является то, что он протекает в два деления: первое деление называется редукционным, второе - эквационным. Интерфаза между делениями очень короткая, во время которой не происходит удвоение ДНК. В механизмах мейоза есть также отличительные особенности, которые затрагивают в большей степени профазу первого деления. Необходимо отметить, что мейоз представляет собой универсальное явление, характерное для всех эукариотических организмов. При мейозе происходит не только редукция числа хромосом до гаплоидного их числа, но и осуществляется чрезвычайно важный генетический процесс обмен участками между гомологичными хромосомами, процесс, получивший название кроссинговера.

 Существует несколько разновидностей мейоза. При зиготном типе мейоза, характерном для аскомицетов, базимицетов, некоторых водорослей, споровиков и др. , для которых в жизненном цикле преобладает гаплоидная фаза, две клетки - гаметы - сливаются, образуя зиготу с двойным (диплоидным) набором хромосом. В таком виде диплоидная зигота (покоящаяся спора) приступает к мейозу. дважды делится, в результате образуются четыре гаплоидные клетки, которые продолжают размножаться. Споровый тип мейоза встречается у высших растений, клетки которых имеют диплоидный набор хромосом. В данном случае в органах размножения растений образовавшиеся после мейоза гаплоидные клетки еще несколько раз делятся. Другой тип мейоза, гаметный, происходит во время созревания гамет предшественников зрелых половых клеток. Он встречается у многоклеточных животных, среди некоторых низших растений. В случае гаметного мейоза при развитии организма происходит выделение клонов герминативных клеток, которые впоследствии будут дифференцироваться в половые клетки. И только клетки этих клонов будут при созревании подвергаться мейозу и превращаться в половые клетки. Следовательно, все клетки развивающихся многоклеточных животных организмов можно разделить на две группы: соматические, из которых будут образовываться клетки всех тканей и органов, и герминативные, которые дадут начало половым клеткам.
Существует несколько разновидностей мейоза. При зиготном типе мейоза, характерном для аскомицетов, базимицетов, некоторых водорослей, споровиков и др. , для которых в жизненном цикле преобладает гаплоидная фаза, две клетки - гаметы - сливаются, образуя зиготу с двойным (диплоидным) набором хромосом. В таком виде диплоидная зигота (покоящаяся спора) приступает к мейозу. дважды делится, в результате образуются четыре гаплоидные клетки, которые продолжают размножаться. Споровый тип мейоза встречается у высших растений, клетки которых имеют диплоидный набор хромосом. В данном случае в органах размножения растений образовавшиеся после мейоза гаплоидные клетки еще несколько раз делятся. Другой тип мейоза, гаметный, происходит во время созревания гамет предшественников зрелых половых клеток. Он встречается у многоклеточных животных, среди некоторых низших растений. В случае гаметного мейоза при развитии организма происходит выделение клонов герминативных клеток, которые впоследствии будут дифференцироваться в половые клетки. И только клетки этих клонов будут при созревании подвергаться мейозу и превращаться в половые клетки. Следовательно, все клетки развивающихся многоклеточных животных организмов можно разделить на две группы: соматические, из которых будут образовываться клетки всех тканей и органов, и герминативные, которые дадут начало половым клеткам.
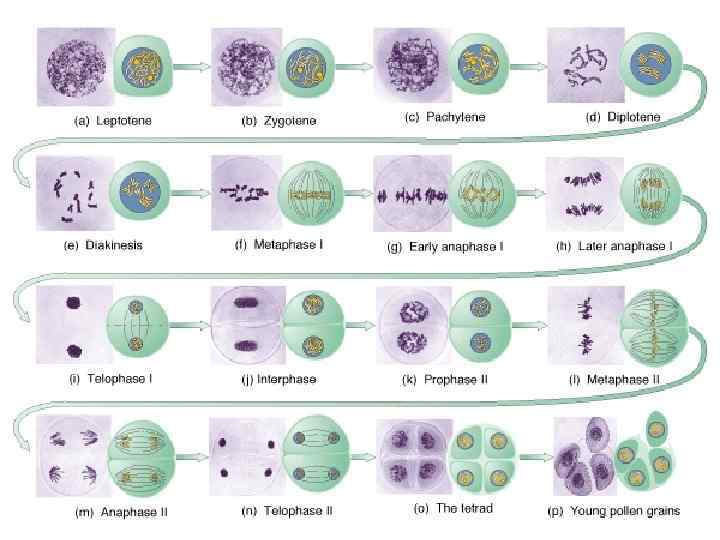
 Стадии мейоза
Стадии мейоза
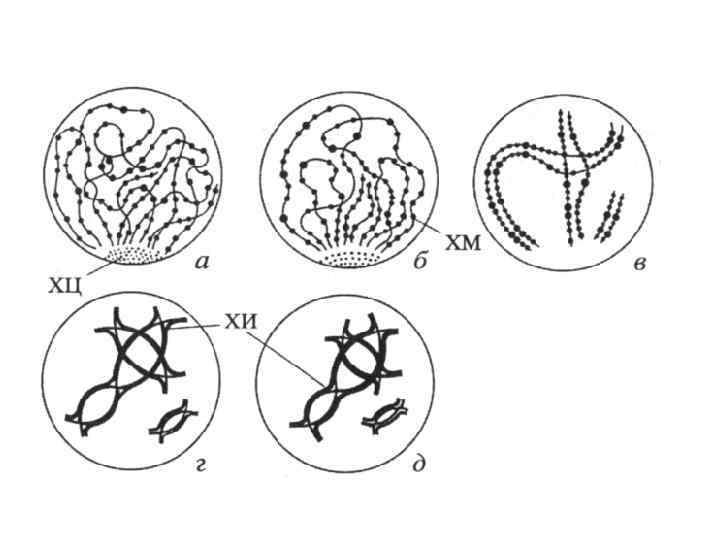
 а - лептотена; б - зиготена; в - пахитена с тремя бивалентами; г - диплотена; д -диакинез; е - метафаза I; ж - анафаза I; з - метафаза II; и - анафаза II. ХЦ -хромоцентр - место локализации теломерных участков хромосом в «букете» ; ХМ -хромомеры; ХИ – хиазма.
а - лептотена; б - зиготена; в - пахитена с тремя бивалентами; г - диплотена; д -диакинез; е - метафаза I; ж - анафаза I; з - метафаза II; и - анафаза II. ХЦ -хромоцентр - место локализации теломерных участков хромосом в «букете» ; ХМ -хромомеры; ХИ – хиазма.
 Профаза 1 - самая продолжительная и сложная стадия мейоза, ее делят на 5 последовательных периодов: лептотена, зиготена, пахитена, диплотена и диакинез. Мейотическая профаза сперматоцита I порядка у самцов мыши длится 12 сут. у человека - 24 дня (плюс еще около двух месяцев до полного созревания сперматозоида). Среди женских половых клеток профаза I порядка тритона обыкновенного длится около 1 года, у мыши от 4 месяцев до 3 лет, у человека профаза I ооцитов начинается на третьем месяце внутриутробного развития и может продолжаться до 50 -летнего возраста женщины.
Профаза 1 - самая продолжительная и сложная стадия мейоза, ее делят на 5 последовательных периодов: лептотена, зиготена, пахитена, диплотена и диакинез. Мейотическая профаза сперматоцита I порядка у самцов мыши длится 12 сут. у человека - 24 дня (плюс еще около двух месяцев до полного созревания сперматозоида). Среди женских половых клеток профаза I порядка тритона обыкновенного длится около 1 года, у мыши от 4 месяцев до 3 лет, у человека профаза I ооцитов начинается на третьем месяце внутриутробного развития и может продолжаться до 50 -летнего возраста женщины.
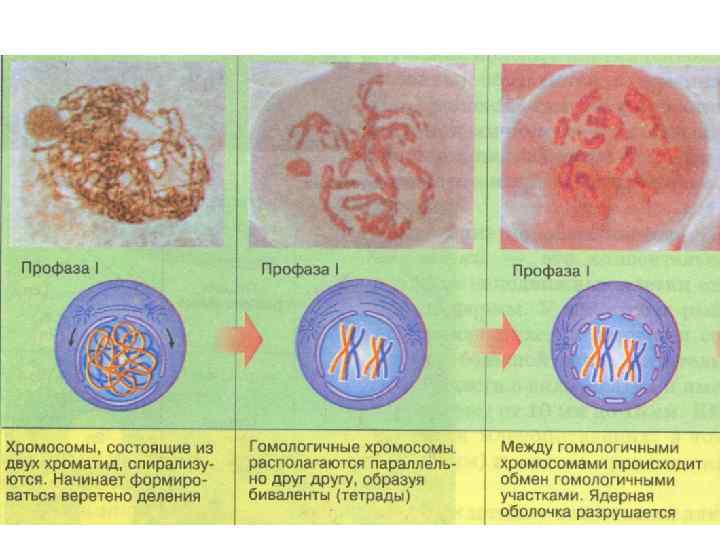
 Лептотена - стадия длинных, тонких, слабо спирализованных хромосом, на которых видны утолщения – хромомеры ( «нитки бус» ). В это время заканчивается репликация ДНК, которая была деспирализована перед мейозом. Хромосомы представляют длинные тонкие нити, каждая хромосома состоит из двух хроматид. Хроматиды имеют хромомерное строение.
Лептотена - стадия длинных, тонких, слабо спирализованных хромосом, на которых видны утолщения – хромомеры ( «нитки бус» ). В это время заканчивается репликация ДНК, которая была деспирализована перед мейозом. Хромосомы представляют длинные тонкие нити, каждая хромосома состоит из двух хроматид. Хроматиды имеют хромомерное строение.
 В лептотене начинается процесс конъюгации гомологичных хромосом, их сближение, которое начинается в теломерных участках, связанных с ядерной оболочкой. В этих местах образуется сложная специальная структура - тяж белковой природы, синаптонемный комплекс, который позже, в зиготене свяжет гомологичные удвоенные хроматиды. Строение бивалента. a 1—а 2 и А 1 -А 2 — сестринские хроматиды. I — плоскости расхождения хроматид в 1 делении; 2— то же во II делении Синаптонемный комплекс (СК). а - вид в электронном микроскопе; б - схема; в - модель. 1 - хроматин сестринских хромосом; 2 - осевой элемент; 3 - боковые элементы
В лептотене начинается процесс конъюгации гомологичных хромосом, их сближение, которое начинается в теломерных участках, связанных с ядерной оболочкой. В этих местах образуется сложная специальная структура - тяж белковой природы, синаптонемный комплекс, который позже, в зиготене свяжет гомологичные удвоенные хроматиды. Строение бивалента. a 1—а 2 и А 1 -А 2 — сестринские хроматиды. I — плоскости расхождения хроматид в 1 делении; 2— то же во II делении Синаптонемный комплекс (СК). а - вид в электронном микроскопе; б - схема; в - модель. 1 - хроматин сестринских хромосом; 2 - осевой элемент; 3 - боковые элементы
 Зиготена - стадия прохождения конъюгации гомологичных хромосом. При этом гомологичные хромосомы образуют новый хромосомный ансамбль, никогда до этого не встречающийся при клеточном делении, - бивалент.
Зиготена - стадия прохождения конъюгации гомологичных хромосом. При этом гомологичные хромосомы образуют новый хромосомный ансамбль, никогда до этого не встречающийся при клеточном делении, - бивалент.
 Хромомеры гомологичных хромосом лежат рядом. Процесс конъюгации может начинаться в нескольких всего в области центромера), хромосомы в конце соединяются по всей длине. Пару конъюгировавших хромосом называют бивалентом. Биваленты укорачиваются и утолщаются. точках (чаще конъюгации
Хромомеры гомологичных хромосом лежат рядом. Процесс конъюгации может начинаться в нескольких всего в области центромера), хромосомы в конце соединяются по всей длине. Пару конъюгировавших хромосом называют бивалентом. Биваленты укорачиваются и утолщаются. точках (чаще конъюгации
 Биваленты - это парные соединения удвоенных гомологичных хромосом, т. е. каждый бивалент состоит из четырех хроматид.
Биваленты - это парные соединения удвоенных гомологичных хромосом, т. е. каждый бивалент состоит из четырех хроматид.
 Число бивалентов на ядро будет равно гаплоидному числу хромосом. Такой порядок объединения виден и на следующей, пахитенной стадии, а на стадии зиготены он только начинается, и, видимо, именно эта стадия как-то определяет течение данного процесса, во многом еще непонятного. В профазе мейоза, а именно на зиготенной стадии, синтезируется небольшое (0, 3% от всей ДНК клетки) количество специфической ДНК, получившей название ZДНК. У лилейных ZДНК обогащена Г-Ц-парами, состоит из уникальных последовательностей нуклеотидов. Эта ДНК распределена небольшими участками по всей длине хромосом лилии. При обычном митотическом цикле она синтезируется одновременно с основной массой ДНК, но при мейозе - только в зиготенной стадии. Было обнаружено, что если на стадии зиготены подавить с помощью ингибиторов этот небольшой дополнительный синтез ДНК, то конъюгация хромосом прекратится. На основании полученных данных было сделано предположение, что специфические по своему строению участки ZДНК на гомологичных хромосомах еще на С 2 -стадии интерфазы ооцита «узнают» друга и на некоторое время образуют стабильные связи, необходимые для закрепления хромосом одна вдоль другой.
Число бивалентов на ядро будет равно гаплоидному числу хромосом. Такой порядок объединения виден и на следующей, пахитенной стадии, а на стадии зиготены он только начинается, и, видимо, именно эта стадия как-то определяет течение данного процесса, во многом еще непонятного. В профазе мейоза, а именно на зиготенной стадии, синтезируется небольшое (0, 3% от всей ДНК клетки) количество специфической ДНК, получившей название ZДНК. У лилейных ZДНК обогащена Г-Ц-парами, состоит из уникальных последовательностей нуклеотидов. Эта ДНК распределена небольшими участками по всей длине хромосом лилии. При обычном митотическом цикле она синтезируется одновременно с основной массой ДНК, но при мейозе - только в зиготенной стадии. Было обнаружено, что если на стадии зиготены подавить с помощью ингибиторов этот небольшой дополнительный синтез ДНК, то конъюгация хромосом прекратится. На основании полученных данных было сделано предположение, что специфические по своему строению участки ZДНК на гомологичных хромосомах еще на С 2 -стадии интерфазы ооцита «узнают» друга и на некоторое время образуют стабильные связи, необходимые для закрепления хромосом одна вдоль другой.
 Пахитена - стадия толстых нитей. Гомологичные хромосомы, составляющие биваленты частично разделяются, как будто отталкиваются друг от друга. Гомологичные хромосомы все еще соединены между собой в нескольких точках, которые называются хиазмами.
Пахитена - стадия толстых нитей. Гомологичные хромосомы, составляющие биваленты частично разделяются, как будто отталкиваются друг от друга. Гомологичные хромосомы все еще соединены между собой в нескольких точках, которые называются хиазмами.
 Хиазма Схема участка пахитенной хромосомы и синаптонемного комплекса: I общий вид: А, А' - сестринские хромосомы одного гомолога; а, а' сестринские хромосомы второго гомолога: 1 - осевой элемент, 2 боковой элемент, 3 рекомбинационный узелок; II— вид в поперечном разрезе.
Хиазма Схема участка пахитенной хромосомы и синаптонемного комплекса: I общий вид: А, А' - сестринские хромосомы одного гомолога; а, а' сестринские хромосомы второго гомолога: 1 - осевой элемент, 2 боковой элемент, 3 рекомбинационный узелок; II— вид в поперечном разрезе.
. Гомологичные хромосомы") Именно в этих участках может происходить обмен генами кроссинговер (crossing-over - перекрест). Гомологичные хромосомы после кроссинговера не расходятся до анафазы – биваленты сохраняются.
Именно в этих участках может происходить обмен генами кроссинговер (crossing-over - перекрест). Гомологичные хромосомы после кроссинговера не расходятся до анафазы – биваленты сохраняются.
 Механизм кроссинговера
Механизм кроссинговера
 Диплотена. Хроматиды гомологичных хромосом продолжают отталкиваться друг от друга. Конфигурация бивалента зависит от числа хиазм.
Диплотена. Хроматиды гомологичных хромосом продолжают отталкиваться друг от друга. Конфигурация бивалента зависит от числа хиазм.
 Диакинез период, в котором отталкивание гомологичных хромосом продолжается, но они еще соединены в биваленты своими концами, образуя характерные фигуры - кольца и кресты. Связь между хроматидами вследствие прошедшего кроссинговера называется хиазмой. В этот период хромосомы максимально спирализованы, укорочены и утолщены. Центриоли расходятся к полюсам клетки, и в позднюю профазу происходит фрагментация кариолеммы.
Диакинез период, в котором отталкивание гомологичных хромосом продолжается, но они еще соединены в биваленты своими концами, образуя характерные фигуры - кольца и кресты. Связь между хроматидами вследствие прошедшего кроссинговера называется хиазмой. В этот период хромосомы максимально спирализованы, укорочены и утолщены. Центриоли расходятся к полюсам клетки, и в позднюю профазу происходит фрагментация кариолеммы.
 -") В оогенезе выделяют еще один период профазы I мейоза (между диплонемой и диакинезом) - диктиотену. Хромосомы представляют собой деконденсированные нити и напоминают по внешнему виду «ламповые щетки» . На этом периоде во время внутриутробного развития прерывается мейоз первичных половых клеток, синтезируется и-РНК. Мейоз возобновляется с началом созревания каждого конкретного фолликула в яичнике. Генетическая характеристика стадии - 2 n 4 c.
В оогенезе выделяют еще один период профазы I мейоза (между диплонемой и диакинезом) - диктиотену. Хромосомы представляют собой деконденсированные нити и напоминают по внешнему виду «ламповые щетки» . На этом периоде во время внутриутробного развития прерывается мейоз первичных половых клеток, синтезируется и-РНК. Мейоз возобновляется с началом созревания каждого конкретного фолликула в яичнике. Генетическая характеристика стадии - 2 n 4 c.
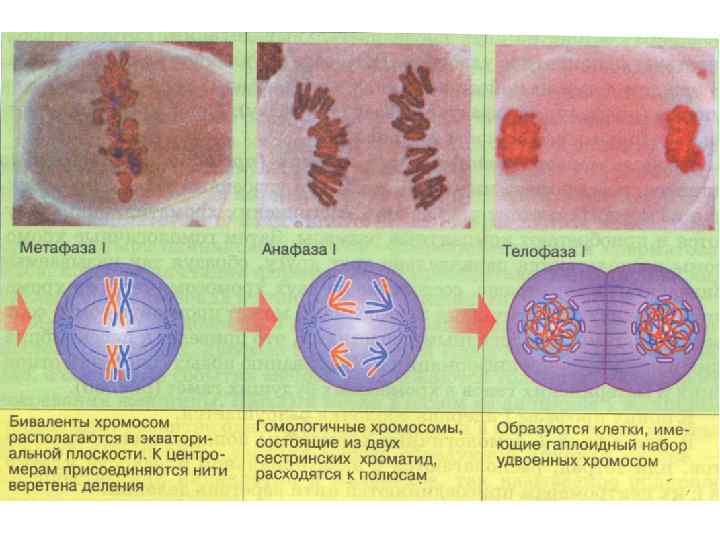
 Метафаза 1. Биваленты выстраиваются в экваториальной плоскости, образуя метафазную пластинку. Центромеры каждой из хромосом, составляющих бивалент, могут взаимодействовать только с одним из полюсов, к которому в дальнейшем и отойдут. Причем в каждом биваленте хромосомы соединяются с полюсами случайным образом, поэтому в анафазе всякий раз образуется новая комбинация хромосом, а в дальнейшем и образовавшиеся гаметы будут различны. Генетическая характеристика прежняя - 2 n 4 c.
Метафаза 1. Биваленты выстраиваются в экваториальной плоскости, образуя метафазную пластинку. Центромеры каждой из хромосом, составляющих бивалент, могут взаимодействовать только с одним из полюсов, к которому в дальнейшем и отойдут. Причем в каждом биваленте хромосомы соединяются с полюсами случайным образом, поэтому в анафазе всякий раз образуется новая комбинация хромосом, а в дальнейшем и образовавшиеся гаметы будут различны. Генетическая характеристика прежняя - 2 n 4 c.
 Анафаза 1. В результате этой фазы мейоза происходит разделение хромосом на два гаплоидных набора, попадающих в дочерние клетки. Причем в отличие от мейоза расходятся не хроматиды, а целые хромосомы. Генетическая характеристика клетки - 2 n 4 c (в целом), на полюсах - по n 2 c. Телофаза 1. Хромосомы разошлись к противоположным полюсам. Число хромосом становиться гаплоидным. Хромосомы состоят из двух хроматид. Вследствие кроссинговера хроматиды неоднородны. На короткое время образуется кариолемма. Хромосомы не деспирализуются. Далее происходит деление цитоплазмы, аналогичное тому, что наблюдается при митозе. Генетическая характеристика образовавшихся клеток n 2 c.
Анафаза 1. В результате этой фазы мейоза происходит разделение хромосом на два гаплоидных набора, попадающих в дочерние клетки. Причем в отличие от мейоза расходятся не хроматиды, а целые хромосомы. Генетическая характеристика клетки - 2 n 4 c (в целом), на полюсах - по n 2 c. Телофаза 1. Хромосомы разошлись к противоположным полюсам. Число хромосом становиться гаплоидным. Хромосомы состоят из двух хроматид. Вследствие кроссинговера хроматиды неоднородны. На короткое время образуется кариолемма. Хромосомы не деспирализуются. Далее происходит деление цитоплазмы, аналогичное тому, что наблюдается при митозе. Генетическая характеристика образовавшихся клеток n 2 c.
 Главная особенность интерфазы второго деления мейоза заключается в том, что в ней отсутствует синтетический период, то есть не происходит репликации ДНК. Это приводит в конечном итоге к появлению гаплоидного набора хромосом в образующихся клетках. Интерфаза II может и отсутствовать. Генетическая характеристика n 2 c. Второе деление мейоза сходно с митозом, однако, время профазы 2 очень короткое, поскольку хроматин конденсирован. Профаза II короткая исчезает кариолемма, центриоли перемещаются к противоположным полюсам клетки. Генетическая характеристика n 2 c.
Главная особенность интерфазы второго деления мейоза заключается в том, что в ней отсутствует синтетический период, то есть не происходит репликации ДНК. Это приводит в конечном итоге к появлению гаплоидного набора хромосом в образующихся клетках. Интерфаза II может и отсутствовать. Генетическая характеристика n 2 c. Второе деление мейоза сходно с митозом, однако, время профазы 2 очень короткое, поскольку хроматин конденсирован. Профаза II короткая исчезает кариолемма, центриоли перемещаются к противоположным полюсам клетки. Генетическая характеристика n 2 c.
 Метафаза II. Хромосомы выстраиваются по экватору клетки. При этом хроматиды случайно взаимодействуют с одним из полюсов, поэтому будут расходиться каждый раз в новых сочетаниях. Генетическая характеристика n 2 c.
Метафаза II. Хромосомы выстраиваются по экватору клетки. При этом хроматиды случайно взаимодействуют с одним из полюсов, поэтому будут расходиться каждый раз в новых сочетаниях. Генетическая характеристика n 2 c.
 Анафаза II. Происходит деление центромер, хроматиды отделяются друг от друга и теперь называются хромосомами, содержащими в целом гаплоидный набор генов. Генетическая характеристика клетки 2 n 2 c (в целом), на полюсах - nc.
Анафаза II. Происходит деление центромер, хроматиды отделяются друг от друга и теперь называются хромосомами, содержащими в целом гаплоидный набор генов. Генетическая характеристика клетки 2 n 2 c (в целом), на полюсах - nc.
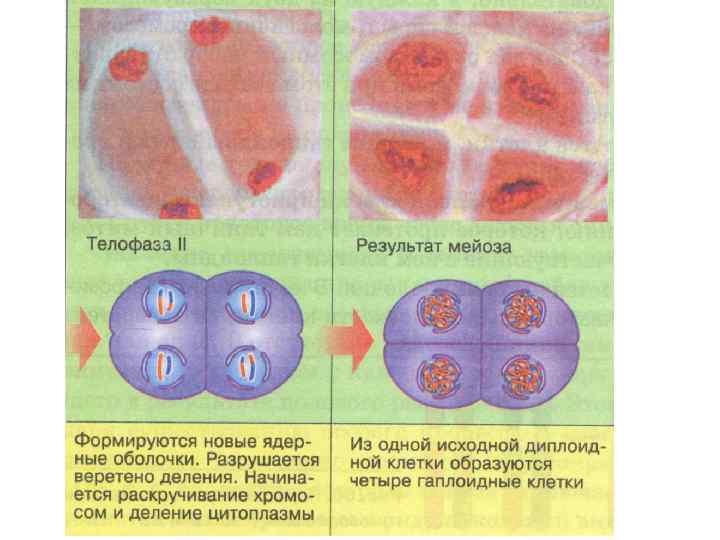
 Телофаза II Очень сходна с телофазой митоза. Генетическая характеристика образующихся клеток nc. Таким образом, из одной исходной клетки родителя образуются четыре дочерних (в случае сперматогенеза).
Телофаза II Очень сходна с телофазой митоза. Генетическая характеристика образующихся клеток nc. Таким образом, из одной исходной клетки родителя образуются четыре дочерних (в случае сперматогенеза).
 Значение мейоза: • У организмов, размножающихся половым путем, в результате мейоза образуются гаплоидные клетки, хромосомные наборы которых сливаются, формируя диплоидный набор зиготы. Если бы не было мейоза, слияние гамет приводило бы к удвоению числа хромосом в каждом новом поколении, полученном половым путем. • Мейоз создает также возможности для возникновения в гаметах новых комбинаций генов, что является причиной появления новых признаков у потомства. Этому способствуют: • случайное слияние оплодотворении; • кроссинговер в профазе первого деления мейоза; • независимое расхождение гомологичных хромосом в анафазе первого деления мейоза; • независимое расхождение хроматид в анафазе второго деления мейоза. яйцеклетки и сперматозоида при
Значение мейоза: • У организмов, размножающихся половым путем, в результате мейоза образуются гаплоидные клетки, хромосомные наборы которых сливаются, формируя диплоидный набор зиготы. Если бы не было мейоза, слияние гамет приводило бы к удвоению числа хромосом в каждом новом поколении, полученном половым путем. • Мейоз создает также возможности для возникновения в гаметах новых комбинаций генов, что является причиной появления новых признаков у потомства. Этому способствуют: • случайное слияние оплодотворении; • кроссинговер в профазе первого деления мейоза; • независимое расхождение гомологичных хромосом в анафазе первого деления мейоза; • независимое расхождение хроматид в анафазе второго деления мейоза. яйцеклетки и сперматозоида при
 Два деления мейоза
Два деления мейоза