Матричные биосинтезы.ppt
- Количество слайдов: 48



Матричные биосинтезы Лекция 1 Нуклеиновые кислоты Репликация
 в живых")
Матричным биосинтезом называется процесс поэтапного синтеза молекул биополимеров (нуклеиновые кислоты и белки) в живых организмах, когда последовательность сборки молекул запрограммирована в последовательности молекул нуклеиновых кислот (матриц)

Процессы в организме, использующие принцип матричного синтеза Процесс Матрица Продукт Функции в организме Основной фермент Репликация ДНК (обе цепи) ДНК (новая цепь) Дублирование генетической информации при делении клеток ДНК-зависимая ДНК-полимераза Транскрипция ДНК (одна из цепей) РНК Начальная стадия реализации генетической информации ДНК-зависимая РНК-полимераза Трансляция м. РНК Полипептид Формирование первичной структур белков организма Пептидилтрансфе раза Репарация ДНК (одна из цепей) ДНК (вставка в поврежденной цепи Восстановление поврежденной генетической информации ДНК-зависимая ДНК-полимераза Обратная транскрипция РНК ДНК Образование встраиваемых в ДНК хозяина копий генетической информации РНК-зависимая ДНК-полимераза

Три обязательные стадии матричных биосинтезов Инициация – процесс, в котором образуется первая связь между мономерными звеньями создаваемой полимерной цепи. Элонгация - процесс поэтапного удлинения цепи растущего полимера. Терминация - процесс остановки роста синтезируемой полимерной цепи и отсоединение ее от матрицы. При нормальном протекании матричного синтеза на каждый акт инициации и терминации приходится большое число актов элонгации.
 – высокомолекулярные соединения с линейной последовательностью мономеров")
Нуклеиновые кислоты (ДНК и РНК) – высокомолекулярные соединения с линейной последовательностью мономеров
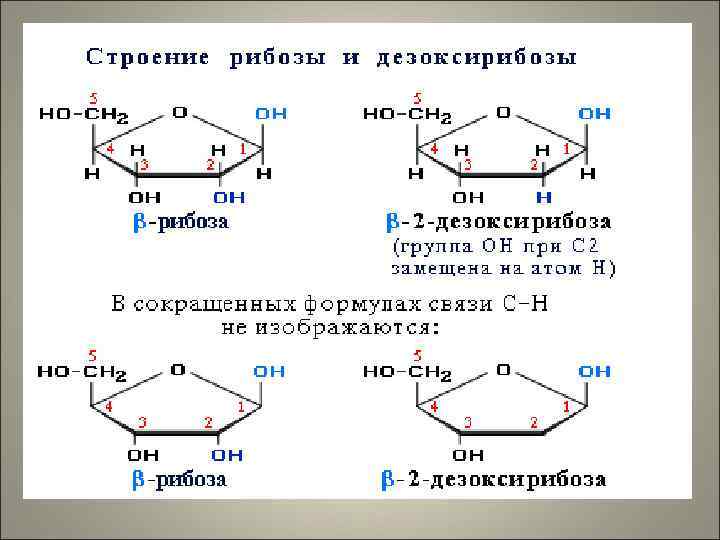
-рибоза или")
Строение нуклеотидов 1. Пуриновое или пиримидиновое азотистое основание 2. Пентоза – D(или β)-рибоза или D (или β)-2 -дезоксирибоза 3. Остатки фосфорной кислоты нуклеозид γ β α Дезоксиаденозинтрифосфат (д. АТФ) – фосфорный эфир нуклеозида

Азотистые основания — это ароматические гетероциклические соединения, производные пиримидина или пурина. Пиримидиновые основания пиримидин урацил (Ura) тимин (Thy) цитозин (Cyt) Пуриновые основания пурин аденин (Ade) гуанин (Gua)

Первичная структура ДНК и РНК • Остатки нуклеотидов соединены между собой 3’, 5’фосфодиэфирными связями (сахарофосфатный остов) • Чередование азотистых оснований нерегулярно • ДНК и РНК различаются по одному пиримидиновому основанию: (тимин в ДНК и урацил в РНК). • Молекулы нуклеиновых кислот полярны

Вторичная структура ДНК Правозакрученная спираль с общей осью, образованная двумя комплементарными, антипараллельно расположенные нуклеотидными цепями
 Уотсон, Джеймс Девей (р. 1928) Американский молекулярный")
Создатели модели структуры ДНК (1953 г. ) Уотсон, Джеймс Девей (р. 1928) Американский молекулярный биолог Крик (Crick) Фрэнсис Харри Комптон (р. 1916) Английский специалист в области молекулярной биологии Лауреаты Нобелевской премии по физиологии и медицине ( 1962 г. )

Антипараллельность Ориентация в пространстве двух полинуклеотидных цепей, образующих молекулу ДНК. 5’- конец одной из них расположен напротив 3’-конца другой

Комплементарность Соответствие азотистого основания одной цепи строго определенному азотистому основанию другой цепи Последовательность нуклеотидов в одной цепи автоматически определяет строго соответствующую ей последовательность нуклеотидов в КОМПЛЕМЕНТАРНОЙ цепи С 1’ Правилам Чаргаффа сумма ПУРИНОВЫХ оснований в молекуле ДНК всегда равна сумме ПИРИМИДИНОВЫХ оснований. • Пару составляют пуриновое и пиримидиновое снование • Тип связи -водородные • Гуанин-цитозин пары прочнее, аденин-тимин •
 взаимодействия гидрофобных азотистых оснований, расположенных перпендикулярно оси")
Силы стабилизирующие молекулу ДНК СТЭКИНГВЗАИМОДЕЙСТВИЯмежплоскостные (Ван-дер. Ваальсовы) взаимодействия гидрофобных азотистых оснований, расположенных перпендикулярно оси спирали П

Ширина большой бороздки - 2 нм, малой - 1. 2 нм, в них связываются белки

ТРЕТИЧНАЯ СТРУКТУРА ДНК Уровни структурной организации хроматина Обозначение 1 Структура 1 Двойная спираль Собственно вторичная структура ДНК 2 Нить нуклеосом Цепь ДНК, через определенное расстояние намотанная на гистоновый октамер 3 Нуклеосомное волокно Плотная однорядная упаковка нуклеосом при участии гистона Н 1 4 Соленоид Плотная трехрядная упаковка нуклеосом 5 5 Петли Спирализация соленоида, организующаяся при участии негистоновых белков хроматина и белков ядерного скелета 6 6 Мини-диски Структурное объединение примерно 20 петель 7 7 Хромосома Плотная стопчатая упаковка минидисков 2 3 4
")
Гистоны (Белки богатые лизином и аргинином)

Химическая модификация гистонов

Модификация Суть модификации Аминокислотные остатки-мишени Биологическая роль Фосфорилирование Присоединение остатков ортофосфорной кислоты Ser Стимулирует конденсацию хроматина Ацетилирование Присоединение остатка уксусной кислоты Lys Стимулирует разворачивание хроматина Убихитинилирование (только для Н 2 а) Ковалентное связывание с низкомолекулярным белком убихитином ? Стимулирует разворачивание хроматина

• Репликация ДНК осуществляется при делении клеток и лежит в основе передачи наследственных признаков • Общие принципы репликации ДНК применимы, с небольшими модификациями, ко всем клеткам. Репликация - процесс образования идентичных копий ДНК

Принципы репликации ДНК 1. Комплементарность 2. Полуконсервативность

Принципы репликации ДНК 3. Антипаралельность 4. Униполярность. Синтез каждой дочерней цепи ДНК происходит антипараллельно матричной цепи и всегда в направлении 5' → 3'

Принципы репликации ДНК 5. Согласованность репликации и клеточного деления 6. Двунаправленность от одной специфической точки начала репликации (в большинстве случаев).
 •")
Основные события и особенности этапов репликации Инициация • Формирование репликативной вилки (создание матрицы) • Образование праймера Элонгация • Ассиметричный синтез ДНК Терминация • Вырезание праймеров • Сшивание отдельных фрагментов цепи ДНК
")
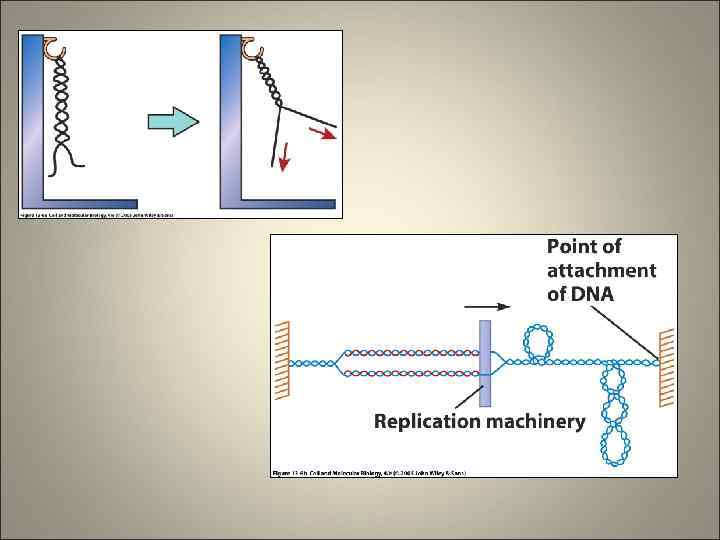
Формирование репликативной вилки (матрицы)

ИНИЦИАЦИЯ РЕПЛИКАЦИИ Этапы Внешняя инициация репликации Регуляция сверхспирализации Стабилизация участков ДНК одноцепочечных Образование праймера Ферменты, белки факторы роста Хеликаза топоизомеразы Процессы Определение точек начала репликации , состоящих приблизительно из 300 нуклеотидов Локальная денатурация ДНК Расплетение спирали ДНК (разрыв водородных связей) Ослабление сверхспирализации SSB белки Поддержание реплицируемого участка ДНК в раскрученном состоянии ДНК-зависимая РНКполимераза: у эукариот –полимераза а, у прокариот - праймаза Соединение нескольких РНК мономеров

Структура и механизм действия хеликазы

Топоизомеразы типа I • временно разрезают одну из двух цепей ДНК • 5’-фосфат разрезанной цепи ковалентно прикрепляется к Tyr фермента • Разорванная нить раскручивается, и цепь зашивается • Энергии извне не требуется

• ДНК –полимеразы не могут начинать синтез ДНК на матрице, а способны только добавлять новые дезоксирибонуклеотидны к 3’-концу полинуклеотидной цепи. • Образованная цепь , к которой добавляются дезоксирибонуклеотиды, называется затравкой (от анг. рrimer –затравка) и состоит из менее 20 рибонуклеотидов.
 - образец, по которому строится")
ЭЛОНГАЦИЯ РЕПЛИКАЦИИ Основные компоненты элонгации 1. Цепи ДНК (матрица) - образец, по которому строится новая цепь ДНК. 2. Нуклеотиды (d. АТФ, d. ГТФ, d. ТТФ, d. ЦТФ) - субстраты из которых строятся дочерние цепи. 3. ДНК-полимераза – фермент, осуществляющий синтез новой цепи ДНК.

Ферментативная реакция, катализируемая ДНКполимеразой При каждом цикле элонгации на начальной его стадия происходит выбор такого субстрата, который соответствует считываемому в данный момент кодовому сигналу на молекуле матриц и только после этого формируется 3’, 5’-фосфодиэфирная связь

Ассиметричный синтез ДНК При синтезе новой молекулы ДНК выделяют лидирующую и запаздывающую цепи. У лидирующей цепи направление синтеза совпадает с движением репликативной вилки • ДНК-полимераза синтезирует запаздывающую цепь малыми фрагментами (фрагменты Оказаки), которые затем должны быть сшиты ферментом ДНК-лигазой. • • • Размеры фрагментов Оказаки: фаги – 1000 -2000 н, E. coli – 1000 н эукариоты – 200 -400 н.

Структура ДНК полимеразы эукариот Полимераза - синтез праймеров Полимераза - синтез лидирующей цепи Полимераза - синтез отстающей цепи Полимераза - удаление РНК праймеров и достраивание ДНК

Механизм ассиметричного синтеза ДНК

Ферменты и белки, участвующие в репликации Эукариоты E. coli Раскручивание двойной спирали в репликативной вилке Хеликаза Подержание участка ДНК в раскрученном состоянии SSB-белки Устранение свехспирализации Семейство Топоизомераз ( I, III) Гидраза Синтез праймеров (инициация) Полимераза Праймаза Элонгация Полимераза (лидирующая цепь) Полимераза (запаздывающая цепь) Полимераза III Удаление РНК праймеров и достраивание ДНК Полимераза I Соединение разрывов в цепи ДНК-лигаза

Проблема «недорепликации» концов линейных молекул Удаление крайних РНКпраймеров, комплементарных 3’концам обеих цепей линейной материнской ДНК приводит к тому , что дочерние цепи оказываются короче на 10 -20 нуклеотидов (величина затравки).

Структура теломер
")
Достраивание выступающих однонитевых 3'-концов линейных молекул ДНК с помощью теломеразы (обратной транскриптазы)

Репарация генетических повреждений – свойство живых организмов восстанавливать повреждения, возникающие в ДНК спонтанно или в результате воздействия разнообразных повреждающих факторов.

Повреждения ДНК и их причины

Основные репарабельные повреждения в ДНК Апуринизация Разрывается N-гликозидная связь между пуриновым основанием и дезоксирибозой. Циклобутановые димеры Под действием ультрафиолетового света происходит ковалентное сшивание рядом стоящих пиримидинов. При сшивании тиминов образуется циклобутановое производное, блокирующее репликацию Дезаминирование Аденин превращается в гипоксантин Гуанин - в ксантин Цитозин в урацил.
 Фермент фотолиаза расщепляет вновь")
Основные принципы различных реакций прямой репарации Фотореактивация (прямая репарация димеров) Фермент фотолиаза расщепляет вновь образовавшиеся связи между соседними пиримидиновыми основаниями и восстанавливает нативную структуру. Репарация АП-сайтов за счет прямой вставки азотистых оснований Ферменты инсертазы, которые могут вставлять в брешь такое же основание, которое было до поражения и соединять его с дезоксирибозой. Структура ДНК приобретает исходный неповрежденный вид.

Общая схема эксцизионной репарации от анг. excision - вырезание Вырезание поврежденных участков из цепи ДНК с последующим застраиванием образовавшейся бреши неповрежденными нуклеотидами

Пострепликативная репарация модифицированных оснований Вид модификации – превращение цитозина в урацил в результате дезаминирования Ферменты репарации: ДНК N-гликозилаза гидролизует связи между аномальным основанием и дезоксирибозой (образуется АПсайт) АП-специфическая эндонуклеаза отщепляет от ДНК сахарофосфатную группу, лишенную основания. ДНК-полимеразой заделывает брешь размером в один нуклеотид. Лигаза сшивает концы ДНК.
 репарация некомплементарных оснований")
Пострепликативная (метилзависимая) репарация некомплементарных оснований

Пострепликативная репарация индуцированных повреждений
Матричные биосинтезы.ppt