Лютеинизирующий гормон реф.pptx
- Количество слайдов: 21



Лютеинизирующий гормон Работу выполнила студентка 4 курса 1 группы Ветеринарнобиологического факультета Грефенштейн Мария

История открытия гормона 1927 г - Бернгард Цондек и Зельмар Ашгейм Им удалось пересадить молодым мышам самкам передние доли гипофиза и вызвать у них преждевременное половое созревание. Это взволновало весь ученый мир, открытие было достойно нобелевской премии. Некоторое время спустя те же исследователи открыли в моче беременных женщин гормоны, названные ими пролан А и пролан В. Хотя это не половые гормоны, но они управляют половыми органами и называются поэтому гонадотропами, что означает гормоны, воздействующие на половые железы
, гормон передней доли гипофиза. Представляет собой гликопротеин,")
Химическая природа гормона Лютеинизирующий гормон (лютропин, ЛГ), гормон передней доли гипофиза. Представляет собой гликопротеин, молекула которого построена из 2 субъединиц - α (содержит 89 -96 аминокислотных остатков) и β (115 -119 аминокислотных остатков), нековалентно связанных друг с другом; молекулярная масса не более 30 к. Да. Для субъединиц лютеинизирующего гормона характерно высокое содержание остатков пролина и дисульфидных связей между остатками цистеина. Олигосахаридные цепи в лютеинизирующем гормоне(их молекулярная масса составляет 16% от молекулярной массы всей молекулы) присоединены к аспарагину с помощью N-гликозидных связей, причем α -субъединица содержит 2 олигосахаридные цепи, β - одну. В их состав входят L-фруктоза, D-галактоза, D-манноза, N-ацетилглюкозамин и Nацетилгалактозамин.

Изолированные α- и β-субъединицы не обладают биологической активностью. Дегликозилированный лютеинизирующий гормон значительно теряет биологическую активность и проявляет свойства физиологического антагониста нативного гормона. Ген, кодирующий α-субъединицу локализован в длинном плече шестой хромосомы (6 q 12. 21). Ген, кодирующий структуру β-субъединицы локализован в скоплении генов LHB/CGB длинного плеча 19 -й хромосомы (19 q 13. 32. ). В отличие от альфа-гена экспрессия гена бета-субъединицы ограничена гонадотропными клетками гипофиза. Активность гена регулируется гипоталамическим гонадотропинрилизинг гормоном.


Биохимические функции и молекулярный механизм действия ЛГ Двумя гипофизарными гормонами, регулирующими функцию половых желез, являются фолликулостимулирующий гормон (ФСГ), который стимулирует рост фолликулов яичника, а также рост яичек и сперматогенез, и лютеинизирующий гормон (ЛГ), который вызывает овуляцию и лютеинизацию яичниковых фолликулов, стимулирует функцию интерстициальных (лейдиговских) клеток яичек и повышает продукцию стероидов как яичниками, так и яичками.

Гонадотропины оказывают биологический эффект на тканимишени посредством активации специфических рецепторов, связаных с мембраной клеток-мишеней. Поверхностные клеточные рецепторы гормона являются белками, входящими в структуру клеточной мембраны. После связывания с гонадотропином мембранные рецепторы стимулируют выработку растворимых внутриклеточных мессенджеров, которые, в свою очередь, обеспечивают клеточный ответ. Клетками-мишенями ЛГ являются клетки Лейдига, в которых стимулируется продукция андрогенов, особенно тестостерона. Рецепторы к ЛГ так же локализуются на лютеальных клетках, и влияние ЛГ опосредуется через стимуляцию аденилатциклазы и внутриклеточного повышения уровня ц. АМФ, который непосредственно или через посредников (протеинкиназа и др. ) активирует ферменты, участвующие в биосинтезе прогестерона.

Под влиянием ЛГ в яичниках и яичках увеличивается количество холестерина, необходимого для синтеза гормонов. Одновременно повышается активность ферментов семейства цитохромов Р 450, отщепляющего боковую цепь в молекуле холестерина. При более длительном влиянии ЛГ стимулирует экспрессию и синтез других ферментов (Зbгидроксистероидная дегидрогеназа, 17 а-гидроксилаза), участвующих в синтезе прогестерона и других стероидов. Таким образом, в желтом теле под влиянием ЛГ усиливаются процессы стероидогенеза на участке конверсии холестерина в прегненолон.

Биосинтез и секреция ЛГ Биосинтез лютеинизирующего гормона осуществляется из α - и β -субъединиц, которые образуются раздельно из соответствующих высокомолекулярных белковпредшественников, аминокислотная последовательность которых кодируется различными генами. Образование "зрелых" полипептидных цепей происходит в результате специфического протеолитического расщепления белковпредшественников. ЛГ и ФСГ — синтезируются гонадотрофами (клетками, расположеными в глубине латеральной части передней доли гипофиза). На поверхности гонадотрофов имеются рецепторы к Гн-РГ(гонадотропный релизинг-гормон), плотность которых зависит от уровня стероидных гормонов в крови и от концентрации Гн-РГ.

Существуют 3 типа секреции гонадотропинов: тонический, циклический и эпизодический, или пульсирующий. Тоническая или базальная секреция угнетается по механизму обратной негативной связи периферическими гормонами. Циклическая - стимулируется по механизму обратной позитивной связи периферическими гормонами. Периодическая (пульсирующая) секреция осуществляется в ответ на выделение гонадолиберинов гипоталамуса каждые 90 -120 мин. Пульсирующая секреция обусловлена активностью гипоталамуса и высвобождением гонадолиберинов.

Регуляция секреции Регуляторами выработки ФСГ и ЛГ, помимо гипоталамических либеринов являются ингибин и активин, которые продуцируются гранулёзными и лютеиновыми клетками яичников, а также клетками цитотрофобласта и клетками Сертоли семенников, также регуляция осуществляется половыми гормонами (эстрогены, тестостерон). Регуляция секреции гонадотропинов обеспечивается за счёт "короткой" и "ультракороткой" цепей обратной связи. Так, повышение уровня ЛГ и ФСГ приводит к торможению их синтеза и высвобождения, а повышенная концентрация гонадолиберина в гипоталамусе угнетает его синтез и высвобождение в портальную систему гипофиза.

На высвобождение гонадолиберина оказывают влияние также катехоламины: дофамин, адреналин и норадреналин. Адреналин и норадреналин стимулируют высвобождение гонадолиберина, тогда как дофамин оказывает такое же действие только у животных, которым предварительно вводились стероидные гормоны. Холецистокинин, гастрин, нейротензин, опиоиды и соматостатин угнетают высвобождение гонадолиберина. Агонисты гонадолиберина (бузерилин, нафарелин, леупролид и др. ) оказывают свой эффект посредством взаимодействия с теми же мембранными рецепторами. Пролактин угнетает продукцию гонадотропных гормонов. Угнетающее действие на выделение ЛГ оказывают также глюкокортикоиды.

Эстрогены повышают чувствительность гонадотрофов к эффекту Гн. РГ; кульминацией всех этих влияний является овуляторный выброс ЛГ и ФСГ. Прогестерон, секретируемый желтым телом, в присутствии эстрогенов также усиливает реакцию гонадотропинов на Гн. РГ, но, по-видимому, оказывает тормозное влияние на гипоталамус по механизму отрицательной обратной связи. Эффект эстрогенов в системе обратной связи гораздо сложнее, поскольку в зависимости от дозы и длительности воздействия эстрогенов можно наблюдать как стимулирующие, так и тормозные влияния на гипофиз. Кратковременное воздействие эстрогенов снижает как исходную секрецию ЛГ, так и реакцию ЛГ на Гн. РГ. При более длительном воздействии эстрогенов, особенно в относительно малых дозах, реакция гонадотропинов усиливается. У мужчин тестостерон и эстрогены оказывают тормозящее действие на выработку ЛГ.
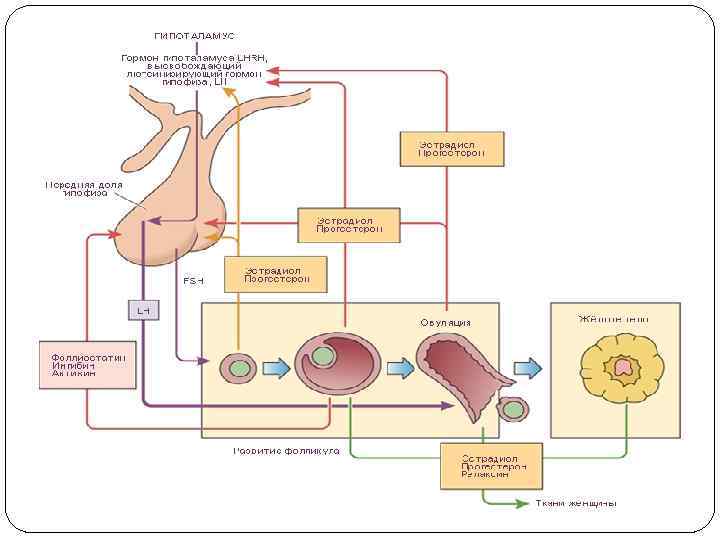

Транспорт лютеинизирующего гормона осуществляется кровью и лимфой. Период полураспада гонадотропинов, циркулирующих в крови, имеет прямое отношение к компоненту сиаловых кислот в молекуле гормона. Доказано, что десиалирование укорачивает период полураспада и биологическую активность гонадотропинов. ФСГ находится в крови в свободной форме и период его полураспада составляет 55 -60 мин, а ЛГ — 25 -30 мин. В репродуктивном возрасте ежедневное высвобождение ЛГ составляет 500 -1100 м. МЕ, в постменопаузе скорость образования ЛГ увеличивается и его количество составляет до 3000 -3500 м. МЕ в день. Скорость секреции ЛГ у взрослых лиц с нормальной функцией половых желез составляет 500— 1000 м. ЕД/сут, а ФСГ — 50— 100 м. ЕД/сут.

Гормональная регуляция менструального цикла Менструальный цикл - это промежуток времени между двумя менструальными выделениями крови (от первого дня предыдущей менструации до первого дня последующей). Длительность менструального цикла 21 -36 дней (у 60 % женщин 28 дней, у 28 % - 21 -27 дней, у 12 % 30 -36 дней). Секреция гормонов яичником пребывает под гипофизарным контролем. Синтез эстрогенов и созревание фолликула определяются стимулированием фолитропином (ФСГ). Одновременно стимулируется 10 -15 фолликулов, созревает 1, остальные подвергаются атрезии. Эстрогены усиливают связь фолитропина с рецепторами, способствуют накоплению его в фолликуле, что стимулирует дальнейшую продукцию эстрогенов и созревание фолликула.

Пик эстрадиола стимулирует образование гонадолиберинов в гипоталамусе с последующей стимуляцией выделения ЛГ и ФСГ гипофизом. Вследствие этого стимулируется разрыв фолликула и овуляция яйцеклетки. На этом заканчивается фолликулярная фаза менструального цикла и начинается лютеиновая. Полость лопнувшего фолликула быстро зарастает гранулезными клетками, заполненными желтым пигментом лютеином. Формируется капиллярная сеть, выделяется прогестерон. Эта фаза (фаза желтого тела) продолжается 7 дней.

Прогестерон инактивирует механизмы активной секреции гонадотропинов, их концентрация уменьшается, достигая минимума в середине лютеиновой фазы. Если не произошло оплодотворение, начинается регресс желтого тела. Это обусловлено снижением концентрации гонадотропинов и низкой чувствительностью к ним лютеиновых клеток. Лютеолиз усиливается простагландинами, выделяемыми маткой. Секреция прогестерона уменьшается и через 1 -2 дня начинается менструация с отслоением функционального слоя эндометрия. Во время менструации уменьшается секреция эстрогенов, прогестерона, что побуждает к секреции ФСГ и ЛГ и началу нового менструального цикла.

Эндокринные патологии, связанные с ЛГ Упорно высокие уровни ЛГ свидетельствуют о ситуации, когда происходит нарушение нормальной отрицательной обратной связи между гонадами и гипоталамусом, ведущее к растормаживанию гипофизарной продукции ЛГ и ФСГ. Такое нормально во время менопаузы, но является отклонением от нормы во время репродуктивного периода. Это может свидетельствовать о таких состояниях как: преждевременная менопауза; дизгенезия гонад, синдром Тернера ( хромосомная болезнь, сопровождающаяся характерными аномалиями физического развития, низкорослостью и половым инфантилизмом. Моносомия по Х-хромосоме (ХО)) кастрация; синдром Суайра (наследственная изолированная гонадальная дисгенезия); некоторые формы врождённой гиперплазии надпочечников; гипофункция яичников. Опухоли гипофиза, секретирующие ЛГ

Пониженная секреция ЛГ может приводить к гипогонадизму, который у мужчин обычно проявляется снижением количества сперматозоидов. У женщин, как правило, наблюдается аменорея. С низким уровнем ЛГ могут протекать такие состояния как: • гипоталамические опухоли, травмы; • наследственные заболевания: -синдром Кальманна -синдром Прадера — Вилли (отсутствуют или не экспрессируются примерно 7 генов из 15 -й хромосомы, унаследованной от отца. )и др. • гипопитуитаризм (вторичному гипогонадизму) • функциональные нарушения -расстройства питания -гиперпролактинемия -аменорея спортсменок • гонадосупрессивная терапия -антагонистами гонадолиберина, -агонистами гонадолиберина (отрицательная регуляция).

Спасибо за внимание!
Лютеинизирующий гормон реф.pptx