Эл.генез. 2-3. Законы тока.ppt
- Количество слайдов: 72



ЛОКАЛЬНЫЙ ОТВЕТ. ЗАКОНЫ ДЕЙСТВИЯ ПОСТОЯННОГО ТОКА. Проведение возбуждения по нерву.

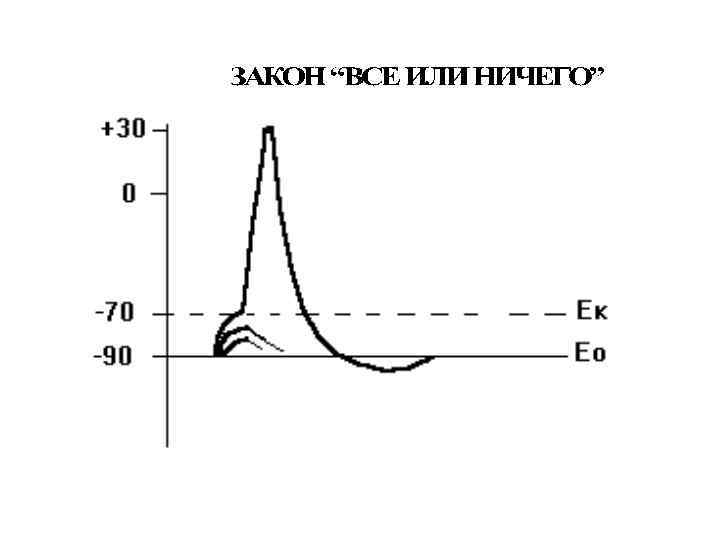
Отличия локального ответа от потенциала действия: Локальный ответ Eк Eo 1. Возникает на подпороговые раздражители. 2. Не распространяется. 3. Не подчиняется правилу “Все или ничего”. 4. Способен к суммации. 5. Возбудимость в период локального ответа повышена. Локальный ответ возникает при действии силы раздражения от 50 до 75 % от уровня критической деполризации. Если локальный ответ достигнет уровня критической деполяризации, он перерастает в потенциал действия.

Основные электрофизиологические феномены в нервном волокне Аэт — анаэлектротон, КУД — критический уровень деполяризации, Кэт — катэлектротон, ЛО — локальный (подпороговый активный) ответ, МПП — мембранный потенциал покоя, ПД — потенциал действия, СП (отр и пол) — следовые потенциалы отрицательный и положительный (временные соотношения пика ПД и СП не выдержаны; отрицательный СП и особенно положительный СП значительно длительнее).

Локальные ответы и закон силы
 -75 м.")
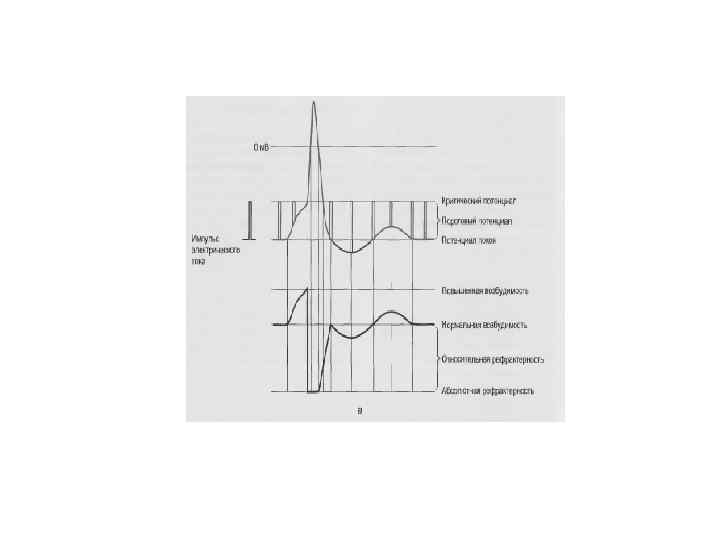
СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИ С ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ 0 Екр( -60 м. В) -75 м. В

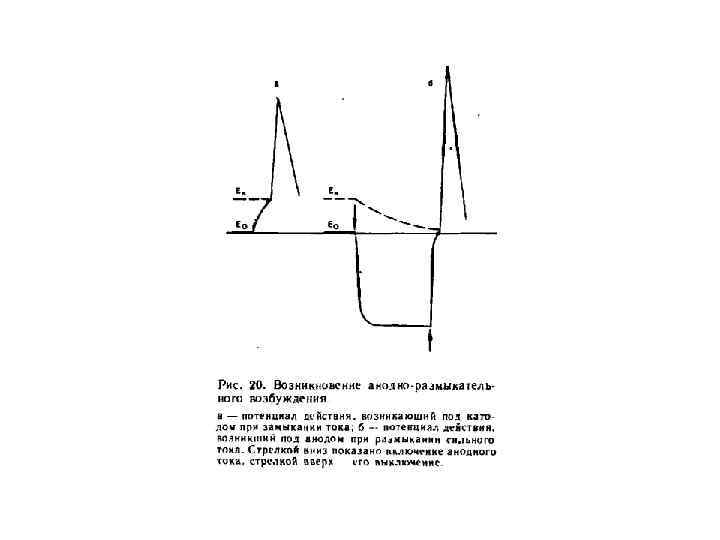
Полярный закон Пфлюгера Возбуждение возникает в момент замыкания цепи под катодом, а в момент размыкания цепи под анодом.

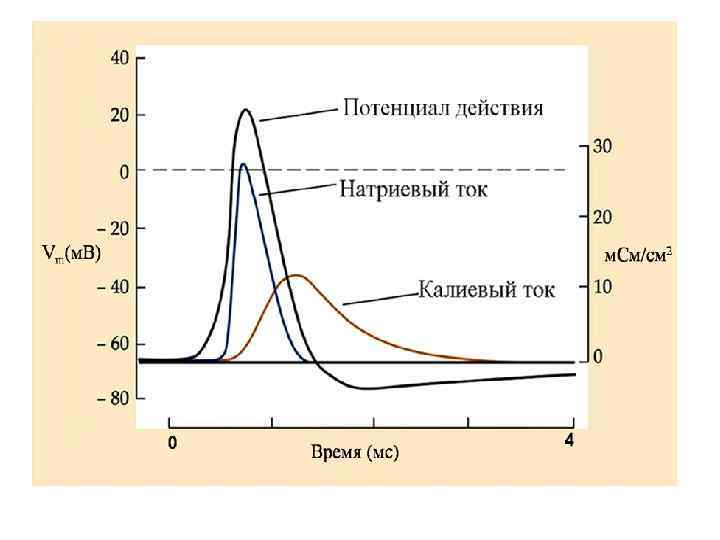
Полярный закон действия тока – возбуждение возникает под катодом при замыкании и под анодом при размыкании тока Цепь событий, развивающихся под катодом раздражающего тока: • • пассивная деполяризация мембраны повышение натриевой проницаемости усиление потока Na++ внутрь волокна активная деполяризация мембраны локальный ответ достижение критического уровня (Ек ) регенеративная деполяризация потенциал действия (ПД).

Пассивные изменения КАТОД АНОД «-» «+» ----------++++ -----
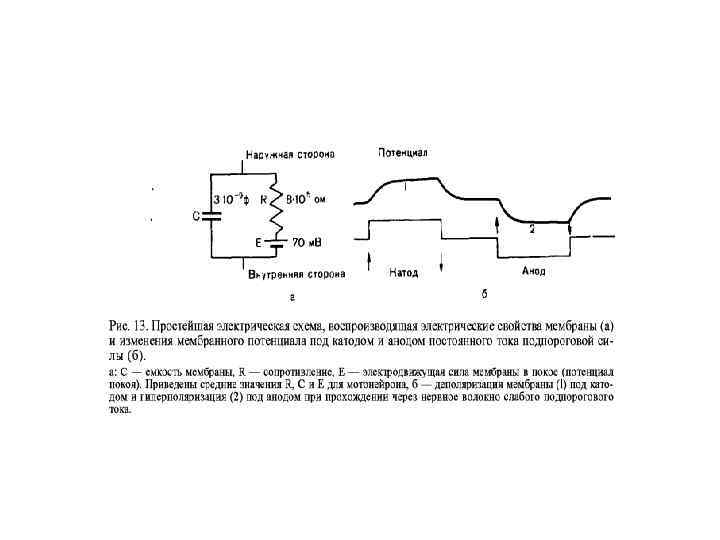

Закон физиологического электротона В момент замыкания цепи возбудимость и проводимость под катодом увеличиваются – катэлектротон; а под анодом – уменьшаются – анэлектротон; При размыкании цепи возбудимость под катодом уменьшается – обратный катэлектротон; а под анодом – увеличивается – обратный анэлектротон.

Физиологический электротон
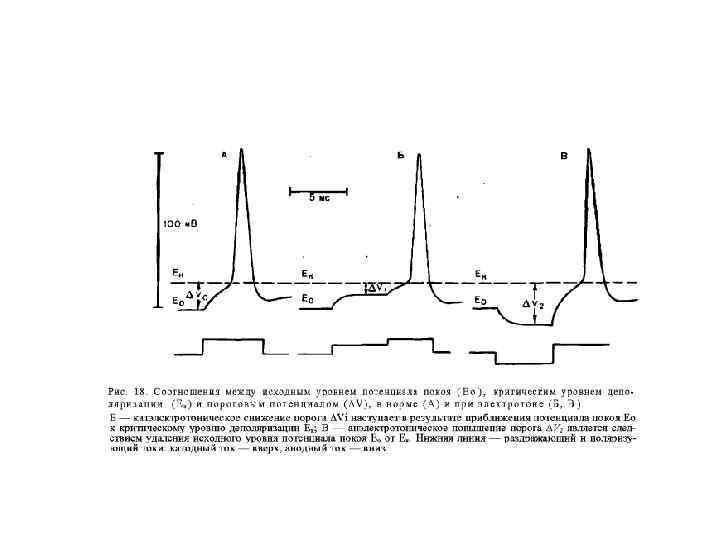

Изменения возбудимости при длительном действии катода Катодическая депрессия Вериго Катодзамыкательное возбуждение Ек-2 Ек-1 Ео замыкание размыкание
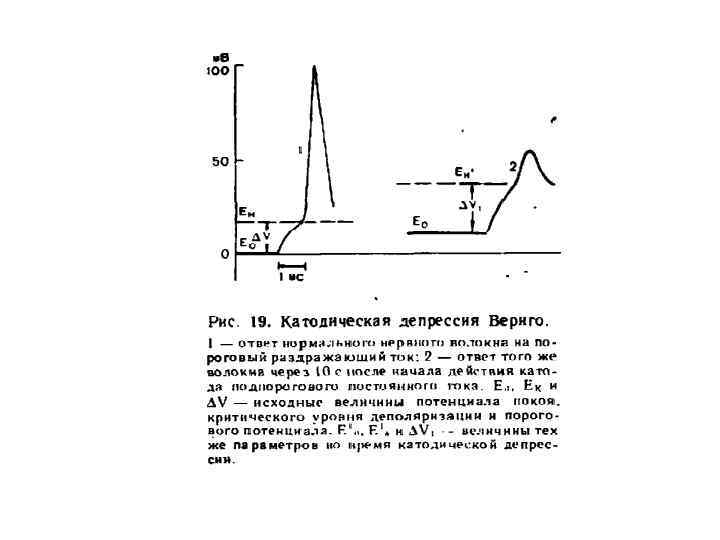

Изменения возбудимости при длительном действии анода Ек-1 Ек-2 Ео замыкание размыкание

ПОТЕНЦИАЛ ПОКОЯ АНОД ВКЛЮЧЕН АНОД ВЫКЛЮЧЕН Увеличение числа реактивированных Nа+-каналов Выходящий К+-ток значительно превышает входящий Na+-ток Уменьшение количества открытых К+-каналов Входящий Nа+-ток превышает Выходящий К+-ток

m m h h Р Е А К Т И В А Ц И Я Na-канала
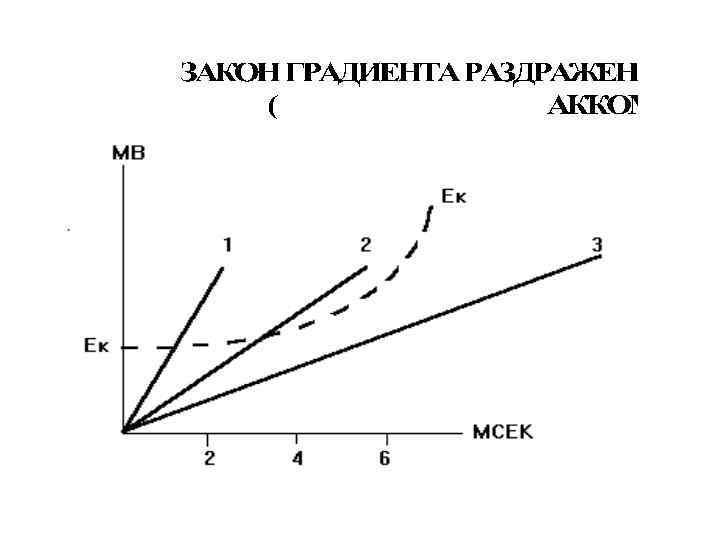
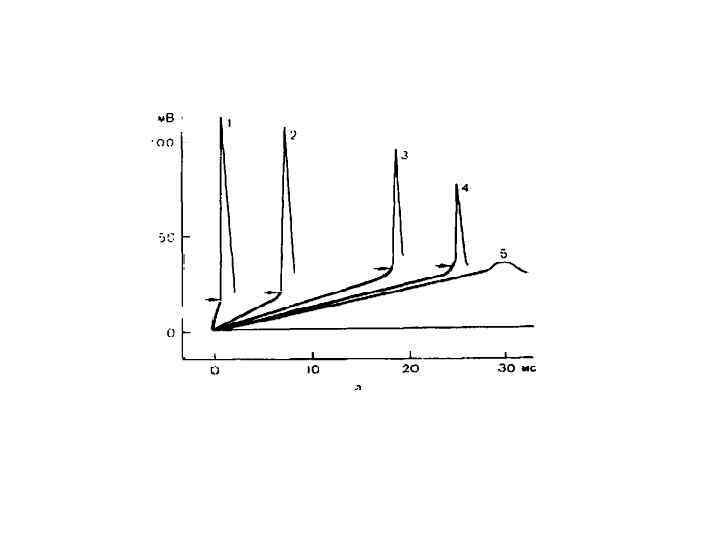

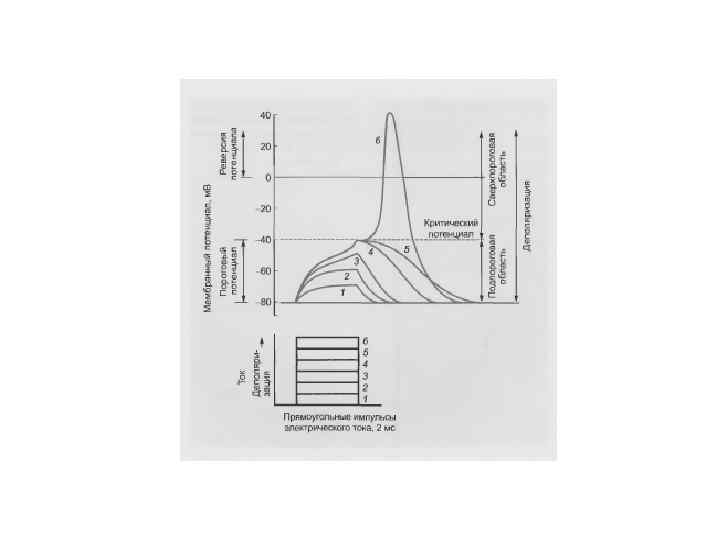
Критический наклон равен отношению реобазы тока с минимальной скоростью нарастания силы раздражителя к реобазе прямоугольного толчка тока КН = Реобаза

Парабиоз Местное нераспростроняющееся и углубляющееся во времени возбуждение. Явление парабиоза обнаружено Введенским при исследовании способности нерва проводить высокочастотные разряды импульсов после воздействия на нерв различных химических агентов. В дальнейшем было показано, что состояние парабиоза можно вызвать действием раздражителя любой природы.

Парабиоз и его фазы • Уравнительная • Парадоксальная • Тормозная нерв NH 4
")
сила раздражения (Гц)
 -75 м.")
СООТНОШЕНИЕ ФАЗ ВОЗБУДИМОСТИ С ФАЗАМИ ПОТЕНЦИАЛА ДЕЙСТВИЯ 0 Екр( -60 м. В) -75 м. В

• Введенский впервые обратил внимание на то, что способность ткани воспроизводить задаваемый ритм связана с ее функциональным состоянием – лабильностью. • Явление парабиоза широко распространено в природе – зимняя спячка (анабиоз), как хладнокровных, так и теплокровных (медведи). • Использование наркотических веществ позволяет проводить многочасовые операции, а анестетики – блокируют болевые ощущения. • Открытие парабиоза, а затем пессимального торможения, позволило Введенскому выдвинуть положение о тормозных явлениях в нервах, как о стойком нераспростроняющемся возбуждении.

Образование миелинового волокна

Миелиновое нервное волокно миелин перехваты Ранвье

Скорость передачи сигнала по нервным волокнам
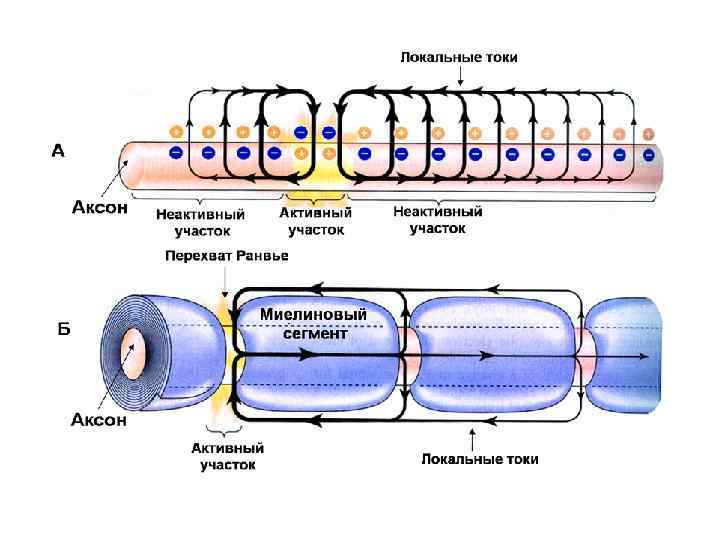

В основе распространения возбуждения - возникновение местных токов между деполяризованным и покоящимся участками. Между различно заряженными участками мембраны возникает локальный ионный ток, который деполяризует мембрану до критического уровня. Причем подпороговый деполяризующий ток идет по аксоплазме, то есть - изнутри. В результате этой подпороговой деполяризации открываются Nа-каналы и возрастает входящий Nа ток. Отношение величины ПД к пороговому току называется фактором надежности: Амплитуда ПД, м. В Величина Екр, м. В • Плотность Nа+-каналов в перехватах Ранвье: 10 000/мм 2 , что в 200 раз больше, чем нервном волокне гигатского аксона кальмара. • Входящий Nа+-ток, пронизывающий невозбужденную мембрану в непосредственной близости от ее возбужденного участка в 5 -6 раз выше порогового тока. То есть фактор надежности настолько высок, что позволяет перескакивать ПД через несколько перехватов Ранвье, при их блокаде анестетиками.
 – мякотные толстые моторные")
Классификация нервных волокон • Волокна типа А (ά, β, δ) – мякотные толстые моторные волокна, скорость проведения возбуждения до 120 м/сек. • Волокна типа В –тонкие мякотные волокна, чаще чувствительные, скорость проведения 3 -18 м/сек. • Волокна типа С – безмякотные, вегетативные, скорость проведения не больше 3 м/сек.
 Миелинизация Скорость проведения")
Типы нервных волокон, их свойства и функциональное назначение Тип Диаметр (мкм) Миелинизация Скорость проведения (м/с) Функциональное назначение А 12– 20 сильная 70– 120 Двигательные волокна соматической НС; чувствительные волокна проприорецепторов А 5– 12 сильная 30– 70 Чувствительные волокна кожных рецепторов А 3– 16 сильная 15– 30 Чувствительные волокна проприорецепторов А 2– 5 сильная 12– 30 Чувствительные волокна терморецепторов, ноцицепторов В 1– 3 слабая 3– 15 Преганглионарные волокна симпатической НС С 0, 3– 1, 3 отсутствует 0, 5– 2, 3 Постганглионарные волокна симпатической НС; чувствительные волокна терморецепторов, ноцицепторов, некоторых механорецепторов

• Составной потенциал действия подкожного нерва млекопитающего, зарегистрированный с помощью внеклеточного электрода. Все волокна нерва одновременно подвергаются стимуляции на некотором расстоянии от участка отведения. До участка отведения в первую очередь доходят потенциалы действия наиболее быстро проводящих волокон группы А; потенциалы действия медленных волокон группы С появляются примерно через 38 мс. Вслед за колебанием потенциала, соответствующим волокнам группы С, возникает продолжительный гиперполяризующий следовой потенциал. Отдельные зубцы волны, соответствующей А–волокнам, отражают активность волокон подгрупп α, β, γ, δ

Законы проведения возбуждения 1. 2. 3. 4. Закон физиологической непрерывности Закон двустороннего проведения Закон изолированного проведения Закон бездекрементного (незатухающего) проведения возбуждения 5. Закон относительной неутомляемости нерва (открыт Введенским)
 на электрическом органе нильского сома.")
Двустороннее проведение возбуждения экспериментально доказано: Бабухиным А. И. (1877) на электрическом органе нильского сома. Кюне В. (1886) на икроножной мышце лягушки.

Опыт Бабухина А. И. 1 2 3 4 5 6

Опыт Бабухина А. И. 1 2 3 разрез 4 разрез 5 6 раздражение

Опыт Кюне В. разрез ие ражен разд

Синапс

Связи между нейронами осуществляются посредством синапсов. Синапс это структура, посредством которой обеспечивается передача информации между нервными клетками, нервными и мышечными клетками. Термин предложен Ч. Шеррингтоном в 1897 г. В переводе с греческого означает – смыкать.

Пути передачи информации

Расположение синапсов на теле нейрона и его дендритах
 центральные (головной и")
КЛАССИФИКАЦИЯ СИНАПСОВ • • • • 1. По местоположению: а. ) центральные (головной и спинной мозг) - аксосоматические, аксональные, аксодендретические; - дендросоматические, дендродендретические. б. ) переферические ( нервно-мышечные, нейросекреторные). 2. По характеру действия: а. ) возбуждающие б. ) тормозные 3. ) По способу передачи сигнала: а. ) электрические; б. ) химические; в. ) смешанные. 4. ) По развитию в онтогенезе: а. ) стабильные (синапсы дуг безусловного рефлекса); б. ) динамичные (появляются в процессе развития индивидуума).
 По природе медиатора химические делятся на следующие группы: I. Группа биогенных аминов:")
5. ) По природе медиатора химические делятся на следующие группы: I. Группа биогенных аминов: а. ) катехоламины – норадреналин, дофамин. б. ) холинергические – ацетилхолин. II. Группа аминокислот: глутаминовая кислота (глутамат), аспарагиновая кислота (аспартат), γ-аминомаслянная кислота (ГАМК). III. Пептидергические а. ) опиоидные (энкефалины, эндорфины), тахикинины (вещество Р, нейрокинин А), вазопрессин и др. IY. Пуринергические: АТФ, аденозин. Y. Газы: NO, CO, SH 2

Локализация синапсов разного типа

Для ЦНС характерен широкий спектр медиаторов: АХ, НА, ГАМК, АТФ, дофамин, серотонин, пептиды и т. д. Функционально делятся на тормозные и возбуждающие. Принцип Дейла (одним нейроном выделяется один медиатор) в настоящее время пересмотрен.


Строениехимического синапса: • пресинаптическая мембрана, • постсинаптическая мембрана, • синаптическая щель В синаптическом окончании содержится много митохондрий и синаптических пузырьков. В каждом пузырьке - тысячи молекул медиатора (например, ацетилхолина) и молекулы АТФ.

Строение химического синапса • Пресинаптическая мембрана образована аксональным окончанием, которое теряет в этом месте миелиновую оболочку. Здесь содержаться синаптические пузырьки, диаметром 30 -50 нм и многочисленные митохондрии. Синаптические везикулы содержат медитор и АТФ (составляющие квант медиатора), имеют отрицательный заряд и отталкиваются от пресинаптической мембраны, сосредоточены везикул в «активных зонах» . • Синаптические пузырьки находятся в нескольких фракциях – резервном и рециркулирующем пуле. Выделяется порционно – квантами. • Ширина синаптической щели составляет 20 -50 нм. Она заполнена межклеточной жидкостью и содержит структурные элементы: базальную мембрану, состоящую из фиброзных волокон, которые соединяют между собой пре- и постсинаптические мембраны. Здесь же находятся ферменты, разрушающие молекулы медиатора.
 имеет многочисленные складки, увеличивающие площадь ее взаимодействия")
• Постсинаптическая мембрана (или концевая пластинка) имеет многочисленные складки, увеличивающие площадь ее взаимодействия с медитором. На мембране нет потенцило-зависимых ионных каналов, зато высока плотность рецептор-управляемых каналов (ионная селективность их низкая). • Число рецепторов на поверхности ПСП мембраны может варьировать. Так, при длительном выделении больших количеств медиатора – происходит десинтезация рецепторов, т. е. снижение их чувствительности к медиатору. В частности, может уменьшаться количество рецепторов на постсинаптической мембране (элиминация рецепторов). • Наоборот, при денервации, когда выделение медиатора резко снижается, количество рецепторов может резко возрастать. • Таким образом, синапс является весьма динамичной структурой, что определяет его пластичность.

Элементы нервно-мышечного синапса

Активные зоны в двигательных нервных окончаниях лягушки и мыши лягушка мышь Синаптический бутон
. Квант медиатора находится в синаптической")
Квантово-везикулярная теория. Освобождение медиатора в синапсе происходит порциями (квантами). Квант медиатора находится в синаптической везикуле и освобождается из нервного окончания посредством экзоцитоза. 1954 Дель Кастилло и Катц детально описывают ПКП и МПКП в нервно-мышечном синапсе. Делается предположение, что медиатор освобождается определенными порциямиквантами. 1955 Пали, Паллад, Де Робертис и Беннетт обнаружили синаптические везикулы с использованием электронного микроскопа.

А В Б Г
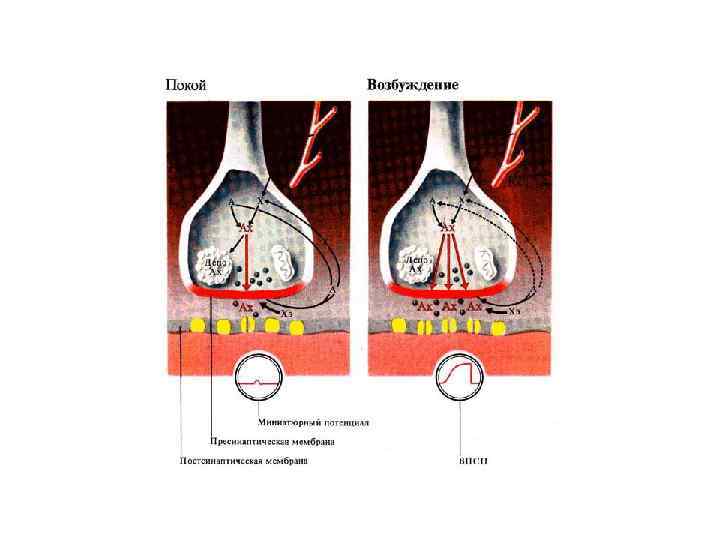
 существует только локально на постсинаптической мембране.")
Потенциал концевой пластинки Возбуждающий потенциал концевой пластинки (ВПСП) существует только локально на постсинаптической мембране. Его величина определяется количеством выделившихся квантов медиатора. 1) Поэтому ВПСП, в отличие от ПД, не подчиняется закону «Все или ничего» . , а подчиняется правилу суммации: Чем больше выделяется медиатора, тем больше величина ВПСП. 2) Второе отличие ВПСП от ПД состоит в электротоническом распространении, т. е. затухании потенциала по мере удаления от концевой пластинки. • Вне возбуждения – на концевой пластинке регистрируются миниатюрные потенциалы концевой пластинки, представляющие собой небольшие волны деполяризации, величиной 0, 5 м. В. Их происхождение связано со спонтанным выделением квантов медиатора из пресинаптичесской мембраны, вследствие самопроизвольного слипания синаптических пузырьков с мембраной (~1 квант в сек). Для возникновения ВПКП необходимо одновременное выделение нескольких сотен квантов медиатора.

Потенциалы и токи концевой пластинки на разных расстояниях от нее

Если синапс возбуждающий, то увеличивается проницаемость постсинаптической мембраны для натрия и калия. Возникает ВПСП. Он существует локально: только на постсинаптической мембране. Но если величина деполяризации достигает критического уровня, то возникает потенциал действия. Если синапс тормозной, то выделяющийся медиатор активирует калиевые каналы и каналы для хлора. Развивающаяся гиперполяризация ( ТПСП ) распространяется на мембрану эфферентной клетки, увеличивает порог возбуждения и снижает возбудимость.

МЕХАНИЗМ ТРАНСФОРМАЦИИ ВПКП В ПД КЛЕТКИ После возникновения ВПКП, между деполяризованной мембраной концевой пластинки и находящемся в покое участком электровозбудимой мембраны мышечного волокна, прилегающего к концевой пластинке – возникает локальный ток. Этот ток обусловлен перераспределением ионов Nа+, вошедших через хемочувствительные каналы - между концевой пластинкой и сарколеммой. Если величина локального тока позволяет деполяризовать мембрану мышечного волокна до Екр. , то открываются потенциалозависимые Са 2+ каналы сарколеммы, которые завершают деполяризацию - возникает ПД, который далее распространяется по мышечному волокну. Таким образом, ВПКП перерастает (или трансформируется) в ПД мышечного волокна.

ТРАНСФОРМАЦИЯ ВПКП В ПД ВПКП -80 м. В Nа+ -30 м. В Са 2+ ПД -60 м. В ПД -30 м. В -60 м. В

Проведение возбуждения в нервных волокнах

Нервно – мышечный синапс Пресинаптическое окончание Постсинаптическая мембрана Электросекреторное сопряжение Выброс ацетилхолина Ацетилхолинэстераза Н – АЦХ рецептора ВПСП ПД сарколеммы Сокращение мышцы Передача возбуждения в нервно – мышечном синапсе

Химические синапсы по эффекту , оказываемому медиатором на постсинаптическую мембрану делят на: 1. Ионотропные 2. Метаботропные

Ионотропный синапс

Передача возбуждения в химическом синапсе 1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся в пресинаптической терминали и концентрирующиеся в активных зонах пресинаптической мембраны. 2. Приходящий по аксону ПД деполяризует пресинаптическую мембрану. 3. Вследствие деполяризации открываются потенциалозависимые Са 2+‑каналы, и Са 2+ поступает в терминаль. 4. Увеличение внутриклеточного [Са 2+] запускает слияние синаптических пузырьков с пресинаптической мембраной и выброс нейромедиатора в синаптическую щель (экзоцитоз).

Передача возбуждения в химическом синапсе 5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть молекул нейромедиатора связывается со специфичными для них рецепторами постсинаптической мембраны. 6. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны либо прямо (поступление ионов через ионотропные рецепторы) либо опосредованно — активация ионных каналов через систему G‑белка (метаботропные рецепторы). 7. Инактивация нейромедиаторов происходит либо путём их ферментной деградации, либо молекулы нейромедиатора захватываются клетками.

Эл.генез. 2-3. Законы тока.ppt