Презентация_видообразование.ppt
- Количество слайдов: 33
 концепция. Вид – это категория мышления, некий класс к которому принадлежат все") Логическая (философская) концепция. Вид – это категория мышления, некий класс к которому принадлежат все объекты, обладающие определенными общими свойствами. Вид – «species» , от глагола «specere» - смотреть, разглядывать, т. е. вид – это то, что можно различить, выделить. Морфологическая концепция вида. Типологическая концепция. Вид – это совокупность особей, сходных по своим морфологическим признакам. Вид – это неделимая категория и неизменная во времени. В систематике - «голотип» и «паратипы» . На данной концепции построены все определеители.
Логическая (философская) концепция. Вид – это категория мышления, некий класс к которому принадлежат все объекты, обладающие определенными общими свойствами. Вид – «species» , от глагола «specere» - смотреть, разглядывать, т. е. вид – это то, что можно различить, выделить. Морфологическая концепция вида. Типологическая концепция. Вид – это совокупность особей, сходных по своим морфологическим признакам. Вид – это неделимая категория и неизменная во времени. В систематике - «голотип» и «паратипы» . На данной концепции построены все определеители.
 Политипическая концепция вида Вид – это система подвидов, каждый из которых занимает определенную область в переделах видового ареала. 21 форма длиннохвостой синицы, подвиды: 1 – Aegithalos caudatus alpinus; 2 – Aeg. c. trphronotus; 3 – Aeg. c. irbii; 4 – Aeg. c. rosaceus; 5– Aeg. c. europaeus; 6 – Aeg. c. caudatus; 7 – Aeg. c. japonicus; 8 – Aeg. c. passakei; 9 – Aeg. c. major; 10 – Aeg. c. tauricus; 11 – Aeg. c. italiae; 12 – Aeg. c. glaucogularis; 13 – Aeg. c. trivirgatus; 14 – Aeg. c. magnus; 15 – Aeg. c. macedonicus. Непрервывность изменчивости признаков или признака нарушена хиатусом (разрывом). Типологические виды разделены хиатусами. Где граница между видом и подвидом?
Политипическая концепция вида Вид – это система подвидов, каждый из которых занимает определенную область в переделах видового ареала. 21 форма длиннохвостой синицы, подвиды: 1 – Aegithalos caudatus alpinus; 2 – Aeg. c. trphronotus; 3 – Aeg. c. irbii; 4 – Aeg. c. rosaceus; 5– Aeg. c. europaeus; 6 – Aeg. c. caudatus; 7 – Aeg. c. japonicus; 8 – Aeg. c. passakei; 9 – Aeg. c. major; 10 – Aeg. c. tauricus; 11 – Aeg. c. italiae; 12 – Aeg. c. glaucogularis; 13 – Aeg. c. trivirgatus; 14 – Aeg. c. magnus; 15 – Aeg. c. macedonicus. Непрервывность изменчивости признаков или признака нарушена хиатусом (разрывом). Типологические виды разделены хиатусами. Где граница между видом и подвидом?
 Биологическая концепция вида Вид - это совокупность популяций, особи которых в природных условиях могут скрещиваться между собой, но репродуктивно изолированных от популяций других видов. Вид - это совокупность организмов явно обособленная и четко ограниченная от других таких же совокупностей, хотя бы по одному из следующих критериев: ØМорфологическому, ØЭтолого-экологическому, ØФизиолого-биохимическому, ØРепродуктивному.
Биологическая концепция вида Вид - это совокупность популяций, особи которых в природных условиях могут скрещиваться между собой, но репродуктивно изолированных от популяций других видов. Вид - это совокупность организмов явно обособленная и четко ограниченная от других таких же совокупностей, хотя бы по одному из следующих критериев: ØМорфологическому, ØЭтолого-экологическому, ØФизиолого-биохимическому, ØРепродуктивному.
 Высшие таксономические категории Это абстрактные логические категории. Реально в природе существует только виды. В основе таксономии – иерархическая система категорий, в которой группы на каждом уровне образуются путем объединения групп, стоящий на одну ступень ниже. .
Высшие таксономические категории Это абстрактные логические категории. Реально в природе существует только виды. В основе таксономии – иерархическая система категорий, в которой группы на каждом уровне образуются путем объединения групп, стоящий на одну ступень ниже. .

 Репродуктивные изолирующие механизмы. В целом изолирующие механизмы подразделяются на два основных типа. 1. Презиготические механизмы, т. е. предшествующие образованию зиготы, создающие препятствия для спаривания особей, относящихся к разным популяциям. 2. Постзиготические механизмы, действующие после образования зиготы, приводящие к снижению жизнеспособности или плодовитости гибридного потомства.
Репродуктивные изолирующие механизмы. В целом изолирующие механизмы подразделяются на два основных типа. 1. Презиготические механизмы, т. е. предшествующие образованию зиготы, создающие препятствия для спаривания особей, относящихся к разным популяциям. 2. Постзиготические механизмы, действующие после образования зиготы, приводящие к снижению жизнеспособности или плодовитости гибридного потомства.
 Презиготические изолирующие механизмы подразделяют на группы в зависимости от факторов, создающих препятствие для спаривания особей: 1. Экологическая изоляция обеспечивается экологическими факторами, когда популяции занимают одну и ту же территорию, но различные местообитания и поэтому не встречаются друг с другом. 2. Временная изоляция — разным временем размножения, если спаривание у животных или цветение у растений происходит в разное время года или в разное время суток. 3. Этологическая, или поведенческая, изоляция — разным поведением в период размножения, что приводит к отсутствию взаимной привлекательности самца и самки. 4. Механическая изоляция обусловлена различиями в размерах или форме половых органов или же в строении цветков 5. Гаметическая изоляция: гаметы самки и самца не взаимодействуют друг с другом или сперматозойды утрачивают жизнеспособность в половых путях самки или на рыльце пестика
Презиготические изолирующие механизмы подразделяют на группы в зависимости от факторов, создающих препятствие для спаривания особей: 1. Экологическая изоляция обеспечивается экологическими факторами, когда популяции занимают одну и ту же территорию, но различные местообитания и поэтому не встречаются друг с другом. 2. Временная изоляция — разным временем размножения, если спаривание у животных или цветение у растений происходит в разное время года или в разное время суток. 3. Этологическая, или поведенческая, изоляция — разным поведением в период размножения, что приводит к отсутствию взаимной привлекательности самца и самки. 4. Механическая изоляция обусловлена различиями в размерах или форме половых органов или же в строении цветков 5. Гаметическая изоляция: гаметы самки и самца не взаимодействуют друг с другом или сперматозойды утрачивают жизнеспособность в половых путях самки или на рыльце пестика
 На Гавайских островах обитает два вида фруктовых мушек, которые внешне очень похожи. Оба вида обитают в одних и тех же местах, питаясь соком одного и того же древесного растения. Однако их экологическое положение различно. §Первый вид проводит жизнь в кроне деревьев, питаясь соком, стекающим по стволам и ветвям верхних ярусов. §Второй — в лесной подстилке, отыскивая лужицы накапавшего с дерева сока. Скрещивания между этими видами никогда не происходит из-за пространственной разобщенности, которая возникает вследствие разной экологической специализации.
На Гавайских островах обитает два вида фруктовых мушек, которые внешне очень похожи. Оба вида обитают в одних и тех же местах, питаясь соком одного и того же древесного растения. Однако их экологическое положение различно. §Первый вид проводит жизнь в кроне деревьев, питаясь соком, стекающим по стволам и ветвям верхних ярусов. §Второй — в лесной подстилке, отыскивая лужицы накапавшего с дерева сока. Скрещивания между этими видами никогда не происходит из-за пространственной разобщенности, которая возникает вследствие разной экологической специализации.
 Интересный пример поведенческой изоляции демонстрируют некоторые виды светлячков. Для каждого из совместно обитающих видов характерна своя световая траектория и свои типы испускаемых световых сигналов. Траектории могут быть зигзагообразными, прямыми или в форме петли, а световые пульсации короткими или длинными в виде устойчивых отблесков. При спаривании особи подбирают друга, строго ориентируясь на тип светового сигнала.
Интересный пример поведенческой изоляции демонстрируют некоторые виды светлячков. Для каждого из совместно обитающих видов характерна своя световая траектория и свои типы испускаемых световых сигналов. Траектории могут быть зигзагообразными, прямыми или в форме петли, а световые пульсации короткими или длинными в виде устойчивых отблесков. При спаривании особи подбирают друга, строго ориентируясь на тип светового сигнала.
 Различные виды реагируют на одни и те же факторы по-разному, что является причиной несовпадения сроков размножения. На рисунке показаны различия в сроках размножения у разных видов амфибий, живущих в одних и тех же районах. У животных с наружным оплодотворением (морских звезд и некоторых видов моллюсков) роль изолирующих факторов играют видоспецифичные различия в строении рецепторов, которые обеспечивают слияние половых клеток. У животных с внутренним оплодотворением эту роль выполняют различия в строении половых органов.
Различные виды реагируют на одни и те же факторы по-разному, что является причиной несовпадения сроков размножения. На рисунке показаны различия в сроках размножения у разных видов амфибий, живущих в одних и тех же районах. У животных с наружным оплодотворением (морских звезд и некоторых видов моллюсков) роль изолирующих факторов играют видоспецифичные различия в строении рецепторов, которые обеспечивают слияние половых клеток. У животных с внутренним оплодотворением эту роль выполняют различия в строении половых органов.
 Постзиготические изолирующие механизмы также подразделяются на группы в зависимости от того: §приводят ли они к нарушениям развития самих гибридов и в конечном счете к их нежизнеспособности §к неспособности гибридов производить полноценные гаметы. Межвидовые гибриды обычно быстро погибают или остаются бесплодными. Мул — гибрид лошади и осла, лошак — гибрид жеребца и ослицы стерильны, не могут произвести потомство из-за того, что его набор хромосом несбалансирован (не проходит мейоз). Бесплодны гибриды • зайца-беляка и зайца-русака, куницы и соболя. Обычно репродуктивная изоляция между видами поддерживается несколькими механизмами. Временная изоляция чаще встречается у растений, а этологическая — у животных
Постзиготические изолирующие механизмы также подразделяются на группы в зависимости от того: §приводят ли они к нарушениям развития самих гибридов и в конечном счете к их нежизнеспособности §к неспособности гибридов производить полноценные гаметы. Межвидовые гибриды обычно быстро погибают или остаются бесплодными. Мул — гибрид лошади и осла, лошак — гибрид жеребца и ослицы стерильны, не могут произвести потомство из-за того, что его набор хромосом несбалансирован (не проходит мейоз). Бесплодны гибриды • зайца-беляка и зайца-русака, куницы и соболя. Обычно репродуктивная изоляция между видами поддерживается несколькими механизмами. Временная изоляция чаще встречается у растений, а этологическая — у животных
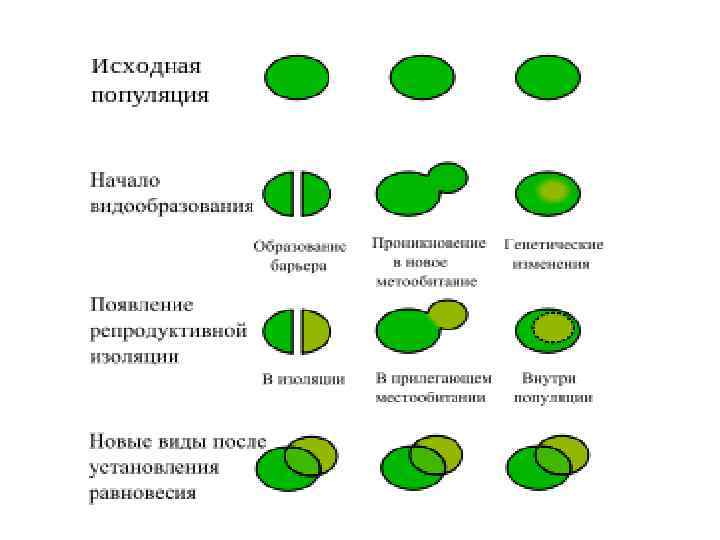
 Наступление моря может привести к изоляции участков суши и популяций, обитающих на них, что является одной из причин образования новых видов
Наступление моря может привести к изоляции участков суши и популяций, обитающих на них, что является одной из причин образования новых видов
") Основные способы видообразования: I—филетическое, II— дивергентное (истинное)
Основные способы видообразования: I—филетическое, II— дивергентное (истинное)
 МЕХАНИЗМЫ 1. Микроэволюционные процессы. 2. Интрогрессия - приобретение генов другого вида при межвидовой гибридизации (естественная и искуственная). Механизм: • образование гибридов первого поколения • возвратное скрещивание с одним или обоими родительскими видами. Если возвратное скрещивание происходит многократно а ряде последовательных поколений, то возникает поток генов из генетической системы одного вида в генетическую систему другого вида. Накапливаемые генетические изменения проводят к филетическому видообразованию. Наиболее распространена у растений.
МЕХАНИЗМЫ 1. Микроэволюционные процессы. 2. Интрогрессия - приобретение генов другого вида при межвидовой гибридизации (естественная и искуственная). Механизм: • образование гибридов первого поколения • возвратное скрещивание с одним или обоими родительскими видами. Если возвратное скрещивание происходит многократно а ряде последовательных поколений, то возникает поток генов из генетической системы одного вида в генетическую систему другого вида. Накапливаемые генетические изменения проводят к филетическому видообразованию. Наиболее распространена у растений.
 Примеры филетического видообразования в ряду ископаемых европейских слонов по ламеллярному индексу (количеству эмали на зубах).
Примеры филетического видообразования в ряду ископаемых европейских слонов по ламеллярному индексу (количеству эмали на зубах).
 Интересный пример усиления гибридизации между родственными видами описан в 1973 г. Л. С. Рябовым: в Воронежской области за последние 13 лет отмечено свыше 100 особей-гибридов волка и домашней собаки, причем наблюдается тенденция к увеличению числа этих гибридов и расширению зоны гибридизации. Известно, что обычные отношения волка и собаки в целом антагонистичны - волки нередко нападают на собак, используя их как добычу. Изменения взаимоотношений этих двух видов, по мнению Л. С. Рябова, связаны с сокращением численности волков и нарушением нормального соотношения числа самцов и самок в их популяциях. Интрогрессивная гибридизация может способствовать усилению наследственной изменчивости родственных видов, обогащая их генофонды новыми аллелями.
Интересный пример усиления гибридизации между родственными видами описан в 1973 г. Л. С. Рябовым: в Воронежской области за последние 13 лет отмечено свыше 100 особей-гибридов волка и домашней собаки, причем наблюдается тенденция к увеличению числа этих гибридов и расширению зоны гибридизации. Известно, что обычные отношения волка и собаки в целом антагонистичны - волки нередко нападают на собак, используя их как добычу. Изменения взаимоотношений этих двух видов, по мнению Л. С. Рябова, связаны с сокращением численности волков и нарушением нормального соотношения числа самцов и самок в их популяциях. Интрогрессивная гибридизация может способствовать усилению наследственной изменчивости родственных видов, обогащая их генофонды новыми аллелями.
 БЫСТРОЕ ВИДООБРАЗОВАНИЕ Генетические механизмы: • Фиксация хромосомных перестроек; • Полиплоидия: üаутополиплоидия, üаллополиплоидия – сетчатое видообразование.
БЫСТРОЕ ВИДООБРАЗОВАНИЕ Генетические механизмы: • Фиксация хромосомных перестроек; • Полиплоидия: üаутополиплоидия, üаллополиплоидия – сетчатое видообразование.
 Опыты Г. Д. Карпетченко Амфидиплоид — плодовитый гибридный организм, сочетающий полные диплоидные наборы хромосом обоих родительских видов. Рафанобрассика ((Raphano-Brassica )- амфидиплоид между капустой и редькой.
Опыты Г. Д. Карпетченко Амфидиплоид — плодовитый гибридный организм, сочетающий полные диплоидные наборы хромосом обоих родительских видов. Рафанобрассика ((Raphano-Brassica )- амфидиплоид между капустой и редькой.

 В настоящее время считается, что зеленые лягушки в Восточной Европе представлены тремя таксонами : • лягушка озерная (R. ridibunda Pallas, 1771) • прудовая (R. lessonae Camerano, 1882) это «хорошие» , «менделевские» виды, • съедобная лягушка (R. kl. esculenta Linneus, 1758) – результат гибридизации указанных выше родительских видов; она имеет большой современный ареал – от Франции до Поволжья, – и большинством специалистов признается в качестве самостоятельного вида. Для данного комплекса характерно необычное видообразование, механизмы которого включают : • гибридизацию, • полуклональное (мероклональное) наследование, • полиплоидию • разнообразие состава популяционных систем, где R. esculenta может быть представлена как одним, так и двумя полами
В настоящее время считается, что зеленые лягушки в Восточной Европе представлены тремя таксонами : • лягушка озерная (R. ridibunda Pallas, 1771) • прудовая (R. lessonae Camerano, 1882) это «хорошие» , «менделевские» виды, • съедобная лягушка (R. kl. esculenta Linneus, 1758) – результат гибридизации указанных выше родительских видов; она имеет большой современный ареал – от Франции до Поволжья, – и большинством специалистов признается в качестве самостоятельного вида. Для данного комплекса характерно необычное видообразование, механизмы которого включают : • гибридизацию, • полуклональное (мероклональное) наследование, • полиплоидию • разнообразие состава популяционных систем, где R. esculenta может быть представлена как одним, так и двумя полами
 Симпатрическое видообразование связано с зарождением нового вида в рамках исходной популяции. Такое зарождение может происходить в результате экологической (например, пищевой) специализации, поэтому данную форму видообразования часто называют экологической Полагают, что пять видов синиц образовались в связи с пищевой специализацией: по выбору мест кормежки, по составу поедаемых кормов, по способам их поиска и добычи. Cиница большая долбит ветви и древесные стволы; мелкие виды (лазоревка) — только стебли травянистых растений. Самые мелкие виды (московки, хохлатые синицы) чаще обследуют в поисках корма концевые ветви деревьев. Синица большая питается крупными насекомыми; лазоревка, гаичка и московка добывают мелких насекомых в щелях коры и в почках; хохлатая синица питается семенами хвойных деревьев.
Симпатрическое видообразование связано с зарождением нового вида в рамках исходной популяции. Такое зарождение может происходить в результате экологической (например, пищевой) специализации, поэтому данную форму видообразования часто называют экологической Полагают, что пять видов синиц образовались в связи с пищевой специализацией: по выбору мест кормежки, по составу поедаемых кормов, по способам их поиска и добычи. Cиница большая долбит ветви и древесные стволы; мелкие виды (лазоревка) — только стебли травянистых растений. Самые мелкие виды (московки, хохлатые синицы) чаще обследуют в поисках корма концевые ветви деревьев. Синица большая питается крупными насекомыми; лазоревка, гаичка и московка добывают мелких насекомых в щелях коры и в почках; хохлатая синица питается семенами хвойных деревьев.
. Симпатрическое видообразование •") . Цихлиды из африканского озера Ньяса - золотые меланохромисы (Melanochromis auratus). Симпатрическое видообразование • В африканском оз. Виктория обитает около 500 эндемичных видов рыб (из семейства цихлидовых) с различающимися экологическими нишами. Есть питающиеся планктоном; есть настоящие хищники, нападающие на других рыб; а есть специализирующиеся на питании моллюсками и даже икрой других видов. • Судя по молекулярно-генетическим данным, эти виды цихлид — близкие родственники, а образование всего веера наблюдаемых форм произошло совсем недавно, поскольку 15 тыс. лет назад оз. Виктория, по свидетельствам геологов, полностью высыхало. (Правда, в последнее время появились сведения о том, что начало дивергенции видов уходит в более далекое прошлое — до 200 тыс. лет назад. ) • В озерах Виктория и Малави (Ньяса) число видов цихлид необычайно велико, а в реках озерных бассейнов очень мало.
. Цихлиды из африканского озера Ньяса - золотые меланохромисы (Melanochromis auratus). Симпатрическое видообразование • В африканском оз. Виктория обитает около 500 эндемичных видов рыб (из семейства цихлидовых) с различающимися экологическими нишами. Есть питающиеся планктоном; есть настоящие хищники, нападающие на других рыб; а есть специализирующиеся на питании моллюсками и даже икрой других видов. • Судя по молекулярно-генетическим данным, эти виды цихлид — близкие родственники, а образование всего веера наблюдаемых форм произошло совсем недавно, поскольку 15 тыс. лет назад оз. Виктория, по свидетельствам геологов, полностью высыхало. (Правда, в последнее время появились сведения о том, что начало дивергенции видов уходит в более далекое прошлое — до 200 тыс. лет назад. ) • В озерах Виктория и Малави (Ньяса) число видов цихлид необычайно велико, а в реках озерных бассейнов очень мало.
 Симпатрическое видообразование форель Озеро Севан
Симпатрическое видообразование форель Озеро Севан
 Симпатрическое видообразование Alectorolo phus major - погремок весенний Погремок осенний На нескашиваемых лугах большой погремок (Alectorolophus major) цветет на протяжении всего лета. Регулярное скашивание травы в середине лета привело к образованию двух экологических рас этого растения, различающиеся по срокам цветения. За несколько десятилетий независимой эволюции у них сформировались и морфологические различия: у весенней расы цветы желтые, у осенней - оранжевые. Помимо этих двух форм возникла еще одна, засоряющая посевы ржи. Созревание семян у этой формы погремка приурочено к срокам жатвы, коробочки перестали открываться, а у семян редуцировались летучки, в результате чего они не отсеиваются от зерен ржи.
Симпатрическое видообразование Alectorolo phus major - погремок весенний Погремок осенний На нескашиваемых лугах большой погремок (Alectorolophus major) цветет на протяжении всего лета. Регулярное скашивание травы в середине лета привело к образованию двух экологических рас этого растения, различающиеся по срокам цветения. За несколько десятилетий независимой эволюции у них сформировались и морфологические различия: у весенней расы цветы желтые, у осенней - оранжевые. Помимо этих двух форм возникла еще одна, засоряющая посевы ржи. Созревание семян у этой формы погремка приурочено к срокам жатвы, коробочки перестали открываться, а у семян редуцировались летучки, в результате чего они не отсеиваются от зерен ржи.
 Одна из форм — однолетник, растущий в более или менее сухих местах. Цветет ранней весной. Вторая представлена многолетними растениями, которые многочисленны во влажных, тенистых прибрежных полосах. Время цветения сдвинуто на летний сезон, а ранняя весна предназначена для накопления зеленой массы. Эти два экотипа, разобщены по времени цветения, и это препятствует их скрещиванию, то есть имеет место предзиготическая изоляция Два экотипа губастика крапчатого — обитатель влажных прибрежий (слева) и более сухих ландшафтов (справа). Разделение на эти экотипы произошло в том числе благодаря инверсии генов, отвечающих за сроки цветения. Американские ученые экспериментально доказали, что хромосомные инверсии способствуют симпатрическому видообразованию. Они подавляют рекомбинацию набора генов с полезными мутациями, потому варианты с инверсией получают преимущество и быстро распространяются в подходящих местообитаниях. Материалом для доказательства послужили популяции губастика крапчатого, которые, получив инверсию, захватившую гены сроков цветения, разделились сообразно местным климатическим условиям.
Одна из форм — однолетник, растущий в более или менее сухих местах. Цветет ранней весной. Вторая представлена многолетними растениями, которые многочисленны во влажных, тенистых прибрежных полосах. Время цветения сдвинуто на летний сезон, а ранняя весна предназначена для накопления зеленой массы. Эти два экотипа, разобщены по времени цветения, и это препятствует их скрещиванию, то есть имеет место предзиготическая изоляция Два экотипа губастика крапчатого — обитатель влажных прибрежий (слева) и более сухих ландшафтов (справа). Разделение на эти экотипы произошло в том числе благодаря инверсии генов, отвечающих за сроки цветения. Американские ученые экспериментально доказали, что хромосомные инверсии способствуют симпатрическому видообразованию. Они подавляют рекомбинацию набора генов с полезными мутациями, потому варианты с инверсией получают преимущество и быстро распространяются в подходящих местообитаниях. Материалом для доказательства послужили популяции губастика крапчатого, которые, получив инверсию, захватившую гены сроков цветения, разделились сообразно местным климатическим условиям.
 О меньшей приспособленности межвидовых гибридов Agapornis rоseicollis – розовощёкий неразлучник Симпатрические виды …поведение, связанное с постройкой гнезда, у разных видов неразлучников. Особи одного вида, Agapornis personata, отрывают кусочки материала для гнезда и переносят их к гнезду в клюве. Представители другого вида, А. rоseicollis, не держат их в клюве, а засовывают под боковые перья. Исследованные Дилгером гибриды F 1 обнаружили смешанный тип поведения, при котором птицы пытались засунуть материал для гнезда в перья, затем вынимали его, брали их в клюв, а потом все начиналось сначала. Таким образом, их поведение представляло собой малоэффективную смесь из двух типов целесообразного поведения родительских форм.
О меньшей приспособленности межвидовых гибридов Agapornis rоseicollis – розовощёкий неразлучник Симпатрические виды …поведение, связанное с постройкой гнезда, у разных видов неразлучников. Особи одного вида, Agapornis personata, отрывают кусочки материала для гнезда и переносят их к гнезду в клюве. Представители другого вида, А. rоseicollis, не держат их в клюве, а засовывают под боковые перья. Исследованные Дилгером гибриды F 1 обнаружили смешанный тип поведения, при котором птицы пытались засунуть материал для гнезда в перья, затем вынимали его, брали их в клюв, а потом все начиналось сначала. Таким образом, их поведение представляло собой малоэффективную смесь из двух типов целесообразного поведения родительских форм.
 Парапатрическое видообразование
Парапатрическое видообразование
 Аллопатрическое видообразование
Аллопатрическое видообразование
 Аллопатрическое видообразование Грандиозные четвертичные колебания климата привели к фрагментации ареалов из-за наступления ледников, затем вторичное расширение ареалов в межледниковья - приводят к вспышкам видообразования.
Аллопатрическое видообразование Грандиозные четвертичные колебания климата привели к фрагментации ареалов из-за наступления ледников, затем вторичное расширение ареалов в межледниковья - приводят к вспышкам видообразования.
 Ареал комплекса больших белоголовых чаек охватывает широким кольцом северное полушарие земного шара. В пределах комплекса выделяют до 15 различных форм, различающихся по окраске спины и крыльев, цветом ног и окологлазничного кольца. Их таксономический статус невозможно точно установить, так как обитающие совместно и негибридизирующие виды (серебристая чайка, чайка-клуша, хохотунья) оказываются связанными друг с другом цепью гибридизирующих подвидов.
Ареал комплекса больших белоголовых чаек охватывает широким кольцом северное полушарие земного шара. В пределах комплекса выделяют до 15 различных форм, различающихся по окраске спины и крыльев, цветом ног и окологлазничного кольца. Их таксономический статус невозможно точно установить, так как обитающие совместно и негибридизирующие виды (серебристая чайка, чайка-клуша, хохотунья) оказываются связанными друг с другом цепью гибридизирующих подвидов.
 серебристая чайка клуша хохотунья
серебристая чайка клуша хохотунья
 Аллопатрическое видообразование Parus maior minor Кольцевой ареал большой синицы Parus maior: I – P. maior; II – P. maior minor III – P. maior cinereus, IV – P. maior boxha renis, V – переходные популяции (maior cinereus), VI - переходные популяции (cinereus minor), VII – перекрывание ареалов maior и minor Встретившиеся в Приамурье подвиды большой синицы не скрещиваются
Аллопатрическое видообразование Parus maior minor Кольцевой ареал большой синицы Parus maior: I – P. maior; II – P. maior minor III – P. maior cinereus, IV – P. maior boxha renis, V – переходные популяции (maior cinereus), VI - переходные популяции (cinereus minor), VII – перекрывание ареалов maior и minor Встретившиеся в Приамурье подвиды большой синицы не скрещиваются