липиды.ppt
- Количество слайдов: 86



ЛИПИДНЫЙ ОБМЕН «Липиды" объединяет вещества, обладающие общим физическим свойством - гидрофобностью, т. е. нерастворимостью в воде. По структуре липиды настолько разнообразны, что у них отсутствует общий признак химического строения. Липиды разделяют на классы, в которые объединяют молекулы, имеющие сходное химическое строение и общие биологические свойства. Основную массу липидов в организме составляют жиры - триацилглицеролы, служащие формой депонирования энергии. Жиры располагаются преимущественно в подкожной жировой ткани и выполняют также функции теплоизоляционной и механической защиты.

ОСНОВНЫЕ ФУНКЦИИ ЛИПИДОВ n Липиды обнаружены в составе всех органов и тканей. Наибольшее количество (90%) содержится в жировой ткани. В мозге липиды составляют половину массу органа; n Липиды в комплексе с белками составляют основу мембран клеток, благодаря чему они участвуют в регуляции деятельности гормонов, ферментов, процессах биологического окисления, транспорта различных веществ и других функциях; Липиды (жиры) являются одним из основных источников энергии: 40 -50% энергопродукции организма обеспечивается триглицеридами, на долю которых приходится более 95% всех липидов. В отличие от углеводов жиры составляют энергетический резерв организма. Преимущество жира в качестве энергетического резерва заключается в том, что жиры являются более восстановленными веществами по сравнению с углеводами (в молекулах углеводов при каждом углеродном атоме есть кислород - группы "-CHOH-"; у жира имеются длинные углеводородные радикалы, в которых преобладают группы "-CH 2 -" - в них нет кислорода). От жира можно отнять больше водорода, который затем проходит по цепи митохондриального окисления с образованием АТФ. n Калорийность углеводов и белков: ~ 4 ккал/грамм. Калорийность жира: ~ 9 ккал/грамм.

В среднем, у человека запас чистых триглицеридов составляет примерно 13 кг. Этих запасов могло бы хватить на 40 дней голодания в условиях умеренной физической нагрузки. Для сравнения: общие запасы гликогена в организме примерно 400 г; при голодании этого количества не хватает даже на одни сутки; n Липиды участвуют в терморегуляции, предохраняют кожу от высыхания, защищают органы от сотрясений; n Липиды, откладываясь в депо служат запасной формой питательных веществ; n Липиды являются источником ненасыщенных высших жирных кислот — витамина Е, одного из незаменимых факторов питания.

КЛАССИФИКАЦИЯ ЛИПИДОВ ЛИПИДЫ Простые Триглицериды Воска Стерины Сложные Фосфолипиды Гликолипиды Липопротеиды

Воска. Представляют собой сложные эфиры высших одноатомных длинноцепочечных спиртов и высокомолекулярных жирных кислот. Это твердые вещества. Они выполняют в основном защитные функции: ланолин у человека предохраняет волосы и кожу от воздействия воды, воск защищает листья и плоды от проникновения воды и микробов и т. д. n Стерины. Это группа высокомолекулярных циклических спиртов, образующих с жирными кислотами сложные эфиры — стериды. Представителем стеринов является холестерин, впервые выделенный из желчных камней Э. Конради еще в ХУII веке.

Холестерин является кристаллическим нерастворимым в воде веществом, способным растворяться в органических растворителях. В организме холестерин выполняет важные функции: является предшественником многих биологически важных соединений (стероидных гормонов, желчных кислот, витамина Е)), входит в состав клеточных мембран, повышает устойчивость эритроцитов к гемолизу, служит своеобразным изолятором для нервных клеток, обеспечивая проведение нервных импульсов. В крови содержатся как свободный, так и эфиросвязанный холестерин, концентрация которого примерно в З раза выше свободного. Содержание общего холестерина в крови составляет 3. 09 -6. 11 ммоль/л (115— 340 мг%), из которых свободного — О, 74— 2, 2 ммоль/л (29— 85 мг%).
 (триглицериды, нейтральные жиры). Являются сложными эфирами глицерина и трех молекул высших")
ТРИГЛИЦЕРИДЫ Триацилглицерины (ТАГ) (триглицериды, нейтральные жиры). Являются сложными эфирами глицерина и трех молекул высших жирных кислот. Они имеют следующую структуру: Н 2 С – О – С = О R 0 НС – О – С = О R 1 Н 2 С – О – С = О R 2 где R 0, R 1, R 2 — остатки насыщенных и ненасыщенных высших жирных кислот.

Н 2 С – О – С = О C 15 H 31 трипальметин (простой жир) Н 2 С – О – С = О C 15 H 31 НС – О – С = О C 16 H 33 Н 2 С – О – С = О C 18 H 36 Пальмитодистерин (смешанный жир)

Фосфолипиды - большой класс липидов, получивший своё название из-за остатка фосфорной кислоты, придающего им свойства амфифильности. Благодаря этому свойству фосфолипиды формируют бислойную структуру мембран, в которую погружены белки. Клетки или отделы клеток, окружённые мембранами, отличаются по составу и набору молекул от окружающей среды, поэтому химические процессы в клетке разделены и ориентированы в пространстве, что необходимо для регуляции метаболизма. Стероиды, представленные в животном мире холестеролом и его производными, выполняют разнообразные функции. Холестерол - важный компонент мембран и регулятор свойств гидрофобного слоя. Производные холестерола (жёлчные кислоты) необходимы для переваривания жиров. Стероидные гормоны, синтезируемые из холестерола, участвуют в регуляции энергетического, водно-солевого обменов, половых функций. Кроме стероидных гормонов, многие производные липидов выполняют регуляторные функции и действуют, как и гормоны, в очень низких концентрациях.

Например, тромбоцитактивирующий фактор - фосфолипид особой структуры - оказывает сильное влияние на агрегацию тромбоцитов в концентрации 10 -12 М; эйкозаноиды, производные полиеновых жирных кислот, вырабатываемые почти всеми типами клеток, вызывают разнообразные биологические эффекты в концентрациях не более 10 -19 М. Из приведённых примеров следует, что липиды обладают широким спектром биологических функций. В тканях человека количество разных классов липидов существенно различается. В жировой ткани жиры составляют до 75% сухого веса. В нервной ткани липидов содержится до 50% сухого веса, основные из них фосфолипиды и сфингомиелины (30%), холестерол (10%), ганглиозиды и цереброзиды (7%). В печени общее количество липидов в норме не превышает 10 -13%. Липиды разных классов существенно отличаются по структуре и функциям. Большинство липидов имеют в своём составе жирные кислоты, связанные сложноэфирной связью с глицеролом, холестеролом или амидной связью с аминоспиртом сфингозином.

Структура, состав и свойства жирных кислот и триглицеридов Жирные кислоты - структурные компоненты различных липидов. В составе триацилглицеролов жирные кислоты выполняют функцию депонирования энергии, так как их радикалы содержат богатые энергией СН 2 -группы. При окислении СН-связей энергии выделяется больше, чем при окислении углеводов, в которых атомы углерода уже частично окислены (-НСОН-). В составе фосфолипидов и сфинголипидов жирные кислоты образуют внутренний гидрофобный слой мембран, определяя его свойства. Жиры и фосфолипиды организма при нормальной температуре тела имеют жидкую консистенцию, так количество ненасыщенных жирных кислот преобладает над насыщенными. В фосфолипидах мембран ненасыщенных кислот может быть до 80 -85%, а в составе жиров подкожного жира - до 60%. В свободном, неэтерифицированном состоянии жирные кислоты в организме содержатся в небольшом количестве, например в крови, где они транспортируются в комплексе с белком альбумином.

Жирные кислоты липидов человека представляют собой углеводородную неразветвлённую цепь, на одном конце которой находится карбоксильная группа, а на другом - метильная группа (ω-углеродный атом). Большинство жирных кислот в организме содержат чётное число атомов углерода - от 16 до 20 (табл. 8 -1 и 8 -2). Жирные кислоты, не содержащие двойных связей, называют насыщенными. Основной насыщенной жирной кислотой в липидах человека является пальмитиновая (до 30 -35%). Жирные кислоты, содержащие двойные связи, называют ненасыщенными. Ненасыщенные жирные кислоты представлены моноеновыми (с одной двойной связью) и полиеновыми (с двумя и большим числом двойных связей). Если в составе жирной кислоты содержатся две и более двойных связей, то они располагаются через СН 2 группу. Имеется несколько способов изображения структуры жирных кислот.

Двойные связи в жирных кислотах в организме человека имеют цисконфигурацию. Это означает, что ацильные фрагменты находятся по одну сторону двойной связи. Цис-конфигурация двойной связи делает алифатическую цепь жирной кислоты изогнутой, что нарушает упорядоченное расположение насыщенных радикалов жирных кислот в фосфолипидах мембран (рис. 81) и снижает температуру плавления. Чем больше двойных связей в жирных кислотах липидов, тем ниже температура их плавления. Жирные кислоты с транс-конфигурацией двойной связи могут поступать в организм с пищей, например в составе маргарина. В этих кислотах отсутствует излом, характерный для цис-связи, поэтому жиры, содержащие такие ненасыщенные кислоты, имеют более высокую температуру плавления, т. е. более твёрдые по консистенции.

Рис. 8 -1. Конфигурации радикалов жирных кислот. А - излом радикала жирной кислоты при двойной связи в цис-конфигурации; Б - нарушение упорядоченного расположения радикалов насыщенных жирных кислот в гидрофобном слое мембран ненасыщенной кислотой с цисконфигурацией двойной связи

Насыщенные Миристиновая 14: 0 Пальмитиновая 16: 0 Стеариновая 18: 0 Моноеновые Пальмитоолеиновая 16: 1Δ 9 Олеиновая 18: 1Δ 9 Полиеновые Линолевая* 18: 2Δ 9, 12 α-Линоленовая*18: 3Δ, 9, 12, 15 Эйкозатриеновая 20: 3 Δ 8, 11, 14 Арахидоновая**20: 4Δ 5, 8, 11, 146 Эйкозапентаеновая 20: 5Δ 5, 8, 11, 14, 17 Докозопентаеновая 22: 5Δ 7, 10, 13, 16, 19 Докозагексаеновая 22: 6Δ 4, 7, 10, 13, 16, 193
 не")
Большинство жирных кислот синтезируется в организме человека, однако полиеновые кислоты (линолевая и α-линоленовая) не синтезируются и должны поступать с пищей. Эти жирные кислоты называют незаменимыми, или эссенциальными. Основные источники полиеновых жирных кислот для человека - жидкие растительные масла и рыбий жир, в котором содержится много кислот семейства ω-3.

Классификация ацилглицеролов, фосфолипидов и сфинголипидов Ацилглицеролы триглицериды, диглицериды моноглицериды Фосфолипиды Сфингомиелины*Глицерофосфолипиды: Фосфатидилхолин Фосфатидилсерин Фосфатидилэтаноламин Фосфатидилглицерол Фосфатидилинозитолбисфосфат Фосфатидная кислота Кардиолипин Сфинголипиды Гликолипиды: Цероброзиды Глобозиды Сульфатиды Ганглиозиды

n Ацилглицеролы - сложные эфиры трёхатомного спирта глицерола и жирных кислот. Глицерол может быть связан с одной, двумя или тремя жирными кислотами, соответственно образуя моно-, ди- или триацилглицеролы (МАГ, ДАГ, ТАГ). Основную массу липидов в организме человека составляют триацилглицеролы - жиры. У человека с массой тела 70 кг в норме содержится до 10 кг жиров. Они запасаются в жировых клетках - адипоцитах и используются при голодании как источники энергии. Жиры, содержащие преимущественно насыщенные кислоты, являются твёрдыми (говяжий, бараний жиры), а содержащие большое количество ненасыщенных кислот - жидкими. Жидкие жиры или масла обычно имеют растительное происхождение. Из животных пищевых жиров наиболее насыщен бараний жир, который практически не содержит незаменимых кислот. Ценными пищевыми жирами являются рыбий жир и растительные масла, содержащие незаменимые жирные кислоты. В организме рыб полиеновые жирные кислоты ω-3 и ω-6 также не синтезируются, рыбы получают их с пищей (водоросли, планктон).

Фосфолипиды - разнообразная группа липидов, содержащих в своём составе остаток фосфорной кислоты. Фосфолипиды делят на глицерофосфолипиды, основу которых составляет трёхатомный спирт глицерол, и сфингофосфолипиды - производные аминоспирта сфингозина. Фосфолипиды имеют амфифильные свойства, так как содержат алифатические радикалы жирных кислот и различные полярные группы. Благодаря своим свойствам фосфолипиды не только являются основой всех клеточных мембран, но и выполняют другие функции: образуют поверхностный гидрофильный слой липопротеинов крови, выстилают поверхность альвеол, предотвращая слипание стенок во время выдоха. Некоторые фосфолипиды участвуют в передаче гормонального сигнала в клетки. Сфингомиелины являются фосфолипидами, формирующими структуру миелиновых оболочек и других мембранных структур нервных клеток.
")
Глицерофосфолипиды. Структурная основа глицерофосфолипидов - глицерол. Глицерофосфолипиды (ранее используемые названия - фосфоглицериды или фосфоацилглицеролы) представляют собой молекулы, в которых две жирные кислоты связаны сложноэфирной связью с глицеролом в первой и второй позициях; в третьей позиции находится остаток фосфорной кислоты, к которому, в свою очередь, могут быть присоединены различные заместители, чаще всего аминоспирты (рис. 8 -3). Если в третьем положении имеется только фосфорная кислота, то глицерофосфолипид называется фосфатидной кислотой. Её остаток называют "фосфатидил"; он входит в название остальных глицерофосфолипидов, после которого указывают название заместителя атома водорода в фосфорной кислоте, например фосфатидилэтаноламин, фосфатидилхолин и т. д. Фосфатидная кислота в свободном состоянии в организме содержится в небольшом количестве, но является промежуточным продуктом на пути синтеза как триацилглицеролов, так и глицерофосфолипидов. У глицерофосфолипидов, как и у триацилглицеролов, во второй позиции находятся преимущественно полиеновые кислоты; в молекуле фосфатидилхолина, входящего в структуру мембран, это чаще всего арахидоновая кислота. Жирные кислоты фосфолипидов мембран отличаются от других липидов человека преобладанием полиеновых кислот (до 80 -85%), что обеспечивает жидкое состояние гидрофобного слоя, необходимое для функционирования белков, входящих в структуру мембран.

Рис. 8 -3. Основные глицерофосфолипиды в организме человека.

Некоторые типы плазмалогенов вызывают очень сильные биологические эффекты, действуя как медиаторы. Например, Тромбоцитактивирующий фактор (ТАФ) стимулирует агрегацию тромбоцитов. ТАФ отличается от других плазмалогенов отсутствием двойной связи в алкильном радикале и наличием ацетильной группы во втором положении глицерола вместо жирной кислоты. ТАФ выделяется из фагоцитирующих клеток крови в ответ на раздражение и стимулирует агрегацию тромбоцитов, участвуя таким образом в свёртывании крови. Этот фактор обусловливает также развитие некоторых признаков воспаления и аллергических реакций.

Рис. 8 -4. Плазмалогены.

Сфинголипиды. Аминоспирт сфингозин, состоящий из 18 атомов углерода, содержит гидроксильные группы и аминогруппу. Сфингозин образует большую группу липидов, в которых жирная кислота связана с ним через аминогруппу. Продукт взаимодействия сфингозина и жирной кислоты называют "церамид" (рис. 8 -5). В церамидах жирные кислоты связаны необычной (амидной) связью, а гидроксильные группы способны взаимодействовать с другими радикалами. Церамиды отличаются радикалами жирных кислот, входящих в их состав. Обычно это жирные кислоты с большой длиной цепи - от 18 до 26 атомов углерода. Сфингомиелины. В результате присоединения к ОН-группе церамида фосфорной кислоты, связанной с холином, образуется сфингомие-лин (рис. 8 -5). Сфингомиелины - основные компоненты миелина и мембран клеток мозга и нервной ткани. Сфингомиелины, как и глицерофосфолипиды, имеют амфифильные свойства, обусловленные, с одной стороны, радикалом жирной кислоты и алифатической цепью самого сфингозина, а с другой полярной областью фосфорилхолина.
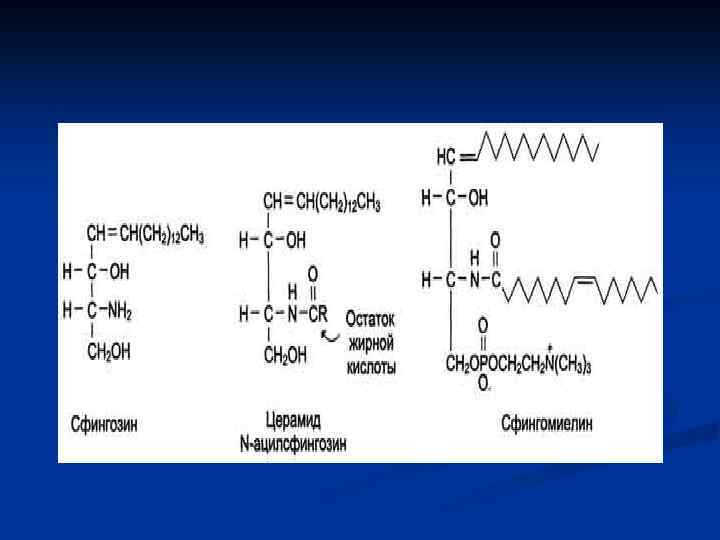

n Гликолипиды. Церамиды - основа большой группы липидов - гликолипидов (см. выше табл. 8 -4). Водород в гидроксильной группе церамида может быть замещён на разные углеводные фрагменты, что определяет принадлежность гликолипида к определённому классу. Гликолипиды находятся в основном в мембранах клеток нервной ткани. Названия "церебро-зиды" и "ганглиозиды" указывают на ткани, откуда они впервые были выделены. n Цереброзиды имеют в своём составе моносахариды. Наиболее распространены цереброзиды, имеющие в своём составе галактозу (галактоцереброзид), реже - глюкозу (глюкоцереброзид). Цереброзиды содержат необычные жирные кислоты, например, галактоцереброзид френозин содержит цереброновую кислоту - 2 гидроксикислоту, содержащую 24 атома углерода (рис. 8 -6).

Рис. 8 -6. Цереброзиды.

Глобозиды, Глобозиды отличаются от цереброзидов тем, что имеют в своём составе несколько углеводных остатков, связанных с церамидом: церамид-глюкоза-галактоза-N-ацетилгалактоза n Цереброзиды и глобозиды относят к нейтральным сфинголипидам, так как они не содержат заряженных групп. n Сульфатиды. Гидроксил у третьего углеродного атома моносахарида, входящего в состав цереброзида, может связывать остаток серной кислоты, т. е. сульфатироваться. В этом случае образуются сульфатиды, обладающие свойствами кислот и поэтому называемые кислыми сфинголипидами (рис. 8 -7). При физиологических значениях р. Н сульфатированный углеводный остаток имеет отрицательный заряд. Около 25% цереброзидов мозга представляют собой сульфатированные производные. Сульфатиды в значительных количествах находят в белом веществе мозга.

Рис. 8 -7. Сульфатиды.

n Ганглиозиды - наиболее сложные по составу липиды. Они содержат несколько углеводных остатков, среди которых присутствует Nацетилнейраминовая кислота. Нейраминовая кислота представляет собой углевод, состоящий из 9 атомов углерода и входящий в группу сиаловых кислот. Ганглиозиды содержатся в основном в ганглиозных клетках нервной ткани, откуда они и получили своё название. Однако ганглиозиды находятся и в плазматических мембранах многих клеток - эритроцитов, гепатоцитов, клеток селезёнки и других органов. Главная роль ганглиозидов определяется их участием в осуществлении межклеточных контактов. Некоторые ганглиозиды служат своеобразными рецепторами для ряда бактериальных токсинов. Строение ганглиозида Gm 2 может быть представлено следующей схемой:

Стероиды - производные восстановленных конденсированных циклических систем - циклопентанпергидрофенантренов. В организме человека основной стероид - холестерол, остальные стероиды - его производные. Растения, грибы и дрожжи не синтезируют холестерол, но образуют разнообразные фитостеролы и микостеролы, не усваиваемые организмом человека. Бактерии не способны синтезировать стероиды. Холестерол входит в состав мембран и влияет на структуру бислоя, увеличивая её жёсткость. Из холестерола синтезируются жёлчные кислоты, стероидные гормоны и витамин D 3. Нарушение обмена холестерола приводит к развитию атеросклероза. В крови 2/3 холестерола находится в этерифицированной форме и 1/3 - в виде свободного холестерола. Эфиры холестерола служат формой его депонирования в некоторых клетках (например, печени, коры надпочечников, половых желёз). Из этих депо холестерол используется для синтеза жёлчных кислот и стероидных гормонов.

Жёлчные кислоты n Жёлчные кислоты обладают поверхностноактивными свойствами и участвуют в переваривании жиров, эмульгируя их и делая доступными для действия панкреатической липазы. Жёлчные кислоты - производные холестерола, в организме человека синтезируются две жёлчные кислоты: холевая и хенодезокеихолевая.

ПЕРЕВАРИВАНИЕ И ВСАСЫВАНИЕ ПИЩЕВЫХ ЛИПИДОВ С пищей в организм ежедневно поступает от 80 до 150 г липидов. Основную массу составляют жиры, наряду с глюкозой служащие главными источниками энергии. Хотя калорийность жиров значительно выше, чем углеводов (9 по сравнению с 4, 7 ккал/моль), при рациональном питании жиры обеспечивают не более 30% от общего количества калорий, поступающих с пищей. Жидкие жиры (масла) содержат в своём составе полиеновые жирные кислоты, которые не синтезируются в организме; поэтому жидкие жиры должны составлять не менее одной трети жиров пищи. С липидами в организм поступают и жирорастворимые витамины A, D, Е, К. Переваривание липидов пищи происходит в кишечнике. Основные продукты гидролиза (жирные кислоты и 2 -моноацилглицеролы) после всасывания подвергаются ресинтезу и последующей упаковке в хиломикроны (ХМ) в клетках слизистой оболочки кишечника.

Эмульгирование жиров Жиры составляют до 90% липидов, поступающих с пищей. Переваривание жиров происходит в тонком кишечнике, однако уже в желудке небольшая часть жиров гидролизуется под действием "липазы языка". Этот фермент синтезируется железами на дорсальной поверхности языка и относительно устойчив при кислых значениях р. Н желудочного сока. Поэтому он действует в течение 1 -2 ч на жиры пищи в желудке. Однако вклад этой липазы в переваривание жиров у взрослых людей незначителен. Основной процесс переваривания происходит в тонкой кишке. Так как жиры - нерастворимые в воде соединения, то они могут подвергаться действию ферментов, растворённых в воде только на границе раздела фаз вода/жир. Поэтому действию панкреатической липазы, гидролизующей жиры, предшествует эмульгирование жиров. Эмульгирование (смешивание жира с водой) происходит в тонком кишечнике под действием солей жёлчных кислот (рис. 8 -11). Жёлчные кислоты синтезируются в печени из холестерола и секретируются в жёлчный пузырь. Содержимое жёлчного пузыря - жёлчь.

Это вязкая жёлто-зелёная жидкость, содержащая главным образом жёлчные кислоты; в небольшом количестве имеются фосфолипиды и холестерол. После приёма жирной пищи жёлчный пузырь сокращается и жёлчь изливается в просвет двенадцатиперстной кишки. Жёлчные кислоты действуют как детергенты, располагаясь на поверхности капель жира и снижая поверхностное натяжение. В результате крупные капли жира распадаются на множество мелких, т. е. происходит эмульгирование жира. Эмульгирование приводит к увеличению площади поверхности раздела фаз жир/вода, что ускоряет гидролиз жира панкреатической липазой. Эмульгированию способствует и перистальтика кишечника.

Рис. 8 -11. Этапы поступления экзогенных жиров в организм.

Гормоны, активирующие переваривание жиров При поступлении пищи в желудок, а затем в кишечник клетки слизистой оболочки тонкого кишечника начинают секретировать в кровь пептидный гормон холецистокинин (панкреозимин). Этот гормон действует на жёлчный пузырь, стимулируя его сокращение, и на экзокринные клетки поджелудочной железы, стимулируя секрецию пищеварительных ферментов, в том числе панкреатической липазы. Другие клетки слизистой оболочки тонкого кишечника в ответ на поступление из желудка кислого содержимого выделяют гормон секретин. Секретин - гормон пептидной природы, стимулирующий секрецию бикарбоната (НСО 3 -) в сок поджелудочной железы.

Панкреатическая липаза выделяется в полость тонкой кишки из поджелудочной железы вместе с белком колипазой. Колипаза попадает в полость кишечника в неактивном виде и частичным протеолизом под действием трипсина превращается в активную форму. Колипаза своим гидрофобным доменом связывается с поверхностью мицеллы эмульгированного жира. Другая часть молекулы способствует формированию такой конформации панкреатической липазы, при которой активный центр фермента максимально приближен к своим субстратам - молекулам жиров, поэтому скорость реакции гидролиза жира резко возрастает. Основными продуктами гидролиза панкреатической липазы являются свободные жирные кислоты и 2 -моноацилглицеролы (βмоноацилглицеролы). Кроме жиров, с пищей поступают фосфолипиды, эфиры холестерола, однако количество этих липидов в составе пищи значительно меньше, чем жиров (≈10%).

Переваривание эфиров холестерола В составе пищи холестерол находится в основном в виде эфиров. Гидролиз эфиров холестерола происходит под действием холестеролэстеразы - фермента, который также синтезируется в поджелудочной железе и секретируется в кишечник. Продукты гидролиза (холестерол и жирные кислоты) всасываются в составе смешанных мицелл.
 крови присутствуют три основных класса")
ТРАНСПОРТ ЖИРОВ ИЗ КИШЕЧНИКА ХИЛОМИКРОНАМИ n В плазме (сыворотке) крови присутствуют три основных класса липидов n Холестерин (ХС) и его эфиры. n Триглицериды (ТГ). n Фосфолипиды (ФЛ). Липиды в водной среде (а значит, и в крови) нерастворимы, поэтому для транспорта липидов кровью в организме образуются комплексы липидов с белками - липопротеины. Все типы липопротеинов имеют сходное строение - гидрофобное ядро и гидрофильный слой на поверхности. Гидрофильный слой образован белками, которые называют апопротеинами, и амфифильными молекулами липидов - фосфолипидами и холестеролом.

Гидрофильные группы этих молекул обращены к водной фазе, а гидрофобные части - к гидрофобному ядру липопротеина, в котором находятся транспортируемые липиды. Некоторые апопротеины интегральные и не могут быть отделены от липопротеина, а другие могут свободно переноситься от одного типа липопротеина к другому. Апопротеины выполняют несколько функций: n формируют структуру липопротеинов; n взаимодействуют с рецепторами на поверхности клеток и таким образом определяют, какими тканями будет захватываться данный тип липопротеинов; n служат ферментами или активаторами ферментов, действующих на липопротеины.

Липопротеиды классифицируются по размеру и плотности. Все липопротеиды имеют сходную структуру. Они состоят из центральной части ( «ядра» ), содержащей нерастворимые в воде липиды (эфиры, холестерин , триглицериды , жирные кислоты) и из оболочки, состоящей из особых белковых молекул (апопротеинов) и растворимых в воде липидов – неэстерифицированного холестерина и фосфолипидов. Молекулы апопротеинов имеют неполярный гидрофобный участок, который связан с липидами, и полярный гидрофильный участок, расположенный на поверхности сферической частицы липопротеинов, и обращенный к окружающей липопротеин водной среде (плазме крови). Гидрофильный участок апопротеина образует водорастворимые связи с молекулами воды. Такая структура липопротеидов определяет их свойство быть частично водорастворимыми, а частично – жирорастворимыми.

Липопротеины плазмы крови.
 путь. Более 95% липидов, поступающих")
Существуют два пути метаболизма липидов и липопротеинов. Экзогенный (пищевой) путь. Более 95% липидов, поступающих с пищей, являются триглицеридами, остальное количество составляют фосфолипиды, свободные жирные кислоты (СЖК), холестерин (в пищевых продуктах присутствует в виде этерифицированного холестерина) и жирорастворимые витамины. Пищевые триглицериды в желудке и двенадцатиперстной кишке под влиянием желудочной и панкреатической липаз превращаются в моноглицериды (МГ) и свободные жирные кислоты. Эфиры холестерина, содержащиеся в пище, подвергаются деэтерификации в свободный холестерин по тому же механизму. Моноглицериды, свободные жирные кислоты и свободный холестерин под действием желчных кислот растворяются и абсорбируются энтероцитами, затем соединяются с триглицеридами и вместе с холестерином включаются в хиломикроны.
 состоят из триглицеридов и являются основной транспортной формой")
Хиломикроны почти полностью (на 80 -95%) состоят из триглицеридов и являются основной транспортной формой экзогенных (пищевых) триглицеридов, перенося их из энтероцитов тонкого кишечника в кровоток. В плазме крови апопротеин C-II на хиломикронах активирует эндотелиальную липопротеинлипазу, под действием которой 90% триглицеридов в хиломикронах расщепляется до глицерина и свободных неэстерифицированных жирных кислот (НЭЖК). НЭЖК используются в жировой и мышечной ткани в качестве энергетического субстрата. Остатки хиломикронов (ремнанты), содержащие холестерин захватываются гепатоцитами и быстро удаляются из кровотока. Этот процесс опосредован аполипопротеином Е (рис. 8 -20).

ХМ имеют довольно большой размер, поэтому после приёма жирной пищи они придают плазме крови опалесцирующий, похожий на молоко, вид. ХМ транспортируют жир к различным тканям, где он утилизируется, поэтому концентрация ХМ в крови постепенно снижается, и плазма опять становится прозрачной. ХМ исчезают из крови в течение нескольких часов. Скорость удаления ХМ из кровотока зависит от: n активности ЛП-липазы; n присутствия ЛПВП, поставляющих апопротеины С-II и Е для ХМ; n активности переноса апо. С-II и апо. Е на ХМ. n Генетические дефекты любого из белков, участвующих в метаболизме ХМ, приводят к развитию семейной гиперхиломикронемии - гиперлипопротеинемии типа I. У таких больных в постабсорбтивном периоде концентрация триацилглицеролов повышена (более 200 мг/дл), плазма крови по виду напоминает молоко и при оставлении на холоде (+4 °С) в ней всплывают белые жирные хлопья, что характерно для гипертриглицеролемии и гиперхиломикронемии.

Рис. 8 -20. Путь экзогенных жиров и хиломикронов. *ЛПЛ - липопротеинлипаза, ЖК - жирные кислоты.

Эндогенный путь В печени из эндогенных триглицеридов и холестерина синтезируются липопротеины очень низкой плотности (ЛПОНП). ЛОНП находятся в кровотоке до тех пор, пока триглицериды, содержащиеся в них, не поступят в периферические ткани. Остатки ЛОНП захватываются гепатоцитами. В результате образуются липопротеины низкой плотности (ЛПНП), которые транспортируют холестерин в периферические ткани. Освобожденный холестерин участвует в синтезе мембран и метаболизме. В то время как в клеточных мембранах происходит обмен веществ, неэстерифицированный холестерин высвобождается в плазму, где связывается с липопротеидами высокой плотности (ЛПВП). Сложные эфиры холестерина ЛПВП превращаются в ЛПОНП и, в итоге, в ЛПНП. Посредством этого цикла ЛПНП доставляет холестерин в клетки, а холестерин возвращается из внепеченочных зон с помощью ЛПВП.
 содержат около 55% триглицеридов, 19% холестерина и 8%")
n Липопротеиды очень низкой плотности (ЛПОНП) содержат около 55% триглицеридов, 19% холестерина и 8% белка (апопротеинов В-100, Е, С-I и C-II). Этот класс липопротеидов синтезируется в печени и является главной транспортной формой эндогенных триглицеридов и холестерина. Благодаря ЛПОНП из печени удаляются ремнанты хиломикронов и триглицериды, образовавшиеся из свободных жирных кислот плазмы. Синтез ЛПОНП прямо коррелирует с повышением содержания свободных жирных кислот в гепатоцитах, что наблюдается при поступлении в организм больших количеств жиров с пищей, а в случаях усиления высвобождения адипоцитами свободных жирных кислот, которые поступают в кровоток (при ожирении, сахарном диабете, резистентном к терапии).

Аполипопротеин C-II на поверхности ЛПОНП активирует эндотелиальную липопротеинлипазу, которая расщепляет триглицериды до свободных жирных кислот и глицерина, которые используются жировой тканью, миокардом и скелетной мускулатурой в качестве энергетического субстрата. Остатки ЛПОНП превращаются в липопротеиды промежуточной плотности (ЛПП), которые затем частично удаляются печенью из кровотока и частично трансформируются в ЛПНП и тоже удаляются из кровотока. n Липопротеины низкой плотности (ЛПНП ) – мелкие частицы, которые являются основной транспортной формой холестерина. Они содержат около 6% триглицеридов, 50% холестерина и 22% белка. Примерно две трети быстрообменивающегося пула холестерина синтезируется в организме, преимущественно в печени (эндогенный холестерин) и одна треть поступает в организм с пищей (экзогенный холестерин). Ключевым ферментом, определяющим скорость синтеза эндогенного холестерина, является гидроксил метил-глутарил-Ко. А-редуктаза (ГМГ-Ко. А-редуктаза).

ЛПНП являются продуктом метаболизма ЛОНП и ЛППП, которые содержат наибольшее количество холестерина. Примерно 40 -60% всех ЛПНП захватываются гепатоцитами при участии аполипопротеина В и липопротеинлипазы печени. Второй путь катаболизма ЛПНП – свободнорадикальное перекисное окисление липидов, в результате которого образуются модифицированные ЛПНП. Последние захватываются макрофагами, которые трансформируются в пенистые клетки, входящие в состав атеросклеротических бляшек. Кроме того, модифицированные ЛППП вызывают повреждение сосудистого эндотелия. Липопротеины промежуточной плотности (ЛППП) – представляют собой ремнанты хиломикронов и ЛОНП, содержащие большое количество холестерина. ЛППП захватываются гепатоцитами или метаболизируются под влиянием липазы печени до ЛПНП (содержат апопротеин В).
 близок по своим физико-химическим свойствам к липопротеинам низкой плотности, отличаясь от них")
Липопротеин (а) близок по своим физико-химическим свойствам к липопротеинам низкой плотности, отличаясь от них наличием в оболочке дополнительного белка - апопротеина (а). Последний близок по своим свойствам к плазминогену и поэтому может конкурировать с ним за места связывания на фибрине и, таким образом, ингибировать фибринолитическую активность крови. Липопротеин (а) относится к числу атерогенных липопротеинов: их повышенный уровень в крови почти всегда ассоциируется с развитием атеросклероза, ИБС и высоким риском тромботических осложнений. n Липопротеины высокой плотности (ЛПВП) – самые мелкие и плотные частицы липопротеинов. Они содержат 5% триглицеридов, 22% холистерина 40% аполипопротеинов А-I, AII и С и относятся к липопротеидам, обладающими антиатерогенными свойствами.

Основной функцией ЛПВП является обратный транспорт холистерина из периферических органов, с поверхности хиломикронов и ЛОНП, макрофагов и гладкомышечных клеток, в печень, где происходят его утилизация и превращение в желчь. Синтез полноценных ЛПВП происходит при обязательном участии хиломикронов, ЛОНП и ЛПНП в энтероцитах и печени. Решающее значение для возникновения и прогрессирования атеросклероза имеет соотношение липопротеидов различных классов: ЛПНП, ЛОНП и липопротеин (а) обладают отчетливым атерогенным, а ЛПВП – антиатерогенным действием. Наиболее высокий риск развития атеросклероза наблюдается у лиц с высоким содержанием ЛПНП и ЛОНП и низким – ЛПВП.
 и их атерогенности обусловлены 1. Нарушением синтеза специфических")
Повышение содержания ЛПНП и липопротеина (а) и их атерогенности обусловлены 1. Нарушением синтеза специфических ЛПНП-рецепторов гепатоцитов, что препятствует элиминации холестерина печеночными клетками. n 2. Нарушением структуры и функции апопротеинов. n 3. Увеличением синтеза эндогенного холестерина. n 4. Увеличением количества модифицированных (окисленных) форм ЛПНП и липопротеина (а), образующихся, например, в результате перекисного окисления липидов. Факторы, способствующие снижению содержания ЛПВП n 1. Мужской пол. n 2. Ожирение. n 3. Гипертриглицеридемия. n 4. Высокое потребление углеводов. n 5. Сахарный диабет. n 6. Курение.
, характеризующиеся в первую очередь повышенным содержанием в крови холестерина и")
Нарушения липидного обмена (дислипидемии), характеризующиеся в первую очередь повышенным содержанием в крови холестерина и триглицеридов, являются важнейшими факторами риска атеросклероза и связанных с ним заболеваний сердечно-сосудистой системы. Концентрация в плазме крови общего холестерина (ХС) или его фракций, тесно коррелирует с заболеваемостью и смертностью от ИБС и других осложнений атеросклероза. Поэтому характеристика нарушений липидного обмена является обязательным условием эффективной профилактики сердечно-сосудистых заболеваний.

Для ориентировочной количественной оценки степени риска атеросклероза А. Н. Климовым в 1977 г. был предложен так называемый холестериновый коэффициент атерогенности Кхс, представляющий собой отношение холестерина (ХС) атерогенных и холестерина неатерогенных ЛП: К хс = ХС общий - ХС ЛПВП / ХС ЛПВПС Таким образом, важным моментом в функционировании ЛПВП является способность ЛПВП обмениваться холестеролом или его эфирами с липопротеидами других классов, циркулирующих в крови.

Существенную роль в этом обмене играет белок апо-Д, выступающий в качестве переносчика эфиров холестерола между отдельными липопротеидными частицами. В целом липопротеиды крови образуют единую липидтранспортную систему крови, ответственную за перенос липидов различных классов как эндогенного, так и экзогенного происхождения. Липопротеиды отдельных классов могут обмениваться между собой как липидными, так и белковыми компонентами. Поэтому нарушения обмена одного из классов липопротеидов обычно сопровождаются сдвигами в метаболизме липопротеидов других классов.

Интеграция метаболических путей обмена липидов Пищевые липиды являются источниками высших жирных кислот, глицерола, аминоспиртов и некоторых других соединений, используемых организмом для синтеза свойственных для него структурных или резервных липидов. Свободные высшие жирные кислоты, наряду с глицеролом и аминоспиртами образуются в организме также при расщеплении резервных или структурных липидов. Еще одним источником высших жирных кислот может служить их синтез из ацетила-Ко. А, который в свою очередь может быть промежуточным продуктом обмена углеводов или аминокислот. Заметим, что жирные кислоты в клетках используются в различных метаболических путях клетки только в их активированной форме - в виде ацил-Ко. А.

Одним из ключевых метаболитов липидного обмена является ацетил. Ко. А, поскольку, во-первых, именно через это соединение осуществляется окислительное расщепление высших жирных кислот; во-вторых, через ацетил-Ко. А атомы углерода жирных кислот могут быть использованы для пластических целей - для синтеза холестерола или полипреноидов; в третьих, через ацетил-Ко. А в гепатоцитах углеродные цепи жирных кислот преобразуются в ацетоновые тела гидрофильные "топливные" молекулы, легко транспортируемые в клетки различных органов и тканей; в четвертых, через ацетил-Ко. А осуществляются метаболические превращения углеродных скелетов аминокислот и моносахаридов в жирные кислоты, используемые в дальнейшем для синтеза более сложных липидных молекул.

Соединения других классов - аминокислоты и моносахариды - в ходе своего метаболизма образуют промежуточные продукты, которые могут в дальнейшем использоваться в клетке как для синтеза высших жирных кислот, так и для образования других мономерных единиц, необходимых для синтеза сложных липидов: глицерола, этаноламина, холина, сфингозина и пр. Таким образом, обмен липидов оказывается тесно связанным с обменом соединений других классов, а метаболические пути обмена липидов различных классов являются частью общей метаболической сети, функционирующей в организме.

Регуляция обмена липидов на уровне организма Липиды выполняют множество функций в организме, одной из важнейших среди них является обеспечение клеток различных органов и тканей энергией, т. к. от 30% до 40% всей необходимой ему энергии человек получает за счет окислительного расщепления соединений липидной природы. Интенсивность и направленность различных превращений липидов должны соответствовать потребности организма в энергетическом и пластическом материале. Поэтому крайне важными становятся как вопросы регуляции обмена липидов на уровне организма, так и проблема координации функционирования метаболических путей обмена липидов и метаболических путей обмена соединений других классов, обеспечивающих снабжение клеток необходимой им энергией. В конечном итоге, эффективная работа регуляторных и координирующих механизмов обеспечивает адаптацию организма к изменяющимся условиям его существования.

Так, в постабсорбционном периоде, когда поступление глюкозы и экзогенных липидов из кишечника во внутреннюю среду организма прекращается, потребность организма в энергии покрывается за счет расщепления резервных триглицеридов, основная масса которых сосредоточена в жировой ткани. В ходе мобилизация резервных триглицеридов образуются высшие жирные кислоты и глицерол, которые поступают из липоцитов вначале в кровь, а затем в клетки различных органов и тканей, где и окисляются с выделением необходимой клеткам энергии.

Этот процесс мобилизация резервных триглицеридов или липолиз стимулируется рядом гормонов, к числу которых относятся адреналин, норадреналин, глюкагон, bлипотропный гормон гипофиза, соматотропин, АКТГ, МСГ, кортизол, тироксин, тестостерон. Многие из этих гормонов являются активаторами гормон-чувствительной липазы липоцитов (триацилглицероллипазы). Для оптимального протекания большинства липолитических процессов необходимо присутствие кортизола, соматотропина и гормонов щитовидной железы. Сами по себе эти гормоны не оказывают прямого влияния на липолиз, а действуют как факторы, стимулирующие действие других гормонов.

Важнейшая роль в мобилизации резервных липидов в организме человека принадлежит адреналину ( вместе с норадреналином ), который выделяется в жировой ткани нервными окончаниями симпатической нервной системы. Вторым источником адреналина является мозговое вещество надпочечников, откуда адреналин доставляется в жировую ткань с током крови. Вероятно, адреналин из мозгового вещества надпочечников играет важную роль в мобилизации триглицеридов жировой ткани в условиях острого эмоционального стресса. Адреналин взаимодействует со своим рецептором на наружной поверхности мембраны липоцита с образованием гормонрецепторного комплекса.

В ответ на образование гормон-рецепторного комплекса с помощью специального механизма происходит активация расположенной на внутренней поверхности наружной клеточной мембраны липоцита аденилатциклазы - фермента, синтезирующего из АТФ циклическую АМФ (ц. АМФ). Увеличение внутриклеточной концентрации ц. АМФ активирует фермент протеинкиназу, которая осуществляет активацию триацилглицероллипазы путем ее фосфорилирования, т. е. путем ковалентной модификации фермента. Поскольку скорость липолиза лимитируется активностью триацилглицероллипазы, активация фермента приводит к ускорению гидролиза резервных триглицеридов и увеличению выхода высших жирных кислот и глицерола из липоцита в русло крови.

Гормоны глюкагон, b-липотропин, меланоцит стимулирующий гормон, кортикотропин активируют липолиз в жировой ткани, увеличивая концентрацию ц. АМФ в липоцитах с помощью механизма, сходного с механизмом активации липолиза под действием адреналина. Интересно, что существуют видовые различия в эффективности функционирования этих регуляторных механизмов: так, у птиц глюкагон является мощным стимулятором липолиза, тогда как липолитический эффект глюкагона у человека крайне незначителен. Соматотропный гормон не оказывает прямого влияния на скорость расщепления триглицеридов в липоцитах, однако соматотропин увеличивает скорость синтеза аденилатциклазы за счет ускорения процесса транскрипции соответствующего гена. Увеличение содержания аденилатциклазы в липоцитах увеличивает эффект воздействия на жировую ткань таких гормонов как адреналин, b-липотропин и др.

Сходным образом оказывает стимулирующее влияние на липолиз и кортизол, поскольку этот гормон увеличивает содержание в липоцитах другого фермента - гормон-чувствительной липазы. Кортизол выступает в качестве стимулятора транскрипции гена, ответственного за синтез этого фермента. Повышение же содержания триацилглицероллипазы в липоцитах способствует более быстрому и более выраженному ответу клеток на воздействие на них гормонов типа адреналина. Механизм действия тироксина на жировую ткань не совсем ясен. Известно, что этот гормон способствует более эффективной передаче стимулирующего сигнала с гормон-рецепторного комплекса на аденилатциклазу, в результате чего при воздействии на липоциты гормонов типа адреналина происходит более быстрая активация липолиза в этих клетках.

Основным гормоном, тормозящим липолиз в жировой ткани, является инсулин. Инсулин снижает содержание ц. АМФ в липоцитах, по-видимому, за счет активации фосфодиэстеразы, переводящей ц. АМФ в обычную АМФ. Снижение концентрации ц. АМФ в клетках приводит как к инактивации протеинкиназы, так и к активации фосфопротеинфосфатазы, в результате чего происходит дефосфорилирование гормон-чувствительной липазы с ее инактивацией и последующим торможением липолиза. Простагландины также снижают содержание ц. АМФ в липоцитах с последующим торможением липолиза в клетках.

В период абсорбции в клетках различных органов и тканей активно идет липогенез. Во внутреннюю среду организма из кишечника поступают глюкоза и другие моносахариды, а также триацилглицерины в составе ХМ или ЛПОНП. Моносахариды, поступающие в липоциты или в гепатоциты, используются в ходе липогенеза, являясь как источниками ацетил -Ко. А для синтеза высших жирных кислот, так и источниками фосфотриоз, необходимых для образования 3 -фосфоглицерола. Триглицериды ХМ или ЛПОНП после их гидролиза липопротеидлипазой также являются источниками высших жирных кислот и глицерола, поступающих в клетки и в дальнейшем используемыми в качестве субстратов для липогенеза.

Гормоном, стимулирующим липогенез, является инсулин. Инсулин ускоряет поступление глюкозы в клетки и стимулирует ее фосфорилирование, запуская тем самым процесс утилизации глюкозы в клетках. Причем стимулируется как процесс аэробного окисления глюкозы до СО 2 и Н 2 О, так и работа пентозного цикла окисления глюкозы, обеспечивающего клетки восстановительными эквивалентами в виде НАДФН+Н+. Инсулин активирует работу пируватдегидрогеназного комплекса, что приводит к увеличению образования ацетил-Ко. А - исходного субстрата для синтеза высших жирных кислот. Инсулин повышает активность фермента ацетил-Ко. А-карбоксилазы, катализирующего превращение ацетил-Ко. А в малонил-Ко. А, также необходимого для синтеза высших жирных кислот.

Ускорение окислительного распада глюкозы в клетке приводит также к увеличению в ней концентрации фосфотриоз - 3 фосфоглицеринового альдегида и фосфодигидроксиацетона, используемых для образования 3 -фосфоглицерола. Таким образом, воздействие инсулина на клетки приводит к наработке в них исходных соединений для синтеза триглицеридов. Кроме того, инсулин активирует в клетках глицеролфосфатацилтрансферазу - фермент, катализирующий перенос ацильного остатка с Ко. А на 3 -фосфоглицерол первую реакцию метаболического пути синтеза триацилглицеринов.

Регуляция активности пируватдегидрогеназного комплекса, ацетил-Ко. А-карбоксилазы и глицеролфосфат-ацилтрансферазы осуществляется путем координированного процесса ковалентной модификации этих ферментов (фосфорилированиедефосфорилирование ). В целом, воздействие инсулина на липоциты приводит, во-первых, к торможению липолиза в клетках, а, вовторых, к активации в них процесса липогенеза, способствуя тем самым накоплению энергетических резервов в организме в виде триацилглицеринов.

Образование хиломикронов Жиры, образовавшиеся в результате ресинтеза в клетках слизистой оболочки кишечника, упаковываются в ХМ. Основной апопротеин в составе ХМ - белок апо. В-48. Этот белок закодирован в том же гене, что и белок ЛПОНП - В-100, который синтезируется в печени. В кишечнике в результате посттранскрипционных превращений "считывается" последовательность м. РНК, которая кодирует только 48% от длины белка В 100, поэтому этот белок называется апо. В-48. Белок апо. В-48 синтезируется в шероховатом ЭР и там же гликозилируется. Затем в аппарате Гольджи происходит формирование ХМ, называемых "незрелыми". По механизму экзоцитоза они выделяются в хилус, образующийся в лимфатической системе кишечных ворсинок, и через главный грудной лимфатический проток попадают в кровь. В лимфе и крови с ЛПВП на ХМ переносятся апопротеины Е (апо. Е) и С-П (апо. С-П); ХМ превращаются в "зрелые". ХМ имеют довольно большой размер, поэтому после приёма жирной пищи они придают плазме крови опалесцирующий, похожий на молоко, вид. ХМ транспортируют жир к различным тканям, где он утилизируется, поэтому концентрация ХМ в крови постепенно снижается, и плазма опять становится прозрачной. ХМ исчезают из крови в течение нескольких часов.

При редком наследственном заболевании - дефекте гена апопротейна В - нарушается синтез белков апо. В-100 в печени и апо. В-48 в кишечнике. В результате в клетках слизистой оболочки кишечника не формируются ХМ, а в печени - ЛПОНП. В клетках этих органов накапливаются капельки жира. Такое заболевание называется абеталипопротеинемия, так как второе название ЛПОНП - пре-β-липопротеины. Действие липопротеинлипазы на ХМ. В крови триацилглицеролы, входящие в состав зрелых ХМ, гидролизуются ферментом липопротеинлипазой, или ЛП-липазой (рис. 8 -20). ЛП-липа-за связана с гепарансульфатом (гетерополисаха-ридом), находящимся на поверхности эндотелиальных клеток, выстилающих стенки капилляров кровеносных сосудов. ЛП-липаза гидролизует молекулы жиров до глицерола и 3 молекул жирных кислот. На поверхности ХМ различают 2 фактора, необходимых для активности ЛП-липазы - апо. С-П и фосфолипиды. Апо. С-П активирует этот фермент, а фосфолипиды участвуют в, связывании фермента с поверхностью ХМ.

ЛП-липаза синтезируется в клетках многих тканей: жировой, мышечной, в лёгких, селезёнке, клетках лактирующей молочной железы. Изоферменты ЛП-липазы в разных тканях отличаются по значению Кm: ЛП-липаза жировой ткани имеет в 10 раз более высокое значение Кm, чем, например, ЛП-липаза сердца, поэтому гидролиз жиров ХМ в жировой ткани происходит в абсорбтивный период. Жирные кислоты поступают в адипоциты и используются для синтеза жиров. В постабсорбтивном состоянии, когда количество жиров в крови снижается, ЛП-липаза сердечной мышцы продолжает гидролизовать жиры в составе ЛПОНП, которые присутствуют в крови в небольшом количестве, и жирные кислоты используются этой тканью как источники энергии, даже при низкой концентрации жиров в крови. ЛП-липазы нет в печени, но на поверхности клеток этого органа имеется другой фермент - печёночная липаза, не действующая на зрелые ХМ, но гидролизующая жиры в ЛППП, которые образуются из ЛПОНП.

В результате действия ЛП-липазы на жиры ХМ образуются жирные кислоты и глицерол. Основная масса жирных кислот проникает в ткани (рис. 8 -20). В жировой ткани в абсорбтивный период жирные кислоты депонируются в виде триацилглицеролов, в сердечной мышце и работающих скелетных мышцах используются как источник энергии. Другой продукт гидролиза жиров, глицерол, растворим в крови, транспортируется в печень, где в абсорбтивный период может быть использован для синтеза жиров.

Рис. 8 -20. Путь экзогенных жиров и хиломикронов. *ЛПЛ - липопротеинлипаза, ЖК - жирные кислоты.

В результате действия ЛП-липазы на ХМ количество жиров в них снижается на 90%, уменьшаются размеры частиц, апопротеин С-П переносится обратно на ЛПВП. Образовавшиеся частицы называются остаточными ХМ. Они содержат в себе фосфолипиды, холестерол, жирорастворимые витамины и апопротеины В 48 и Е. Остаточные ХМ захватываются гепатоцитами, которые имеют рецепторы, взаимодействующие с этими апопротеинами. Путём эндоцитоза остаточные ХМ попадают внутрь клеток, и ферментами лизосом белки и липиды гидролизуются, а затем утилизируются. Жирорастворимые витамины и экзогенный холестерол используются в печени или транспортируются в другие ткани. Скорость удаления ХМ из кровотока зависит от: n активности ЛП-липазы; n присутствия ЛПВП, поставляющих апопротеины С-II и Е для ХМ; n активности переноса апо. С-II и апо. Е на ХМ. n Генетические дефекты любого из белков, участвующих в метаболизме ХМ, приводят к развитию семейной гиперхиломикронемии - гиперлипопротеинемии типа I. У таких больных в постабсорбтивном периоде концентрация триацилглицеролов повышена (более 200 мг/дл), плазма крови по виду напоминает молоко и при оставлении на холоде (+4 °С) в ней всплывают белые жирные хлопья, что характерно для гипертриглицеролемии и гиперхиломикронемии.

Метаболизм ЛПОНП и ЛПНП Основная масса липопротеидов очень низкой плотности или ЛПОНП образуется в печени и участвует в транспорте синтезированных в гепатоцитах липидов к клеткам различных органов или тканей, т. е. участвует в транспорте эндогенных липидов. Небольшая часть ЛПОНП, как мы уже упоминали, образуется в кишечнике, принимая участие в транспорте экзогенных липидов. Образование ЛПОНП в гепатоцитах начинается с синтеза апопротеина В 100 на рибосомах, параллельно в гладком эндоплазматическом ретикуме идет синтез липидных компонентов ЛПОНП: триглицеридов, фосфолипидов и холестерола. Комплексы, состоящие из апо-протеинов и синтезированных липидов, представляющие собой так называемые насцентные ЛПОНП, поступают в аппарат Гольджи, где белки подвергаются гликозилированию, а затем путем обратного пиноцитоза поступают в кровяное русло. В русле крови к насцентным ЛПОНП присоединяются апобелки апо-С и апо -Е, источником которых, вероятно являются липопротеиды других классов, уже циркулирующие в крови. В результате обогащения апобелками насцентные ЛПОНП превращаются в зрелые ЛПОНП.

Катаболизм ЛПОНП начинается на поверхности эндотелия капилляров периферических органов и тканей, куда они доставляются током крови. Под действием имеющейся на клетках эндотелия липопротеидлипазы происходит гидролиз триглицеридов ЛПОНП с образованием глицерола и высших жирных кислот. Продукты гидролиза поступают в клетки органов и тканей, хотя часть их может уносится током крови в другие органы. Потеряв в ходе воздействия на них липопротеидлипазы большую часть своих триглицеридов ЛПОНП превращаются в липопротеиды промежуточной плотности (ЛППП). Примерно 50% образовавшихся ЛППП захватываются печенью с помощью имеющихся в гепатоцитах В, Е-рецепторов. Эти рецепторы способны избирательно связать липопротеидные частицы, в составе которых имеются апопротеины В или апопротеины Е. К ним, в частности, относятся и ЛППП. После рецепторного захвата ЛПППрецепторные комплексы поступают в гепатоциты, где и расщепляются. Вторая часть ЛППП превращается в русле крови в ЛПНП, что было однозначно доказано с помощью введения в кровь меченых по апо-В ЛПОНП, поскольку введенная метка обнаруживалась вначале в ЛППП, а затем в ЛПНП. Механизм преобразования ЛППП в ЛПНП окончательно не выяснен.

Наиболее вероятным является предположение, согласно которому ЛППП подвергаются в русле крови воздействию еще одного фермента - печеночной триглицеридлипазы (гепарин-освобождаемая липаза печени), под действием которой продолжается гидролиз триглицеридов с снижением их содержания в липопротеидной частице в конечном итоге в среднем до 8%. В результате ЛППП превращаются в ЛПНП. В ходе преобразования ЛППП в ЛПНП происходят также изменения в апопротеидном составе липопротеидных частиц: ЛППП практически полностью теряют апо-С и апо-Е, которые, повидимому, переносятся на липопротеидные частицы других классов. При утилизации ЛПНП в клетках периферических органов и тканей на первом этапе ЛПНП проникают из кровяного русла в межклеточное пространство или путем активного переноса через клетки эндотелия, или же через межэндотелиальные щели. Затем ЛПНП взаимодействуют с рецепторами ЛПНП (В-рецепторы) на поверхности клеток разного типа. Образующийся комплекс поглощается клетками и поступает в лизосомы, где и происходит его полное разрушение, а продукты расщепления ЛПНП используются клетками.

ЛПНП снабжают клетки периферических органов и тканей в основном холестеролом ( 50% массы ЛПНП ) и в какой-то мере фосфолипидами ( 22% массы ЛПНП ). Фосфолипиды, возможно, используются клетками для построения или обновления своих мембран. Холестерол, поступающий в составе ЛПНП, также используется в клетках для построения мембран. Избыточный холестерол подвергается этерификации при участии фермента ацил-Ко. А-холестерол ацилтрансферазы (АХАХ) и резервируется в клетке в виде вакуолей, содержащих преимущественно олеиновые эфиры холестерола. Поступающие с ЛПНП белки и триглицериды гидролизуются, а продукты их расщепления: аминокислоты, глицерол и высшие жирные кислоты утилизируются клетками. Избыток холестерола в мембранах клеток нарушает их микровязкость и нарушая тем самым работу трансмембранных транспортных систем. Другими словами, избыток холестерола оказывает на клетки токсический эффект. Клетки периферических тканей располагают несколькими механизмами, предотвращающими избыточное накопление холестерола в их мембранах. Вопервых, при избыточном поступлении холестерола в клетку за счет рецепторопосредованного захвата ЛПНП, количество В-рецепторов на поверхности клетки уменьшается.

Во-вторых, излишнее накопление холестерола в мембранах тормозит работу собственного механизма синтеза холестерола в клетке путем угнетения активности ГМГ-редуктазы. Наконец, в третьих, избыток холестерола активирует работу АХАТ, переводя тем самым молекулы свободного холестерола в его эфиры, последние же резервируются в клетке в составе специальных вакуолей. В печени ситуация несколько иная, так как холестерол, поступающий в гепатоциты вместе с ЛПНП, может или использоваться в ходе синтеза новых липопротеидных частиц, или может превращаться в желчные кислоты, или же может секретироваться в желчь и выводиться вместе с ней в кишечник. Следует отметить, что в последнее время появилось представление о двух раздельных пулах холестерола в гепатоцитах. Один пул формируется за счет его синтеза и используется для образования липопротеидов различных классов. Другой пул формируется в основном за счет холестерола, поступающего в гепатоциты из крови в составе ЛППП, ЛПНП и ЛПВП; этот холестерол или используется для синтеза желчных кислот, или секретируется в желчь. В целом же система липопротеидных частиц ЛПОНП Д> ЛППП Д> ЛПНП обеспечивает транспорт липидов, синтезированных в печени, в клетки периферических органов и тканей.
, циркулирующих в крови, формируется из трех")
Метаболизм ЛПВП Общий пул липопротеидов высокой плотности (ЛПВП), циркулирующих в крови, формируется из трех источников: за счет их образования в печени, за счет их поступления из кишечника и за счет их образования из ремнантных хиломикронов. При образовании ЛПВП в печени вначале из липидов и апобелков, главным из которых является апо-А, формируются насцентные дисковидные липопротеидные частицы. Существенными, хотя и минорными белковыми компонентами ЛПВП являются апо-Е, апо-С и фермент лецитин: холестерол -ацилтрансфераза (ЛХАТ). ЛХАТ катализирует реакцию между расположенными в наружном монослое липопротеида фосфолипидом и холестеролом с образованием эфира холестерола. Эфиры холестерола являются полностью гидрофобными молекулами, вследствие чего они переходят из внешнего монослоя частицы в ее гидрофобное ядро. Дисковидная частипостепенно превращается в зрелый сферический ЛПВП, поступающий в кровоток.

ЛПВП являются липопротеидными частицами, осуществляющими захват избыточного холестерола из мембран клеток периферических тканей и транспортирующими этот холестерол или в печень, или в кишечник. В печени этот холестерол превращается в желчные кислоты, а его избыток может секретироваться гепатоцитами непосредственно в желчь и далее поступать в просвет кишечника. Поступивший вместе с ЛПВП в стенку кишечника холестерол или используется для синтеза хиломикронов и ЛПОНП, или же может секретироваться в просвет кишечника. В любом случае функционирование ЛПВП будет способствовать выведению излишнего холестерола из организма. Важнейшую роль в акцепторной функции ЛПВП по отношению к холестеролу клеточных играет фермент ЛХАТ катализирует реакцию превращения свободного холестерола, входящего в состав амфифильной оболочки липопротеида, в его эфир, который будучи полностью гидрофобным. погружается из оболочки ЛПВП в его ядро, освобождая таким образом место в оболочке для связывания новой молекулы холестерола, источником которой и служат мембраны клеток, с которыми контактирует ЛПВП.

Возможно, что доставку свободного холестерола из клеточных мембран на ЛПВП осуществляют специальные белки ( или белок ) - переносчики холестерола. В ходе процесса, катализируемого ЛХАТ происходит обогащение ЛПВП холестеролом. Подтверждением реальности этого процесса является наличие в плазме двух фракции ЛПВП - ЛПВП 2 и ЛПВП 3 , которые различаются по содержанию холестерола: в ЛПВП 3 холестерола в среднем около 17%, а в ЛПВП 2 - около 23%. В таком случае ЛПВП 3 поступают в кровь из печени или из кишечника, захватывают холестерол из клеточных мембран, переходя в ЛПВП 2 , а ЛПВП 2 поглощаются клетками печени или кишечника. В основе поглощения ЛПВП печенью лежит рецепторопосредованный их захват гепатоцитами с помощью имеющихся на мембранах клеток В, Е-рецепторов, поскольку известно, что в составе ЛПВП имеется апо-Е. Возможен другой вариант поступления холестерола с ЛПВП в гепатоциты: на поверхности гепатоцитов имеется специальный фермент гепарин-освобождаемая липаза ( ГОЛП ). Этот фермент катализирует расщепление фосфолипидов ЛПВП при их контакте с поверхностью гепатоцита. В результате этого расщепления в наружном монослое ЛПВП нарушается баланс между количеством холестерола и фосфолипидов, который восстанавливается за счет перехода части холестерола, ставшего избыточным, с ЛПВП в гепатоцит.
липиды.ppt