10319022ed50799bd48e489d55ef3a0f.ppt
- Количество слайдов: 89



Летняя школа «Теоретические и прикладные проблемы когнитивной психологии» , посвященная 110 -летию со дня рождения Карла Дункера 31 августа - 4 сентября 2013 Психофизиология внимания Б. В. Чернышев bchernyshev@hse. ru зав. лаб. когнитивной психофизиологии НИУ ВШЭ http: //psy. hse. ru/cognpp/ доцент каф. высшей нервной деятельности МГУ

Тезисы лекции Начиная с У. Джеймса, внимание понимается в психологии как ворота в сознание, осуществляющее селективный допуск репрезентации различных объектов и событий в сознание. В когнитивной психологии был предложен ряд теоретических моделей, объясняющих внимание в традиционных для когнитивной психологии понятиях потоков сигналов и обработки информации. В этих теориях внимание рассматривается либо как фильтр, отсекающий часть сигналов, либо как усилие, обеспечивающее выделение ресурсов на обработку информации, либо как перцептивное действие. Основные классические модели внимания были построены в когнитивной психологии на основе экспериментов с анализом времени реакции, точности воспроизведения информации и других поведенческих показателей. Психофизиологические исследования внимания, основанные на исследовании активности мозга, были начаты в 60 -х гг. XX века. В 1965 г. С. Саттон впервые применил методику одд-болл (парадигму необычного стимула), ставшую "золотым стандартом" психофизиологии внимания, и открыл компонент вызванного потенциала P 300. Уже почти полвека P 300 активно исследуется, и подтверждается его связь со вниманием и осознанием. Исследования С. А. Хильярда 1973 года, в которых совмещались методика оддболл и дихотическое прослушивание, показали усиление компонента вызванного потенциала N 100 при произвольном внимании к каналу поступления информации [Hillyard et al. , 1973]. Интерпретация результатов основывалась на теориях фильтров. Вскоре в дискуссию с С. А. Хильярдом включился другой известнейший психофизиолог, Р. Наатанен, который в 1978 году также провел эксперименты с дихотическим прослушиванием, но выдвинул другую интерпретацию, которая по своей сути ближе к ресурсной теории, чем к теории фильтра [Näätänen et al. , 1978].
 Начиная с конца 80 -х гг. психофизиология в дополнение к классическому")
Тезисы лекции (продолжение) Начиная с конца 80 -х гг. психофизиология в дополнение к классическому арсеналу электрофизиологических методов получила в свое распоряжение новые современные методы исследования активности мозга, позволяющие точно локализовать активность в мозге – что, в свою очередь, привело к появлению ряда ставших классическими психофизиологических теорий внимания. Эти теории основываются на выделении нескольких взаимосвязанных систем внимания, и подчеркивают взаимодействие лобных и теменных областей мозга, а также роль активационных систем мозга: это, прежде всего теории внимания М. Познера (Posner, Di. Girolamo, 2000), М. Мезулама (Mesulam, 1998), М. Корбетты (Corbetta, Shulman, 2002) и Д. Лаберджа (La. Berge, 2000). Отдельного внимания заслуживает премоторная теория внимания Д. Риццолатти (Rizzolatti et al. , 1987), основанная на изучении программирования движений глаз. Важную роль в современном психофизиологическом понимании внимания уделяют синхронизации в работе нейронных сетей, которая обеспечивает связывание признаков в целостное восприятие – функцию, тесно связанную со вниманием, а также фильтрацию информации. В данном аспекте изучена синхронизация на нескольких ритмах, включая альфа- и гамма-диапазоны (Basar et al. , 2001; Мачинская, 2003; Singer, 2003; Klimesch et al. , 2007).

Что такое внимание?

Внимание – «это овладение умом в четкой и яркой форме одним из нескольких наличествующих объектов. Суть этого процесса – фокусирование, концентрирование сознания. Внимание приводит к отвлечению от некоторых вещей для того, чтобы можно было эффективно заняться другими. Состояние внимания противоположно рассеянному, затемненному состоянию. » Уильям Джеймс (James, 1890)
![Основные подходы к вниманию в когнитивной психологии [По Фаликман, 2006]](https://present5.com/presentation/10319022ed50799bd48e489d55ef3a0f/image-6.jpg "Основные подходы к вниманию в когнитивной психологии [По Фаликман, 2006]")
Основные подходы к вниманию в когнитивной психологии [По Фаликман, 2006]
![Расположение фильтров в моделях ранней и поздней фильтрации [Massaro, 1975 по Николаева, 2003]](https://present5.com/presentation/10319022ed50799bd48e489d55ef3a0f/image-7.jpg "Расположение фильтров в моделях ранней и поздней фильтрации [Massaro, 1975 по Николаева, 2003]")
Расположение фильтров в моделях ранней и поздней фильтрации [Massaro, 1975 по Николаева, 2003]

Модель распределения ресурсов по Д. Канеману Согласно Д. Канеману, количество выделяемых ресурсов зависит от уровня активации. Причем имеющееся количество ресурсов может быть выделено на одну задачу, или ресурсы могут быть в той или иной пропорции распределены между несколькими задачами. [По: Канеман, 2006] Отчего внимания к важному стимулу может не хватить? ● Внимание направлено на что-то другое (ресурсы отданы другой задаче) ● Внимание снижено, т. к. недостаточен уровень активации мозга (ресурсы не выделены вообще)

Отчего внимания к важному стимулу может не хватить?


Непроизвольное внимание связано с рефлекторными установками. Оно устанавливается и поддерживается независимо от сознательного намерения человека. …Произвольное внимание - это сознательно направляемое и регулируемое внимание, в котором субъект сознательно избирает объект, на который оно направляется. … Непроизвольное внимание обычно представляется как пассивное, произвольное - как активное (Джеймс). Первое направляют независящие от нас факторы: внезапно раздавшийся шум, яркая окраска, ощущение голода; второе направляем мы сами. С. Л. Рубинштейн. Основы общей психологии. СПб. , 1998.

В современной нейрофизиологической литературе выделяют две приблизительно аналогичных понятия – две формы индукции внимания (хотя понятие «сознание» здесь намеренно не используется): Восходящее влияние ( «bottom-up» influence) - под воздействием сенсорной стимуляции процесс распространяется от структур низкого уровня (в том числе стволовые сенсорные образования и стволовая ретикулярная формация) вверх (на кору больших полушарий). Нисходящее влияние ( «top-down» influence) - высшие области (префронтальные и другие области коры больших полушарий) запускают процесс, распространяющийся на мозговые структуры более низкого уровня.
 Functional components of attention. Processes that contribute to attention")
Функциональные компоненты внимания (Knudsen, 2007) Functional components of attention. Processes that contribute to attention are shown in red. Information about the world (green ellipse) is transduced by the nervous system and is processed by salience filters that respond differentially to infrequent or important stimuli (bottom-up). Neural representations in various hierarchies encode information about the world, movements, memories, the animal’s emotional state, etc. A competitive process selects the representation with the highest signal strength for entry into the circuitry that underlies working memory. Working memory can direct top-down bias signals that modulate the sensitivity of representations that are being processed in working memory. The selection process can also direct top-down bias signals that reflect the result of the competitive selection. Working memory and competitive selection direct eye movements and other orienting behaviors that modify the effects of the world on the animal’s nervous system. Corollary discharges associated with gaze control modulate sensitivity control. Voluntary attention involves working memory, top-down sensitivity control, and competitive selection operating as a recurrent loop (dark arrows).
 – автоматизированные")
Стадии обработки информации Предвнимание (Neisser, 1967; Treisman, Gelade, 1980 и др. ) – автоматизированные процессы, выполняющиеся параллельно, позволяют одновременно обрабатывать множество единиц информации без явной конкуренции за ресурсы. В небольших пределах может модулироваться в зависимости от сознательной установки. внимание – всегда связано с селекцией информации ( «фокус внимания» ), подчиняется ресурсным ограничениям, тесно связано с текущим содержанием сознания. Собственно фокальное Для слуховой модальности переход между предвнимательной и внимательной стадиями происходит примерно около 200 мс после предъявления стимула.

Что такое мозг?

Френологические карты Ф. Галля

Металлический микроэлектрод в зрительной коре обезьяны.

Магниторезонансная томография
")
Магнитоэнцефалография (МЭГ)

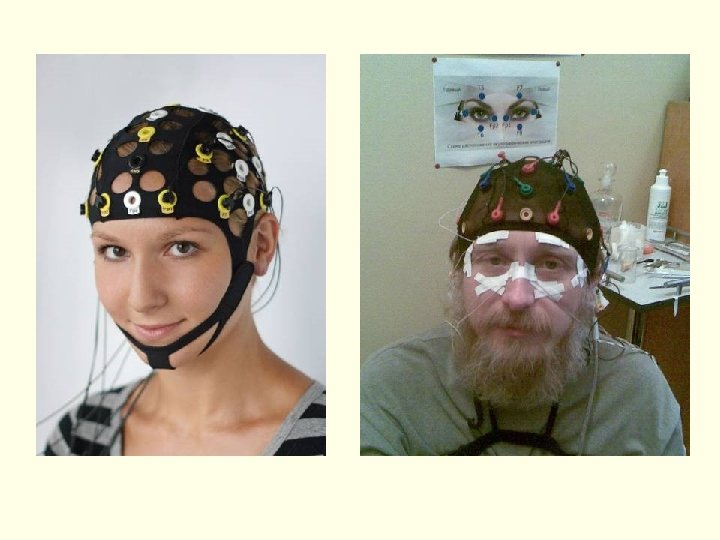
Энцефалограмма
 -5 N 2 Potential ( 0 5 P 2")
-10 N 1 m V) -5 N 2 Potential ( 0 5 P 2 P 3 10 -200 0 200 400 600 Time (ms) Слуховые вызванные потенциалы

Проявления внимания в вызванных потенциалах
 - парадигма необычного стимула (Sutton et al. , 1965): Активный одд-болл: испытуемый")
Одд-болл (oddball) - парадигма необычного стимула (Sutton et al. , 1965): Активный одд-болл: испытуемый должен считать про себя или нажимать на кнопку в ответ на редкий ( «девиантный» ) целевой стимул S 1 и игнорировать частый ( «стандартный» ) нецелевой стимул S 2, S 1, S 2, S 2, S 1, S 2… Пассивный одд-болл: происходит аналогично, но испытуемому не дают никакой инструкции.

Risto Näätänen University of Helsinki

Steven Hillyard Department of Neurosciences University of California at San Diego
 – mismatch negativity (MMN) (Открыта Р. Наатаненом) – разность между девиантным")
Негативность рассогласования (НР) – mismatch negativity (MMN) (Открыта Р. Наатаненом) – разность между девиантным и стандартным стимулами в одд-болле. Достигает максимума между 160 – 220 мс, максимально выражена в центральных отведениях по средней линии. Может накладываться на N 1, P 2 и N 2. Иногда НР называют N 2 a (в этом случае классический N 2 получает название N 2 b). Негативность рассогласования является автоматическим процессом сравнения текущего стимула со следом от предыдущего. Она возникает независимо от инструкции или от произвольного внимания испытуемого (предвнимание). Тем не менее, НР подавляется на сигналы в одно ухо, если испытуемый активно направляет внимание на сигналы в другом ухе.

Негативность рассогласования Зависимость негативности рассогласования от разницы по высоте тона между стандартным и девиантным стимулом (Naatanen et al. , 2007). Испытуемые различают стимулы только для 1016 и 1032 Гц (две нижних записи).
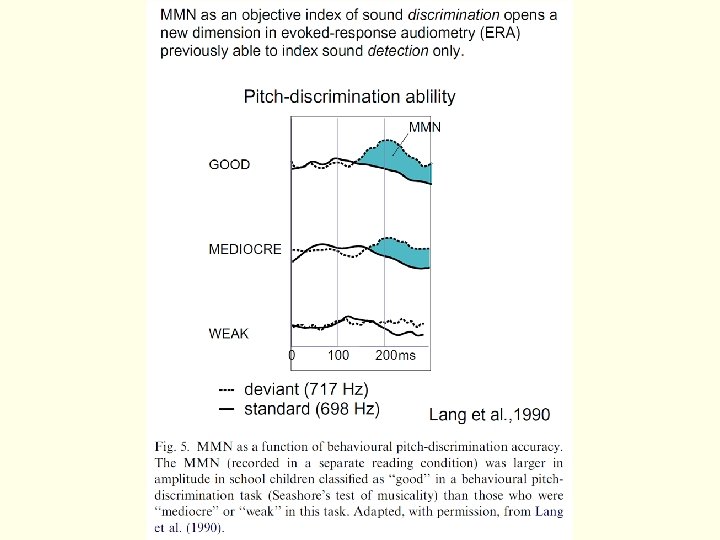
 – processing negativity (PN) (Наатанен) – разность между нецелевым стимулом в")
Процессная негативность (ПН) – processing negativity (PN) (Наатанен) – разность между нецелевым стимулом в значимом (релевантном) канале и нецелевым же стимулом в незначимом (нерелевантном) канале. Для таких экспериментов требуется иметь два потока стимулов – на один из них испытуемый обращает внимание (например, в одно ухо), а второй игнорируется им (во второе ухо). Иными словами, нужны четыре стимула, которые образуют два параллельно идущих одд-болла, один из которых по инструкции активный, а другой – игнорируемый. Либо производится сравнение активного и пассивного одд-болла (при чтении книги). По времени примерно соответствует негативности рассогласования. Разностная негативность (РН) (Nd) (Хилльярд) – фактически примерно то же самое, что процессная негативность по Наатанену (открыто раньше!). Однако Наатанен подводит теоретическую базу под их различия.
. «Внимаемый» и игнорируемый каналы различаются по высоте")
Процессная негативность Разностная негативность (Hillyard, Kutas, 1983). «Внимаемый» и игнорируемый каналы различаются по высоте тона. Девиантный стимул немного больше по длительности стандартного в обоих каналах.

Процессная негативность N 1 P 2 N 2 P 3 Процессная негативность (Naatanen, Gaillard, Mantysalo, 1978). Методика: Вызванные потенциалы на звуковые стимулы. Дихотическое прослушивание: в каждое ухо подается последовательность из частых «стандартов» и редких «сигналов» , привлекающих внимание. Инструкция требует от испытуемого слушать только в одно «канале» информации (обращать внимание только на звуки в одном ухе). Процессная негативность – негативный потенциал, накладывающийся на вызванный потенциал в состоянии внимания в сравнении с состоянием невнимания. Хорошо виден для «стандартов» . Связан с повышенной обработкой информации в том канале, на который обращено внимание.

Оба негативных отклонения – как негативность рассогласования, так и процессная негативность – указывают на усиление обработки информации в мозге.

P 3, или P 300 P 3 возникает через 250 - 500 мс и более после включения стимула. P 3 a – возникает при непроизвольном внимании. Имеет фронтальное распределение на скальпе. P 3 b – возникает при произвольном внимании. Имеет фронтопариетальное распределение. Т. е. его максимум расположен кзади по отношению к максимуму P 3 a, а латентность больше. P 3 может возникнуть на пропущенный стимул, ожидавшийся испытуемым – т. е. в отсутствии реального физического стимула. P 3 связывают с «обновлением контекста памяти» , осознанием стимула, его классификацией и др. (осознание подразумевает, что испытуемый может дать эксплицитный отчет о стимуле). Латентность P 3 увеличивается при повышении трудности задачи. Однако латентность P 3 не связана с латентностью поведенческой реакции испытуемого! Поведенческая реакция испытуемого может иногда проявиться даже раньше P 3!

P 3, или P 300 N 1 N 2 P 3 Вызванные потенциалы на стимулы при отсутствии внимания (синяя линия) и при наличии внимания к стимулу (красная линия) – методика одд-болл. Обратите внимание, что линии не совпадают и в предстимульном интервале (из-за случайных процессов)!

Поиск систем внимания в мозге на основе томографических, энцефалографических, нейропсихологических методов и регистрации нейронной активности

Michael Posner Department of Psychology, Institute of Cognitive and Decision Sciences Institute of Neuroscience University of Oregon

Maurizio Corbetta Dept. of Neurology Washington University in St. Louis

Marsel Mesulam Cognitive Neurology and Alzheimer's Disease Center Institute of Neuroscience Northwestern University

Экспериментальная парадигма М. Познера

Временная динамика экзогенной и эндогенной ориентировки внимания в парадигме М. познера
 (Posner, Peterson, 1990)")
Передняя система внимания (система исполнительного контроля) (Posner, Peterson, 1990)
 (Posner, Peterson, 1990)")
Задняя система внимания (система ориентирования) (Posner, Peterson, 1990)

Экспериментальная схема для выявления сетей внимания, основанная на методике М. Познера (Fan et al. , 2002)
")
Согласно точке зрения М. Познера (сформировавшейся на основе томографических и энцефалографических экспериментов на людях) в мозге человека существуют три нервные сети внимания: Сеть Функция Структуры мозга Сеть Выбор цели и исполнительного управление контроля поведением, направленным на эту цель; детекция ошибок; разрешение конфликтов; подавление рефлекторных реакций Медиальные области лобной доли, включая переднюю часть поясной извилины и дополнительную моторную область; Передняя дорсолатеральная область лобной доли (префронтальная кора); Базальные ганглии (стриарная система) – в особенности хвостатое ядро Сеть пробуждения Состояние бодрствования и готовности к действиям Правая лобная доля (в особенности верхняя часть поля 6); Правая теменная доля; Голубое пятно (источник норадреналина); Холинергические ядра (добавлено мной - БЧ) Сеть ориентирования Ориентировочная реакция на сенсорные стимулы Теменная доля; Веретеновидная (фузиформная) извилина

Система исполнительного контроля Фронто-париетальная система отвечает за выбор цели, а цингуло-оперкулярная – за удержание цели, включая проверку и разрешение конфликтов. Two networks of the executive control system. The circled region indicates the original member of the executive control system from Posner & Petersen (1990). The remaining regions come from the elaboration of the original cingulo-opercular system (black) and the addition of the frontoparietal system (yellow) (adapted from Dosenbach et al. 2007) (Petersen, Posner 2012).

430

431
 и вентральная (восходящий контроль). Вентральная система асимметрична и имеет")
Система ориентирования: дорсальная (нисходящий контроль) и вентральная (восходящий контроль). Вентральная система асимметрична и имеет правую латерализацию. The dorsal and ventral orienting networks (after Corbetta & Shulman 2002). The dorsal attention network (light green) consists of frontal eye fields (FEF) and the intraparietal sulcus/superior parietal lobe (IPS/SPL). The ventral attention network (teal ) consists of regions in the temporoparietal junction (TPJ) and the ventral frontal cortex (VFC). (Petersen, Posner 2012).

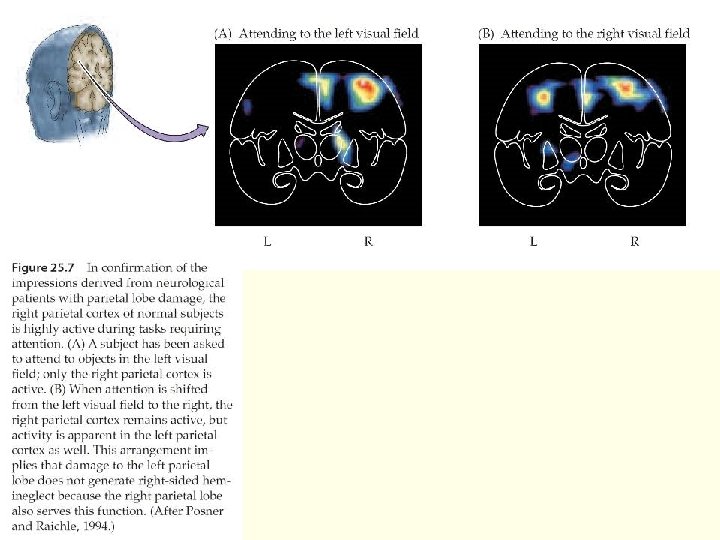
Система ориентирования: Дорсальная система Human brain activity produced by attending to a location. Subjects see an arrow that cues one of two locations and covertly attend to the location indicated in preparation for a target at that location. The functional magnetic resonance imaging (f. MRI) response is averaged over 13 subjects. The graphs show the time course of the f. MRI signal after the cue. Signals are transient in occipital regions, but sustained (grey arrow) in the parietal cortex and frontal eye field (FEF). a. IPs, anterior intraparietal sulcus; Fus, fusiform cortex; MT+, middle temporal complex; p. IPs, posterior IPs; v. IPs, ventral IPs (junction of the v. IPs and transverse occipital sulcus).

Система ориентирования: Дорсальная система Human brain activity produced by attending to a direction of motion. Subjects see an arrow that cues a direction of motion and prepare for a subsequent target moving in that direction. The response is averaged over 14 subjects. The graphs contrast the response seen when the arrow cues a direction of motion with that observed when a cue instructs the subject to view the display passively. Directional cues produce sustained signals in frontal (FEF) and parietal (a. IPs, p. IPs) areas, but transient signals in occipital areas (MT+).

Система ориентирования: Вентральная система Spatial reorienting to unattended targets. Maps of activation showing areas of differential activation for infrequent unattended spatial targets (invalid) versus frequent attended (valid) targets. The time course indicates that the response is stronger and more sustained to unattended targets than to attended targets. FEF, frontal eye field; IFg, inferior frontal gyrus; IPs, intraparietal sulcus; TPJ, temporoparietal junction.

Кортикальные области, пораженные у больных с односторонним пространственным игнорированием
")
1452 Рисунок больного с синдромом одностороннего игнорирования (по Лурия, 1973)

1452 Игнорирование левой стороны при воспроизведении изображения у больных с односторонней оптической агнозией {синдромом одностороннего игнорирования}. а и б — Больной M — опухоль правой теменно-затылочной области. В рисунках больного следует отметить игнорирование левой стороны изображения. (по А. Р. Лурия, 1969)

1455

1451

1451

Премоторная теория Дж. Риццолатти

Giacomo Rizzolatti University of Parma

Премоторная теория Дж. Риццолатти: В отличие от большинства других авторов, Дж. Риццолатти настаивает, что система внимания неотделима от системы премоторного контроля (управления движением)
 Vertical meridian effect. Schematic drawing of the experimental")
Эффект вертикального меридиана (перемещение скрытого внимания) Vertical meridian effect. Schematic drawing of the experimental set up used by Rizzolatti, Riggio, Dascola, Umiltà (1987) to test the presence of the Vertical meridian effect. The visual display comprised one fixation box and four boxes for stimulus presentation. The stimulus boxes were marked by an adjacent digit (1 -4). The fixation box was always shown at the geometrical centre of the screen, whereas the position of the other boxes varied according to different blocks (horizontally in the upper or lower hemifield, vertically in the right or left hemifield). The cue used for directing attention was a digit (1 -4), presented in the fixation box and indicating a probability of 80% for the target to appear in the relative box (valid trial) and 20% in one of the other possible boxes (invalid trials). Participants had to respond to target appearance by pressing a key with their right index finger. The importance of maintaining fixation was stressed and eye movements were monitored. The presence of the Vertical meridian effect was specifically tested by comparing reaction times to stimulus presentation in two locations that were at the same distance from the cued box (e. g. , box 2) but in different hemifields. This comparison was significant and showed that reaction times were faster in the same (e. g. , stimulus in box 1) than in the opposite hemifield (e. g. , stimulus in box 3), indicating that passing from one hemifield to the other caused an additional delay of about 21 ms, regardless of which meridian had to be crossed.

Отклонение вертикальных саккад в сторону, противоположную направленности скрытого внимания The figure reports an example of the experimental paradigm used by Sheliga, Riggio, and Rizzolatti (1995) to support the premotor theory of attention. The basic assumption was that if visuospatial attention involves an activation of oculomotor circuits, then this activation should influence an overt oculomotor response. In contrast, if attention is not related to oculomotion, there is no reason why such an influence should occur. The basic experimental situation was similar to that used in experiments on meridian effect (see Figure 1). Participants were instructed to fixate a central cross and to pay attention to one of the possible stimulus locations according to the cue. The major difference was that the measured variable was not a manual response but a vertical saccade directed to a box located below or above the fixation point. Specifically, the visual display comprised three filled and two empty boxes. The three filled boxes were arranged horizontally. The two empty boxes were located above and below the central filled box. The central box was the fixation box. The cue was a thin line attached to this box, indicating the left box if pointing left, the right box if pointing right, the central box if pointed up. The stimulus consisted in a white vertical line across one of the filled boxes. In response to stimulus presentation, participants had to make a vertical saccade directed to one of the empty boxes, according to instructions. The figure shows all saccadic responses from one representative participant. Left side. Saccadic trajectories in response to imperative stimulus in the left box (upper panel), in the central box (middle panel), in the right box (lower panel). Right side. Mean horizontal deviation of saccades with imperative stimulus presented in the left box (upper panel) or in the right box (lower panel) after point-by-point subtraction of the mean horizontal deviation of saccades with the imperative stimulus presented in the central box (abscissa: horizontal deviation; ordinate: time). Main results showed that saccade trajectory deviated contralaterally to stimulus location, in particular the strength of the deviation was greater when trials were valid, that is when active attention was on the locus of imperative stimulus presentation. Finally, the deviation was present also when the imperative stimulus consisted not in a visual one, but in a computer-generated sound presented centrally, demonstrating that the deviation of response saccades doesn’t depend from the location of visual stimulus presentation.

Поиск нейросетевых механизмов внимания – «треугольные» сети Д. Ла. Берджа

David La. Berge Department of Cognitive Sciences University of California, Irvine

La. Berge, 2000

La. Berge, 2000

La. Berge, 2000

La. Berge, 2000

La. Berge, 2000

Частотная синхронизация как механизм связывания признаков воспринимаемого объекта и внимание
, целостный образ представлен в виде ансамбля взаимосвязанных нейронов.")
Согласно идеям Д. Хебба (Hebb, 1949), целостный образ представлен в виде ансамбля взаимосвязанных нейронов. При этом связь между синхронно активными нейронами усиливается Каким образом одни и те же нейроны могут в разное время принимать участие в разных репрезентациях? «Связывание» достигается на основе временной синхронизации активности нейронов на определенной частоте (Abeles, 1982; von der Malsburg, 1986; Engel et al. , 1992; Singer, 1993; Basar et al. , 2001; Мачинская, 2003; Singer, 2003; Klimesch et al. , 2007). Именно так, в частности, достигается нисходящая (top-down) селективная преднастройка внимания (Niebur et al. , 1993). Согласно идеям Д. Хебба (Hebb, 1949), получившим надежное экспериментальное подтверждение, .

Альфа-ритм и внимание

Wolfgang Klimesch Fachbereich Psychologie Universität Salzburg

The basic principles underlying the inhibition–timing hypothesis can be illustrated by considering the phase of oscillatory activity together with the level of excitation of single neurons. We assume that oscillatory activity is induced by inhibitory cells and reflects rhythmic changes between phases of maximal and minimal inhibition. Depending on the amplitude of the oscillation (and the excitation level of single cells), two different firing patterns can be distinguished. (A) If the amplitude of the oscillation is small, cells with a high level of excitation fire tonically, not entrained to the phase of the oscillation. (B) If the amplitude is large even cells with a high level of excitation will fire rhythmically, entrained to the phase of the oscillation. Thus, increased rhythmic activity allows for a more precise timing of neural activity.

Регина Ильинична Мачинская Института возрастной физиологии РАО

Гамма-ритм и внимание

В настоящее время распространена точка зрения, что во внимании огромную роль играет синхронизация между областями мозга на высоких частотах (в особенности на частотах гамма-ритма, 25 -70 Гц). Такая синхронизация теоретически позволяет создавать определенные каналы обмена информацией между конкретными областями коры, настроенные на определенный характер передаваемой информации – и благодаря этому обеспечить обработку одних сигналов и игнорирование других другие. Синхронизация разряда нейронов на частоте гаммаритма возможно лишь при достаточном уровне возбуждения в них, которое поддерживается медиаторами системы бодрствования, из которых, видимо, особенно важны ацетилхолин и норадреналин.

Компьютерное моделирование нервных сетей показало, что синхронизация клеток коры в диапазоне гамма-ритма, возникающая при воздействии ацетилхолина, усиливает реакцию сети на сенсорный стимул и снижает ее порог [Börgers et al. , 2005] - имитация продолжительного внимания (бдительности). Растры импульсной активности 160 возбудительных клеток коры в ответ на сенсорный стимул, адресованный к клеткам 11 -30, при подавлении токов адаптации Мхолинергическими агонистами (a); при отсутствии такого подавления реакция на тот же стимул выражена значительно слабее (b). [Börgers et al. , 2005].

… а также способствует контрастированию реакции на два стимула, действующие одновременно [Börgers et al. , 2005] - имитация селективного внимания. Растры импульсной активности 160 возбудительных клеток коры в ответ на слабый сенсорный стимул, адресованный к клеткам 11 -30, и более сильный сенсорный стимул, адресованный к клеткам 121 -140 при подавлении токов адаптации М-холинергическими агонистами (a); при отсутствии такого подавления контраст между реакциями на два стимула выражен значительно слабее (b).

Активация и внимание

Фильтры по У. Найссеру по А. Трейсман по Р. Шиффрину Длиннолатентные ВП Первичный анализ ранний предвнимание автоматическая переработка информации До 200 мс: P 1, N 1, P 2 Детальный анализ и принятие решения поздний фокальное внимание связывание (? ) контролируемая переработка информации После 200 мс: N 2, P 3, N 4

10319022ed50799bd48e489d55ef3a0f.ppt