

lektsiya_8_fotosintez_ch_1_2010-11.ppt
- Количество слайдов: 33
 Лекція 8 розділ ФОТОСИНТЕЗ
Лекція 8 розділ ФОТОСИНТЕЗ
 План лекції 1. Сутність, значення та масштаби фотосинтезу 2. Історія вивчення фотосинтезу 3. Морфологія та ультраструктура хлоропласта
План лекції 1. Сутність, значення та масштаби фотосинтезу 2. Історія вивчення фотосинтезу 3. Морфологія та ультраструктура хлоропласта
 1. Сутність, значення та масштаби фотосинтезу Фотосинтез – процес, в якому рослини виконують роль посередника між енергією сонця та всім живим на землі. Фотосинтез – асиміляція сонячної енергії, її перетворення в хімічну енергію за рахунок синтезу багатих енергією органічних сполук з неорганічних сполук – води та вуглекислого газу.
1. Сутність, значення та масштаби фотосинтезу Фотосинтез – процес, в якому рослини виконують роль посередника між енергією сонця та всім живим на землі. Фотосинтез – асиміляція сонячної енергії, її перетворення в хімічну енергію за рахунок синтезу багатих енергією органічних сполук з неорганічних сполук – води та вуглекислого газу.
 Без зелених рослин і властивого лише їм процесу фотосинтезу існування інших організмів було б неможливе. Космічна роль зелених рослин, на яку вказував засновник сучасного вчення про фотосинтез К. А. Тімірязєв.
Без зелених рослин і властивого лише їм процесу фотосинтезу існування інших організмів було б неможливе. Космічна роль зелених рослин, на яку вказував засновник сучасного вчення про фотосинтез К. А. Тімірязєв.
 В процесі фотосинтезу нагромаджуються органічні речовини, що є джерелом енергії та будівельним матеріалом для усіх гетеротрофів, зокрема і людини. Зелені рослини – єдині продуценти кисню. Вміст кисню в атмосфері сталий за рахунок процесу фотосинтезу. Акумульована в процесі фотосинтезу енергія може зберігатися дуже довго: вугілля, нафта, природний газ – продукти організмів, що існували мільйони років тому, а енергією ми користуємося зараз.
В процесі фотосинтезу нагромаджуються органічні речовини, що є джерелом енергії та будівельним матеріалом для усіх гетеротрофів, зокрема і людини. Зелені рослини – єдині продуценти кисню. Вміст кисню в атмосфері сталий за рахунок процесу фотосинтезу. Акумульована в процесі фотосинтезу енергія може зберігатися дуже довго: вугілля, нафта, природний газ – продукти організмів, що існували мільйони років тому, а енергією ми користуємося зараз.
 Масштаби фотосинтезу • За рік зелені рослини фіксують 17, 4 • 1010 т вуглецю, що становить 20 -25% від його запасів у повітрі у вигляді СО 2 та 0, 30, 4% загальної кількості вуглецю у гідросфері та тропосфері. • Загальна продукція органічної речовини, що синтезується всією рослинністю земної кулі досягає 4, 5 • 1011 т (в перерахунку на глюкозу). • Людина використовує 3, 5% органічного вуглецю, синтезованого наземною флорою та 5 • 10 -5% вуглецю, асимільованого рослинністю морів та океанів. • Одночасно рослини виділяють в атмосферу 5 • 1011 т кисню.
Масштаби фотосинтезу • За рік зелені рослини фіксують 17, 4 • 1010 т вуглецю, що становить 20 -25% від його запасів у повітрі у вигляді СО 2 та 0, 30, 4% загальної кількості вуглецю у гідросфері та тропосфері. • Загальна продукція органічної речовини, що синтезується всією рослинністю земної кулі досягає 4, 5 • 1011 т (в перерахунку на глюкозу). • Людина використовує 3, 5% органічного вуглецю, синтезованого наземною флорою та 5 • 10 -5% вуглецю, асимільованого рослинністю морів та океанів. • Одночасно рослини виділяють в атмосферу 5 • 1011 т кисню.
 На земну поверхню щороку потрапляє 5 • 1023 ккал сонячної енергії, частка рослин на землі становить 40%, отже рослини можуть засвоїти 2 • 1023 ккал. Враховуючи втрати енергії внаслідок розсіювання, відбиття тощо, а також енергетичний вихід фотосинтезу (мах 2%), то загальна кількість енергії, яка щорічно запасається в продуктах фотосинтезу, становить 5 • 1022 ккал. (Рубін Б. А. )
На земну поверхню щороку потрапляє 5 • 1023 ккал сонячної енергії, частка рослин на землі становить 40%, отже рослини можуть засвоїти 2 • 1023 ккал. Враховуючи втрати енергії внаслідок розсіювання, відбиття тощо, а також енергетичний вихід фотосинтезу (мах 2%), то загальна кількість енергії, яка щорічно запасається в продуктах фотосинтезу, становить 5 • 1022 ккал. (Рубін Б. А. )
 Значення фотосинтезу • Поповнення втрат органічних сполук, яке відбувається безперервно внаслідок життєдіяльності гетеротрофних організмів та в результаті виробничої діяльності людини. • Нагромадження величезної кількості хімічної енергії в продуктах фотосинтезу • Забезпечення сталого вмісту кисню в атмосфері, необхідного для існування більшості організмів. • Ліквідація надлишкового нагромадження вуглекислого газу в атмосфері.
Значення фотосинтезу • Поповнення втрат органічних сполук, яке відбувається безперервно внаслідок життєдіяльності гетеротрофних організмів та в результаті виробничої діяльності людини. • Нагромадження величезної кількості хімічної енергії в продуктах фотосинтезу • Забезпечення сталого вмісту кисню в атмосфері, необхідного для існування більшості організмів. • Ліквідація надлишкового нагромадження вуглекислого газу в атмосфері.
 Унікальна роль фотосинтезу – забезпечення кругообігу вуглецю: вуглекислий газ, кінцевий продукт розпаду органічної матерії, завдяки фотосинтезу стає джерелом утворення органічної речовини. Фотосинтез відіграє визначну роль в енергетиці біосфери в цілому.
Унікальна роль фотосинтезу – забезпечення кругообігу вуглецю: вуглекислий газ, кінцевий продукт розпаду органічної матерії, завдяки фотосинтезу стає джерелом утворення органічної речовини. Фотосинтез відіграє визначну роль в енергетиці біосфери в цілому.
 2. Історія вивчення фотосинтезу 1771 р. – початок експериментального відкриття фотосинтезу Джозеф Прістлі 1733 -1804 Дослід Д. Прістлі з мишами Прістлі встановив, що рослини роблять повітря придатним для горіння і дихання (виділяють кисень)
2. Історія вивчення фотосинтезу 1771 р. – початок експериментального відкриття фотосинтезу Джозеф Прістлі 1733 -1804 Дослід Д. Прістлі з мишами Прістлі встановив, що рослини роблять повітря придатним для горіння і дихання (виділяють кисень)
 У 1779 р. Інгенхауз встановив, що процес здійснюється лише на світлі Ян Інгенхауз 1730 -1799 Елодея У 1782 р. Сенеб‘є встановив, що кисень виділяють лише зелені частини рослини при наявності у повітрі СО 2 Жан Сенеб'є 1742 -1809
У 1779 р. Інгенхауз встановив, що процес здійснюється лише на світлі Ян Інгенхауз 1730 -1799 Елодея У 1782 р. Сенеб‘є встановив, що кисень виділяють лише зелені частини рослини при наявності у повітрі СО 2 Жан Сенеб'є 1742 -1809
 У 1804 р. Н. Сосюр встановив, що об'єм поглинутого на світлі СО 2 дорівнює об'єму виділеного О 2. Поглинання СО 2 – вуглецеве живлення рослин, яке здійснюється за участі води. У 1817 р. Ж. Пельтье і Ж. Каванту виділили із листків зелений пігмент і назвали його хлорофілом (від грецького «хлорос» — зелений і «філлон» — листок). Жозеф Каванту Жан Пельть'є 1795— 1877 1788 -1842
У 1804 р. Н. Сосюр встановив, що об'єм поглинутого на світлі СО 2 дорівнює об'єму виділеного О 2. Поглинання СО 2 – вуглецеве живлення рослин, яке здійснюється за участі води. У 1817 р. Ж. Пельтье і Ж. Каванту виділили із листків зелений пігмент і назвали його хлорофілом (від грецького «хлорос» — зелений і «філлон» — листок). Жозеф Каванту Жан Пельть'є 1795— 1877 1788 -1842
 У 1840 р. Бусенго склав загальне рівняння фотосинтезу: 6 СО 2+6 Н 2 О=С 6 Н 12 О 6+6 О 2 Жан Батист Буссенго 1802 -1887 У 1864 р. Ю. Сакс експериментально довів, що головний продукт синтезу на світлі у листках – крохмаль Проба Сакса Юліус Сакс 1832 -1897
У 1840 р. Бусенго склав загальне рівняння фотосинтезу: 6 СО 2+6 Н 2 О=С 6 Н 12 О 6+6 О 2 Жан Батист Буссенго 1802 -1887 У 1864 р. Ю. Сакс експериментально довів, що головний продукт синтезу на світлі у листках – крохмаль Проба Сакса Юліус Сакс 1832 -1897
 У 1869 р. засновник сучасного вчення про фотосинтез К. А. Тімірязєв експериментально обгрунтував акумуляцію і перетворення енергії зеленим листком. Довів, що для цього необхідне червоне світло. Показав, що процес фотосинтезу здійснюється за законом перетворення і збереження енергій. У 1877 р. Пфеффер ввів термін фотосинтез Вільям Пфеффер 1845 -1920 У 1893 – 98 рр А. Н. Бах показав, що фотосинтез – це серія спряжених окисно-відновних реакцій, в результаті яких відбувається асиміляція вуглекислого газу та фотохімічне відновлення кисню з води Климент Аркадійович Тімірязєв 1843 -1920 А. Н. Бах
У 1869 р. засновник сучасного вчення про фотосинтез К. А. Тімірязєв експериментально обгрунтував акумуляцію і перетворення енергії зеленим листком. Довів, що для цього необхідне червоне світло. Показав, що процес фотосинтезу здійснюється за законом перетворення і збереження енергій. У 1877 р. Пфеффер ввів термін фотосинтез Вільям Пфеффер 1845 -1920 У 1893 – 98 рр А. Н. Бах показав, що фотосинтез – це серія спряжених окисно-відновних реакцій, в результаті яких відбувається асиміляція вуглекислого газу та фотохімічне відновлення кисню з води Климент Аркадійович Тімірязєв 1843 -1920 А. Н. Бах
 У 1903 р, започаткувавши хроматографічний метод аналізу, М. С. Цвєт відкрив 2 форми хлорофілу у вищих рослин У 1905 р. Ф. Блекман встановив двофазний характер фотосинтезу. Хроматографічна колонка з 2 зонами хлорофілу У 1913 р. Р. Вільштеттер встановив хімічний склад молекули хлорофілу. Разом із учнем А. Штролем він опублікував фундаментальну працю «Дослідження хлорофілу» , за що у 1915 р. отримав Нобелівську премію з хімії Ріхард Вільштеттер 1872— 1942
У 1903 р, започаткувавши хроматографічний метод аналізу, М. С. Цвєт відкрив 2 форми хлорофілу у вищих рослин У 1905 р. Ф. Блекман встановив двофазний характер фотосинтезу. Хроматографічна колонка з 2 зонами хлорофілу У 1913 р. Р. Вільштеттер встановив хімічний склад молекули хлорофілу. Разом із учнем А. Штролем він опублікував фундаментальну працю «Дослідження хлорофілу» , за що у 1915 р. отримав Нобелівську премію з хімії Ріхард Вільштеттер 1872— 1942
 – швидкі фотохімічні і повільні біохімічні реакції") Мелвін Кальвін Р. Емерсон, А. Арнольд (1932) – швидкі фотохімічні і повільні біохімічні реакції фотосинтезу; (1942 -1957) – наявність 2 фотосистем. Р. Хілл (1937) – нециклічний потік електронів, джерело електронів – вода, акцептори – окислені сполуки. Для виділення О 2 необхідна вода. К. Ван-Ніль, А. П. Віноградов, Р. В. Тейс, С. Рубен, М. Камен (1935 -1941) – кисень у фотосинтезі утворюється внаслідок фотолізу води. М. Кальвін, Бенсон, Басем (1947 -1954) – метаболізм карбону при фотосинтезі (С 3 -цикл асиміляції вуглецю).
Мелвін Кальвін Р. Емерсон, А. Арнольд (1932) – швидкі фотохімічні і повільні біохімічні реакції фотосинтезу; (1942 -1957) – наявність 2 фотосистем. Р. Хілл (1937) – нециклічний потік електронів, джерело електронів – вода, акцептори – окислені сполуки. Для виділення О 2 необхідна вода. К. Ван-Ніль, А. П. Віноградов, Р. В. Тейс, С. Рубен, М. Камен (1935 -1941) – кисень у фотосинтезі утворюється внаслідок фотолізу води. М. Кальвін, Бенсон, Басем (1947 -1954) – метаболізм карбону при фотосинтезі (С 3 -цикл асиміляції вуглецю).
 – циклічне і нециклічне фотофосфорилювання – фотоенергетичні реакції хлоропластів. Р.") Д. Арнон (1954 -1957) – циклічне і нециклічне фотофосфорилювання – фотоенергетичні реакції хлоропластів. Р. Хілл (1961) – Z-схема фотосинтезу. М. Хеч, К. Слек, Ю. Карпілов (1957 -1964) – С 4 -тип фотосинтетичного метаболізму. О. Варбург (1920), Н. Воскресенська (1968 -1974) – фотодихання, гліоксилатний шлях фотосинтезу. Видатні науковці, які вивчали фотосинтез: Курсанов А. Л. , Ничипорович А. А. , Рубін Б. А. , Гродзинський Д. М. , Оканенко А. С. , Годнєв Т. Н. , Красновський А. В.
Д. Арнон (1954 -1957) – циклічне і нециклічне фотофосфорилювання – фотоенергетичні реакції хлоропластів. Р. Хілл (1961) – Z-схема фотосинтезу. М. Хеч, К. Слек, Ю. Карпілов (1957 -1964) – С 4 -тип фотосинтетичного метаболізму. О. Варбург (1920), Н. Воскресенська (1968 -1974) – фотодихання, гліоксилатний шлях фотосинтезу. Видатні науковці, які вивчали фотосинтез: Курсанов А. Л. , Ничипорович А. А. , Рубін Б. А. , Гродзинський Д. М. , Оканенко А. С. , Годнєв Т. Н. , Красновський А. В.
 Сучасний етап вивчення фотосинтезу: • Розшифрована генетична система хлоропластів, відомі гени, що відповідають за фотосинтетичні процеси. • Вивчена структура фотосистем I та II. • Сформульовані уявлення про механізми виділення кисню (фотоліз води). • Визначена роль фотосинтезу в донорно-акцепторних відносинах рослини. • Активно вивчаються екологічні аспекти фотосинтезу. • Розробляються прикладні дослідження фотосинтезу.
Сучасний етап вивчення фотосинтезу: • Розшифрована генетична система хлоропластів, відомі гени, що відповідають за фотосинтетичні процеси. • Вивчена структура фотосистем I та II. • Сформульовані уявлення про механізми виділення кисню (фотоліз води). • Визначена роль фотосинтезу в донорно-акцепторних відносинах рослини. • Активно вивчаються екологічні аспекти фотосинтезу. • Розробляються прикладні дослідження фотосинтезу.
 3. Морфологія та ультраструктура хлоропласта Основні функції листка • Орган автотрофного живлення • Біосинтез органічних речовин (в т. ч ауксинів, гіберелінів, АБК) • Транспірація
3. Морфологія та ультраструктура хлоропласта Основні функції листка • Орган автотрофного живлення • Біосинтез органічних речовин (в т. ч ауксинів, гіберелінів, АБК) • Транспірація
: стовпчастий та губчастий Стовпчастий мезофіл") Основна тканина листка, де відбувається фотосинтез – мезофіл (паренхіма): стовпчастий та губчастий Стовпчастий мезофіл Губчастий мезофіл
Основна тканина листка, де відбувається фотосинтез – мезофіл (паренхіма): стовпчастий та губчастий Стовпчастий мезофіл Губчастий мезофіл
 Морфологія хлоропластів • Хлоропласти вищої рослини – диски, плоскі еліпсоїди або лінзоподібні тіла довжиною 4 -10 мкм та товщиною 2 -5 мкм. • Кількість хлоропластів у клітині – від 1 до 100 шт. Число хлоропластів в клітині не є постійним, залежить від виду та віку рослини, рівня мінерального живлення, водозабезпечення тощо. Так, в ході онтогенезу кількість хлоропластів у клітині мезофілу кукурудзи зростає з 8 до 14, а розмір – з 4 до 8 мкм. Під впливом мінерального живлення збільшуються розміри клітин мезофілу листків цукрового буряка і число хлоропластів у них. Фон живлення Розмір клітин, мкм Кількість хлоропластів, шт PK 46 • 31 102 NPK 57 • 38 241 N 2 PK 62 • 33 270
Морфологія хлоропластів • Хлоропласти вищої рослини – диски, плоскі еліпсоїди або лінзоподібні тіла довжиною 4 -10 мкм та товщиною 2 -5 мкм. • Кількість хлоропластів у клітині – від 1 до 100 шт. Число хлоропластів в клітині не є постійним, залежить від виду та віку рослини, рівня мінерального живлення, водозабезпечення тощо. Так, в ході онтогенезу кількість хлоропластів у клітині мезофілу кукурудзи зростає з 8 до 14, а розмір – з 4 до 8 мкм. Під впливом мінерального живлення збільшуються розміри клітин мезофілу листків цукрового буряка і число хлоропластів у них. Фон живлення Розмір клітин, мкм Кількість хлоропластів, шт PK 46 • 31 102 NPK 57 • 38 241 N 2 PK 62 • 33 270
 Будова хлоропласта • Оболонка, складається з внутрішньої і зовнішньої мембрани, виконує ізолюючу функцію, бере участь в іонному обміні • Система фотосинтетичних мембран – ламел (тилакоїди, грани), де зосереджені фотохімічні та фотофізичні процеси (світлова фаза фотосинтезу) • Строма – матрикс хлоропласта, де зосереджені біохімічні процеси (темнова фаза фотосинтезу).
Будова хлоропласта • Оболонка, складається з внутрішньої і зовнішньої мембрани, виконує ізолюючу функцію, бере участь в іонному обміні • Система фотосинтетичних мембран – ламел (тилакоїди, грани), де зосереджені фотохімічні та фотофізичні процеси (світлова фаза фотосинтезу) • Строма – матрикс хлоропласта, де зосереджені біохімічні процеси (темнова фаза фотосинтезу).
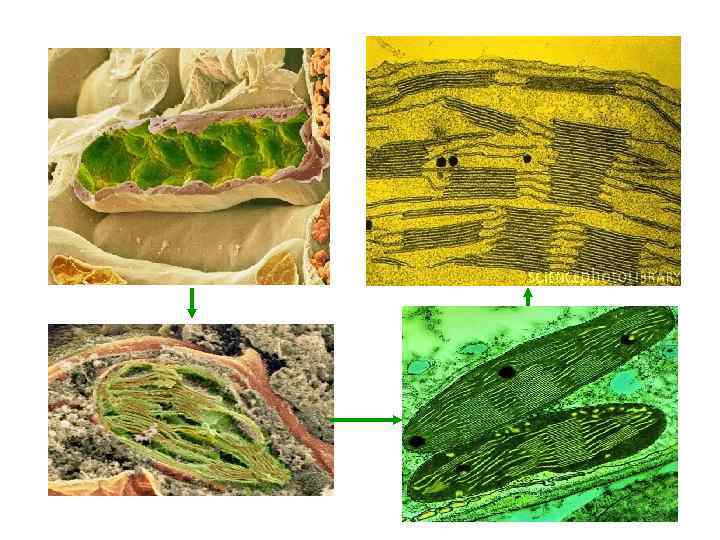
 Будова хлоропласту зовнішня мембрана внутрішня мембрана тилакоїди грана строма
Будова хлоропласту зовнішня мембрана внутрішня мембрана тилакоїди грана строма
 Міжмембранний простір Зовнішня мембрана тилакоїд Ламели грани нашарування") Ультраструктура хлоропласта Ламели строми, (місцерозміщення ФСІ) Міжмембранний простір Зовнішня мембрана тилакоїд Ламели грани нашарування тилакоїдів, місцерозміщення ФСІІ тилакоїд строма Внутрішня мембрана Люмен тилакоїда Товщина ламели строми – 20 -30 А 0 ламели грани – 40 -60 А 0 Грана, нашарування тилакоїдів Ламела строми
Ультраструктура хлоропласта Ламели строми, (місцерозміщення ФСІ) Міжмембранний простір Зовнішня мембрана тилакоїд Ламели грани нашарування тилакоїдів, місцерозміщення ФСІІ тилакоїд строма Внутрішня мембрана Люмен тилакоїда Товщина ламели строми – 20 -30 А 0 ламели грани – 40 -60 А 0 Грана, нашарування тилакоїдів Ламела строми
 Фотосинтетична мембрана Тилакоїдна мембрана має зернисту структуру, вона містить функціональні комплекси, в яких локалізовані механізми перетворення енергії – квантосоми (спряжений фактор)
Фотосинтетична мембрана Тилакоїдна мембрана має зернисту структуру, вона містить функціональні комплекси, в яких локалізовані механізми перетворення енергії – квантосоми (спряжений фактор)
 Будова фотосинтетичної мембрани
Будова фотосинтетичної мембрани
 Молекулярна структура тилакоїдної мембрани Тилакоїдна мембрана строма люмен 1 2 3 4 5 1 – світлозбиральний комплекс 2 – фотосистема ІІ 3 – цитохром b 6 f 4 – фотосистема І 5 – АТФ-синтаза
Молекулярна структура тилакоїдної мембрани Тилакоїдна мембрана строма люмен 1 2 3 4 5 1 – світлозбиральний комплекс 2 – фотосистема ІІ 3 – цитохром b 6 f 4 – фотосистема І 5 – АТФ-синтаза
 Строма – гідрофільний, гомогенний біоколоїд хлоропласта, де здійснюються реакції фотосинтетичної асиміляції вуглецю. Строма містить: • кільцеву ДНК, • всі види РНК • рибосоми, • пластоглобули • рибулозо-бісфосфаткарбоксилазу-оксидазу (RUBISCO) – ключовий фермент вуглеводного обміну фотосинтезу, інші ферменти вуглеводного циклу фотосинтезу • ферменти синтезу фотосинтетичних пігментів • ферменти синтезу полярних ліпідів мембран хлоропластів • ферменти синтезу білків мультипептидних комплексів мембран тилакоїдів • водорозчинні органічні сполуки та неорганічні іони.
Строма – гідрофільний, гомогенний біоколоїд хлоропласта, де здійснюються реакції фотосинтетичної асиміляції вуглецю. Строма містить: • кільцеву ДНК, • всі види РНК • рибосоми, • пластоглобули • рибулозо-бісфосфаткарбоксилазу-оксидазу (RUBISCO) – ключовий фермент вуглеводного обміну фотосинтезу, інші ферменти вуглеводного циклу фотосинтезу • ферменти синтезу фотосинтетичних пігментів • ферменти синтезу полярних ліпідів мембран хлоропластів • ферменти синтезу білків мультипептидних комплексів мембран тилакоїдів • водорозчинні органічні сполуки та неорганічні іони.
 Положення хлоропластів у клітині Хлоропласти в клітині переміщуються пасивно разом із цитоплазмою, та активно – реагуючи на інтенсивність освітлення, дію фізіологічно активних речовин тощо. Сильне освітлення викликає негативний фототаксис хлоропластів, слабке – позитивний. Активний рух хлоропластів – енергозатратний процес, в якому використовується АТФ, він пов'язаний з дією скоротливих білків на їх поверхні та у цитоплазмі. Клітини у темряві Слабке освітлення Сильне освітлення
Положення хлоропластів у клітині Хлоропласти в клітині переміщуються пасивно разом із цитоплазмою, та активно – реагуючи на інтенсивність освітлення, дію фізіологічно активних речовин тощо. Сильне освітлення викликає негативний фототаксис хлоропластів, слабке – позитивний. Активний рух хлоропластів – енергозатратний процес, в якому використовується АТФ, він пов'язаний з дією скоротливих білків на їх поверхні та у цитоплазмі. Клітини у темряві Слабке освітлення Сильне освітлення
 3 -мірна модель організації внутрішніх мембран хлоропластів
3 -мірна модель організації внутрішніх мембран хлоропластів
 Онтогенез хлоропластів Ініціальна частинка строма Інвагінації внутрішньої мембрани Утворення внутрішньої ламелярної системи Формування хлоропласта може відбуватися двома шляхами: • Безпосереднє перетворення пропластид в хлоропласти 1 мкм пропластида Крохмальне зерно Світло Темрява 2 мкм 3 мкм грана Ламела строми Пухирці, утворені інвагінаціями внутрішньої мембрани Проламелярне тіло Світло Утворення ламел 4 мкм • Утворення хлоропластів з етіопластів 5 мкм Повністю сформований хлоропласт Крапля жиру
Онтогенез хлоропластів Ініціальна частинка строма Інвагінації внутрішньої мембрани Утворення внутрішньої ламелярної системи Формування хлоропласта може відбуватися двома шляхами: • Безпосереднє перетворення пропластид в хлоропласти 1 мкм пропластида Крохмальне зерно Світло Темрява 2 мкм 3 мкм грана Ламела строми Пухирці, утворені інвагінаціями внутрішньої мембрани Проламелярне тіло Світло Утворення ламел 4 мкм • Утворення хлоропластів з етіопластів 5 мкм Повністю сформований хлоропласт Крапля жиру
 Хлоропласти – напівавтономні органели • Містять власну ДНК • Діляться незалежно від ядра • Містять власну білок-синтезувальну систему
Хлоропласти – напівавтономні органели • Містять власну ДНК • Діляться незалежно від ядра • Містять власну білок-синтезувальну систему