

Лек 8.2014.ppt
- Количество слайдов: 46
 Лекция 8 Пероксисомы Митохондрии Хлоропласты
Лекция 8 Пероксисомы Митохондрии Хлоропласты
 Пероксисомы Имеются во всех эукариотических клетках. Диаметр 0, 3 - 1, 5 мкм, лграничены одной мембраной. Плотный кристаллический центр – уратоксидаза
Пероксисомы Имеются во всех эукариотических клетках. Диаметр 0, 3 - 1, 5 мкм, лграничены одной мембраной. Плотный кристаллический центр – уратоксидаза
 Пероксисомы Впервые пероксисомы идентифицировали как органоиды, вырабатывающие перекись водорода. Пероксисомы содержат различные оксидазы, включая ацил-Ко. Аоксидазу, оксидазы гликолата, урата, алкоголя и др. Оксидазы окисляют субстрат и восстанавливают кислород до перекиси водорода. Каталаза восстанавливает перекись водорода до воды, её большое количество содержится в пероксисомах. Задача пероксисом – «убрать» перекисные соединения, которые повреждают и разрушают клеточные структуры. Пероксисомы осуществляют биологическое окисление, но без синтеза АТФ и НАДН. Все пероксисомы содержат ферменты, разрушающие жирные кислоты. Пероксисомы вовлечены в синтез желчных кислот и холестрина, в метаболизм аминокислот и пуринов. Пероксисомы – «катаболические» органоиды.
Пероксисомы Впервые пероксисомы идентифицировали как органоиды, вырабатывающие перекись водорода. Пероксисомы содержат различные оксидазы, включая ацил-Ко. Аоксидазу, оксидазы гликолата, урата, алкоголя и др. Оксидазы окисляют субстрат и восстанавливают кислород до перекиси водорода. Каталаза восстанавливает перекись водорода до воды, её большое количество содержится в пероксисомах. Задача пероксисом – «убрать» перекисные соединения, которые повреждают и разрушают клеточные структуры. Пероксисомы осуществляют биологическое окисление, но без синтеза АТФ и НАДН. Все пероксисомы содержат ферменты, разрушающие жирные кислоты. Пероксисомы вовлечены в синтез желчных кислот и холестрина, в метаболизм аминокислот и пуринов. Пероксисомы – «катаболические» органоиды.
 Новые пероксисомы образуются: 1. Путем деления 2. При участии цистерн ЭПР
Новые пероксисомы образуются: 1. Путем деления 2. При участии цистерн ЭПР
 Деление пероксисом в клетках печени крысы
Деление пероксисом в клетках печени крысы
 Р Р Электронная томография Связь ЭПР и ламелл Электронная томография Связь пероксисомы с ламеллой
Р Р Электронная томография Связь ЭПР и ламелл Электронная томография Связь пероксисомы с ламеллой
 L") Связь пероксисом, мембранных ламелл и ЭПР Р Р P – пероксисомы (зеленый цвет) L – ламеллы (светло-голубой) ER- ЭПР (фиолетовый) Рибосомы – красный
Связь пероксисом, мембранных ламелл и ЭПР Р Р P – пероксисомы (зеленый цвет) L – ламеллы (светло-голубой) ER- ЭПР (фиолетовый) Рибосомы – красный
 Пероксисомы в одной клетке различаются между собой по набору и количеству ферментов, различаются они также в разных клетках. Функции пероксисом различаются у разных видов, однако общим является наличие ферментов, разрушающих жирные кислоты. Белки пероксисом – пероксины. Синтезируются на свободных полисомах и попадают в цитозоль, где после формирования третичной структуры они распознаются рецепторами цитозоля и направляются в пероксисомы. Пероксисомы не участвуют в пузырьковом транспорте. Пероксины ответственны за транспорт веществ в матрикс пероксисом. В растительных клетках пероксисомы выполняют целый ряд функций, связанных с фотодыханием и метаболизмом жирных кислот. Пероксисомы функционально тесно связаны с митохондриями в метаболизме жирных кислот.
Пероксисомы в одной клетке различаются между собой по набору и количеству ферментов, различаются они также в разных клетках. Функции пероксисом различаются у разных видов, однако общим является наличие ферментов, разрушающих жирные кислоты. Белки пероксисом – пероксины. Синтезируются на свободных полисомах и попадают в цитозоль, где после формирования третичной структуры они распознаются рецепторами цитозоля и направляются в пероксисомы. Пероксисомы не участвуют в пузырьковом транспорте. Пероксины ответственны за транспорт веществ в матрикс пероксисом. В растительных клетках пероксисомы выполняют целый ряд функций, связанных с фотодыханием и метаболизмом жирных кислот. Пероксисомы функционально тесно связаны с митохондриями в метаболизме жирных кислот.
 В настоящее время появляются все новые данные о важной роли пероксисом в старении отдельной клетки. Показано, что «молодые» клетки успешно проводят детоксикацию, «правильно» окисляют жирные кислоты и справляются с окислительным стрессом, для чего необходима координированная и «правильная» работа пероксисом и митохондрий. В «старой» клетке эти механизмы нарушены, происходит её повреждение. В 2011 году было показано, что старение единичной клетки и старческие изменения целого организма связаны. Это ставит пероксисомы в разряд «критических» структур, определяющих развитие различных «возрастных» заболеваний. Задача новых исследований – понять, как пероксисомы взаимодействуют с мембранными органоидами, вовлеченными в процесс старения клетки.
В настоящее время появляются все новые данные о важной роли пероксисом в старении отдельной клетки. Показано, что «молодые» клетки успешно проводят детоксикацию, «правильно» окисляют жирные кислоты и справляются с окислительным стрессом, для чего необходима координированная и «правильная» работа пероксисом и митохондрий. В «старой» клетке эти механизмы нарушены, происходит её повреждение. В 2011 году было показано, что старение единичной клетки и старческие изменения целого организма связаны. Это ставит пероксисомы в разряд «критических» структур, определяющих развитие различных «возрастных» заболеваний. Задача новых исследований – понять, как пероксисомы взаимодействуют с мембранными органоидами, вовлеченными в процесс старения клетки.
 Два главных механизма выработки энергии в клетке: 1. Митохондрии – обеспечивают выработку энергии за счет расщепления жиров и углеводов являются главными поставщиками энергии в клетке. 2. Хлоропласты – утилизируют энергию солнечного света, и запасают ее в виде АТФ и углеводов, синтезируемых из СО 2 и воды. Хлоропласты - гаранты всей жизни на Земле
Два главных механизма выработки энергии в клетке: 1. Митохондрии – обеспечивают выработку энергии за счет расщепления жиров и углеводов являются главными поставщиками энергии в клетке. 2. Хлоропласты – утилизируют энергию солнечного света, и запасают ее в виде АТФ и углеводов, синтезируемых из СО 2 и воды. Хлоропласты - гаранты всей жизни на Земле
 МИТОХОНДРИИ наружная мембрана внутренняя мембрана кристы матрикс межкристное пространство межмембранное пространство
МИТОХОНДРИИ наружная мембрана внутренняя мембрана кристы матрикс межкристное пространство межмембранное пространство
 Mitochondrion -от греч. «Mitos» - нить + «Chondrion» – зернышко Размеры митохондрий - 0, 5 х 1 - 10 мкм В клетке может быть до 2 000 митохондрий В клетках печени митохондрии составляют более 20% общего объема цитоплазмы и содержат около 30— 35% общего количества белка Хондриом – совокупность всех митохондрий в клетке а - разрозненные митохондрии, б - группа митохондрий в клетках сетчатки, а б в в - митохондриальный ретикулум одноклеточной водоросли хлореллы
Mitochondrion -от греч. «Mitos» - нить + «Chondrion» – зернышко Размеры митохондрий - 0, 5 х 1 - 10 мкм В клетке может быть до 2 000 митохондрий В клетках печени митохондрии составляют более 20% общего объема цитоплазмы и содержат около 30— 35% общего количества белка Хондриом – совокупность всех митохондрий в клетке а - разрозненные митохондрии, б - группа митохондрий в клетках сетчатки, а б в в - митохондриальный ретикулум одноклеточной водоросли хлореллы
 А Б В Митохондриальный ретикулум в поперечнополосатой скелетной мышце: А - на продольном сечении мышечного волокна видны мелкие сечения митохондрий; Б - на поперечном сечении митохондрии (М) имеют вид разветвленных сложных сетей; МФ – миофибриллы В - митохондриальная система кардиомиоцитов
А Б В Митохондриальный ретикулум в поперечнополосатой скелетной мышце: А - на продольном сечении мышечного волокна видны мелкие сечения митохондрий; Б - на поперечном сечении митохондрии (М) имеют вид разветвленных сложных сетей; МФ – миофибриллы В - митохондриальная система кардиомиоцитов
 L – линейная форма;") Разные формы митохондрий. Высоковольтная электронная микроскопия целой клетки (не срез!) L – линейная форма; Br – разветвленная, C – циркулярная, B – шарообразная
Разные формы митохондрий. Высоковольтная электронная микроскопия целой клетки (не срез!) L – линейная форма; Br – разветвленная, C – циркулярная, B – шарообразная
 Схема строения митохондрии рибосомы
Схема строения митохондрии рибосомы
 Трехмерная модель митохондрии гепатоцита крысы Криоэлектронная томография Диаметр митохондрии – около 700 нм
Трехмерная модель митохондрии гепатоцита крысы Криоэлектронная томография Диаметр митохондрии – около 700 нм
 Белки митохондрий в основном синтезируются на свободных рибосомах и для их доставки не используется пузырьковый транспорт. Белки митохондрий не проходят обработку в аппарате Гольджи Липидный состав наружной мембраны аналогичен другим мембранам клетки. Липидный состав внутренней мембраны отличается и схож с бактериальным. Внутренняя мембрана митохондрий имеет высокое содержание белков (75%)
Белки митохондрий в основном синтезируются на свободных рибосомах и для их доставки не используется пузырьковый транспорт. Белки митохондрий не проходят обработку в аппарате Гольджи Липидный состав наружной мембраны аналогичен другим мембранам клетки. Липидный состав внутренней мембраны отличается и схож с бактериальным. Внутренняя мембрана митохондрий имеет высокое содержание белков (75%)
 Порины внешней мембраны митохондрий обеспечивают перенос веществ с молекулярной массой до 10 к. Да. Внутренняя мембрана митохондрий проницаема лишь для О 2, СО 2, Н 20. Внутренняя мембрана удерживает не только продукты промежуточного метаболизма (например, пируват и ацетил-Ко. А), но и неорганические ионы (Н+ и Na+). Поэтому в цитоплазме и митохондриях существуют независимые пулы ионов и метаболитов.
Порины внешней мембраны митохондрий обеспечивают перенос веществ с молекулярной массой до 10 к. Да. Внутренняя мембрана митохондрий проницаема лишь для О 2, СО 2, Н 20. Внутренняя мембрана удерживает не только продукты промежуточного метаболизма (например, пируват и ацетил-Ко. А), но и неорганические ионы (Н+ и Na+). Поэтому в цитоплазме и митохондриях существуют независимые пулы ионов и метаболитов.
 цитозоль наружная мембрана внутренняя мембрана Транспорт белков в митохондрии. матрикс Фосфолипиды митохондрий также доставляются из цитозоля.
цитозоль наружная мембрана внутренняя мембрана Транспорт белков в митохондрии. матрикс Фосфолипиды митохондрий также доставляются из цитозоля.
 Митохондрии имеют собственный геном, который транскрибируется и транслируется внутри органоидов. Белки, обеспечивающие синтез митохондриальной ДНК, кодируются ядерной ДНК. Митохондрии имеют автономную систему белкового синтеза, молекулы ДНК, не связаны с гистонами. На ДНК митохондрий происходит синтез молекул РНК разных типов: информационных, транспортных и рибосомальных. Рибосомы митохондрий локализованы в матриксе и отличаются от рибосом цитоплазмы. Рибосомы митохондрий обеспечивают синтез ряда митохондриальных белков, не кодируемых ядром. Собственная система синтеза обеспечивает синтез не более 5% от всех белков митохондрий. Основная часть белков, составляющих структурные и функциональные компоненты митохондрий, кодируется геномом ядра и синтезируется на рибосомах цитоплазмы.
Митохондрии имеют собственный геном, который транскрибируется и транслируется внутри органоидов. Белки, обеспечивающие синтез митохондриальной ДНК, кодируются ядерной ДНК. Митохондрии имеют автономную систему белкового синтеза, молекулы ДНК, не связаны с гистонами. На ДНК митохондрий происходит синтез молекул РНК разных типов: информационных, транспортных и рибосомальных. Рибосомы митохондрий локализованы в матриксе и отличаются от рибосом цитоплазмы. Рибосомы митохондрий обеспечивают синтез ряда митохондриальных белков, не кодируемых ядром. Собственная система синтеза обеспечивает синтез не более 5% от всех белков митохондрий. Основная часть белков, составляющих структурные и функциональные компоненты митохондрий, кодируется геномом ядра и синтезируется на рибосомах цитоплазмы.
 Митохондрии млекопитающих тесно связаны с пероксисомами в катаболизме жирных кислот. Многие длинноцепочечные жирные кислоты не могут быть разрушены в митохондриях, и транспортируются в пероксисомы. И наоборот, жирные кислоты с прямой цепью не разрушаются в пероксисомах, и направляются в митохондрии. В пузырьках? ? Существует поток жирных кислот между митохондриями и пероксисомами. Есть регулируемое взаимодействие митохондрий и эндосом, обеспечивающее прямую передачу ионов железа Есть прямое взаимодействие митохондрий и ЭПР посредством трансмембранных белков. Митохондрии и везикулярный транспорт? ?
Митохондрии млекопитающих тесно связаны с пероксисомами в катаболизме жирных кислот. Многие длинноцепочечные жирные кислоты не могут быть разрушены в митохондриях, и транспортируются в пероксисомы. И наоборот, жирные кислоты с прямой цепью не разрушаются в пероксисомах, и направляются в митохондрии. В пузырьках? ? Существует поток жирных кислот между митохондриями и пероксисомами. Есть регулируемое взаимодействие митохондрий и эндосом, обеспечивающее прямую передачу ионов железа Есть прямое взаимодействие митохондрий и ЭПР посредством трансмембранных белков. Митохондрии и везикулярный транспорт? ?
 Почкование везикул на поверхности бактерий и митохондрий. Негативное контрастирование целых бактериальных клеток. Разные везикулы на поверхности бактерий (A) Burkholderia cepacia C 5424 и (B) Serratia marcescens ATCC 13880. (C) Пузырек на поверхности митохондрии клетки He. La, экспрессирующей белок MAPL+желтый Флуоресцентный белок. (D) – флуоресцентная микроскопия клетки He. La, красным показан пузырек с MAPL-YFP. (E) – две митохондрии на поперечном срезе папиллярной мышцы сердца кошки. Пузырьки имеют диаметр 70 -100 нм
Почкование везикул на поверхности бактерий и митохондрий. Негативное контрастирование целых бактериальных клеток. Разные везикулы на поверхности бактерий (A) Burkholderia cepacia C 5424 и (B) Serratia marcescens ATCC 13880. (C) Пузырек на поверхности митохондрии клетки He. La, экспрессирующей белок MAPL+желтый Флуоресцентный белок. (D) – флуоресцентная микроскопия клетки He. La, красным показан пузырек с MAPL-YFP. (E) – две митохондрии на поперечном срезе папиллярной мышцы сердца кошки. Пузырьки имеют диаметр 70 -100 нм
 Бактериальные везикулы выполняют множество функций, передавая сигналы внутри колонии. Обсуждается возможная передача информации между бактериями и растениями, бактериями и животными. Везикулы митохондрий обнаружены недавно. Показано, что везикулы, содержащие белок MAPL (митохондриальная лигаза), адресованы в пероксисомы и внутри переносят материал из митохондрий. Другие везикулы не содержат MAPL, их функции не установлены.
Бактериальные везикулы выполняют множество функций, передавая сигналы внутри колонии. Обсуждается возможная передача информации между бактериями и растениями, бактериями и животными. Везикулы митохондрий обнаружены недавно. Показано, что везикулы, содержащие белок MAPL (митохондриальная лигаза), адресованы в пероксисомы и внутри переносят материал из митохондрий. Другие везикулы не содержат MAPL, их функции не установлены.
 Доказательства эндосимбиотического происхождения митохондрий:") Митохондрии происходят из эндосимбиотических бактерий (Р. Альтман, 1890 г. ) Доказательства эндосимбиотического происхождения митохондрий: - совпадение размеров и формы; - кольцевая форма молекул ДНК, нет гистонов; - нуклеотидные последовательности р- и т-РНК митохондрий отличаются от ядерных, и схожи с аналогичными РНК некоторых аэробных ГР- бактерий; - белковый синтез в митохондриях и бактериях подавляется одними и теми же антибиотиками, не влияющими на синтез белка у эукариот; - липидный состав внутренней мембраны митохондрий и бактериальной плазмалеммы сходен, и существенно отличается от такового наружной мембраны митохондрий, гомологичной другим мембранам эукариотических клеток
Митохондрии происходят из эндосимбиотических бактерий (Р. Альтман, 1890 г. ) Доказательства эндосимбиотического происхождения митохондрий: - совпадение размеров и формы; - кольцевая форма молекул ДНК, нет гистонов; - нуклеотидные последовательности р- и т-РНК митохондрий отличаются от ядерных, и схожи с аналогичными РНК некоторых аэробных ГР- бактерий; - белковый синтез в митохондриях и бактериях подавляется одними и теми же антибиотиками, не влияющими на синтез белка у эукариот; - липидный состав внутренней мембраны митохондрий и бактериальной плазмалеммы сходен, и существенно отличается от такового наружной мембраны митохондрий, гомологичной другим мембранам эукариотических клеток
 Доказательства эндосимбиотического происхождения митохондрий: - кристы, образуемые внутренней митохондриальной мембраной, являются эволюционными аналогами мезосомных мембран многих прокариот; - до сих пор сохранились организмы, имитирующие промежуточные формы на пути к образованию митохондрий из бактерий (примитивная амеба Pelomyxa не имеет митохондрий, но всегда содержит эндосимбиотические бактерии)
Доказательства эндосимбиотического происхождения митохондрий: - кристы, образуемые внутренней митохондриальной мембраной, являются эволюционными аналогами мезосомных мембран многих прокариот; - до сих пор сохранились организмы, имитирующие промежуточные формы на пути к образованию митохондрий из бактерий (примитивная амеба Pelomyxa не имеет митохондрий, но всегда содержит эндосимбиотические бактерии)
 Окисление глюкозы и жирных кислот – главный источник энергии в клетке глюкоза пируват пир. Жирные кислоты Ац. Ко. А ж. к. ЦТК цитоплазма При расщеплении глюкозы путем гликолиза: на одну молекулу глюкозы продуцируется 4 молекулы АТФ. В цикле Кребса в митохондриях на одну молекулу глюкозы продуцируется 24 -28 молекул АТФ.
Окисление глюкозы и жирных кислот – главный источник энергии в клетке глюкоза пируват пир. Жирные кислоты Ац. Ко. А ж. к. ЦТК цитоплазма При расщеплении глюкозы путем гликолиза: на одну молекулу глюкозы продуцируется 4 молекулы АТФ. В цикле Кребса в митохондриях на одну молекулу глюкозы продуцируется 24 -28 молекул АТФ.
 Процесс окислительного фосфорилирования локализован на внутренней мембране митохондрий, и поставляет основную часть энергии в клетке. «Окислительное фосфорилирование» отъем электрона от одной молекулы, и присоединение фосфатной группы к другой, с формированием макроэргической связи. При этом происходит перенос электронов с высокой энергией на молекулярный кислород посредством молекул-переносчиков электронов. Энергия, выделяющаяся в реакциях переноса электронов, превращается в потенциальную энергию, которая запасается в виде градиента протонов в пространстве между внутренней и наружной мембранами митохондрий и затем используется для синтеза АТФ.
Процесс окислительного фосфорилирования локализован на внутренней мембране митохондрий, и поставляет основную часть энергии в клетке. «Окислительное фосфорилирование» отъем электрона от одной молекулы, и присоединение фосфатной группы к другой, с формированием макроэргической связи. При этом происходит перенос электронов с высокой энергией на молекулярный кислород посредством молекул-переносчиков электронов. Энергия, выделяющаяся в реакциях переноса электронов, превращается в потенциальную энергию, которая запасается в виде градиента протонов в пространстве между внутренней и наружной мембранами митохондрий и затем используется для синтеза АТФ.
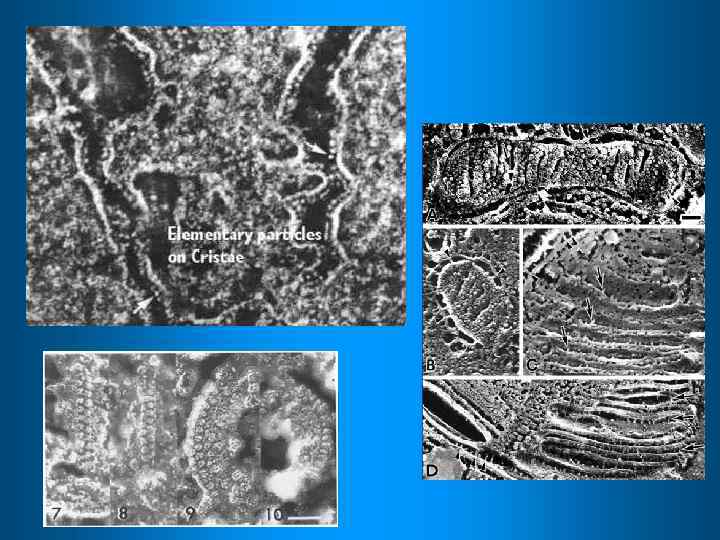
 На каждую переносимую пару электронов выделяется 52, 5 ккал/моль Цепь последовательного переноса электронов нужна для постепенной утилизации этой энергии Кофермент Q Н+ Синтез АТФ и перенос эл-нов в матрикс Цитохром С IV V межмебранное пространство III I НАДН Н+ Молек. О 2 матрикс При переносе электронов по комплексам I-IV протоны Н+ попадают в межмембранное пространство. Генерируется эл/х потенциал. Комплекс V возвращает протоны в матрикс, а энергия потенциала используется для синтеза АТФ
На каждую переносимую пару электронов выделяется 52, 5 ккал/моль Цепь последовательного переноса электронов нужна для постепенной утилизации этой энергии Кофермент Q Н+ Синтез АТФ и перенос эл-нов в матрикс Цитохром С IV V межмебранное пространство III I НАДН Н+ Молек. О 2 матрикс При переносе электронов по комплексам I-IV протоны Н+ попадают в межмембранное пространство. Генерируется эл/х потенциал. Комплекс V возвращает протоны в матрикс, а энергия потенциала используется для синтеза АТФ
") II IV V III сукцинат Роль комплекса II - «сбор» электронов с сукцината (ЦТК)
II IV V III сукцинат Роль комплекса II - «сбор» электронов с сукцината (ЦТК)
 цитозоль + р. Н 7 Концентр. градиент р. Н 8 матрикс Электр. Потенциал ~14 в - Теория хемиосмотического сопряжения 1961 г. , П. Митчел
цитозоль + р. Н 7 Концентр. градиент р. Н 8 матрикс Электр. Потенциал ~14 в - Теория хемиосмотического сопряжения 1961 г. , П. Митчел
 Н+ Матрикс АДФ Комплекс V АТФ-синтаза АТФ Н+ АТФ-синтаза формирует протонный канал
Н+ Матрикс АДФ Комплекс V АТФ-синтаза АТФ Н+ АТФ-синтаза формирует протонный канал
 Транспорт электронов и окислительное фосфорилирование – главные функции белковых комплексов внутренней мембраны митохондрий, обеспечивающие клетку энергией. Большая часть АТФ, продуцируемого митохондриями, доставляется в цитоплазму с помощью АДФ/АТФ – транслоказы в обмен на АДФ Фосфат поступает в митохондрии вместе с протонами, независимо от транспорта АДФ/АТФ.
Транспорт электронов и окислительное фосфорилирование – главные функции белковых комплексов внутренней мембраны митохондрий, обеспечивающие клетку энергией. Большая часть АТФ, продуцируемого митохондриями, доставляется в цитоплазму с помощью АДФ/АТФ – транслоказы в обмен на АДФ Фосфат поступает в митохондрии вместе с протонами, независимо от транспорта АДФ/АТФ.
 «Конденсированная» «Ортодоксальная» Изменения топологии внутренней мембраны митохондрии в зависимости от содержания в ней АДФ Конденсированная и ортодоксальная конфигурации
«Конденсированная» «Ортодоксальная» Изменения топологии внутренней мембраны митохондрии в зависимости от содержания в ней АДФ Конденсированная и ортодоксальная конфигурации
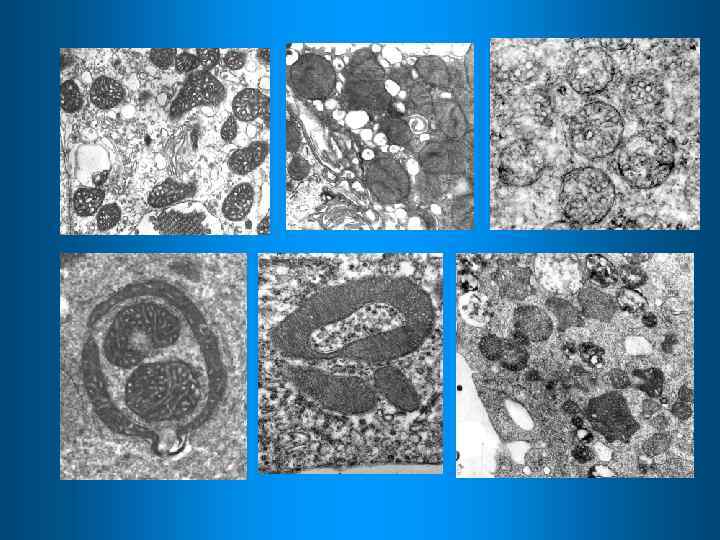
 Полученные в последние десятилетия сведения о свободных радикалах внесли существенный вклад в понимание физиологических процессов в норме и патологии. Установлено, что реактивные соединения кислорода и азота являются важными медиаторами клеточных реакций в ответ на стресс, а их действие опосредовано различными механизмами, включая аутофагию, митохондриальные реакции, передачу сигналов и апоптоз. Изменения структуры митохондрий при воздействии Н 2 О 2. А – интактные митохондрии. B, C, D – воздействие Н 2 О 2. Нарушение организации крист, изменение формы митохондрий, появление электронно-плотного вещества в матриксе.
Полученные в последние десятилетия сведения о свободных радикалах внесли существенный вклад в понимание физиологических процессов в норме и патологии. Установлено, что реактивные соединения кислорода и азота являются важными медиаторами клеточных реакций в ответ на стресс, а их действие опосредовано различными механизмами, включая аутофагию, митохондриальные реакции, передачу сигналов и апоптоз. Изменения структуры митохондрий при воздействии Н 2 О 2. А – интактные митохондрии. B, C, D – воздействие Н 2 О 2. Нарушение организации крист, изменение формы митохондрий, появление электронно-плотного вещества в матриксе.
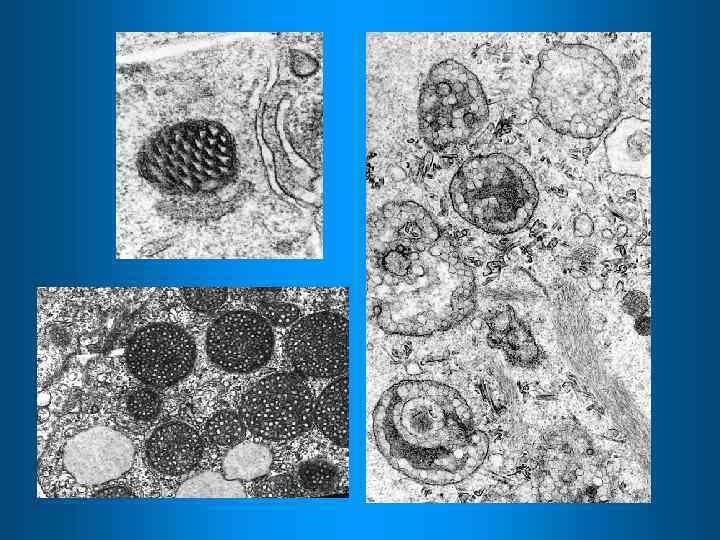
 Хлоропласты
Хлоропласты
 Хлоропласты
Хлоропласты
 Схема строения хлоропласта
Схема строения хлоропласта
 Граны в хлоропласте листа пшеницы
Граны в хлоропласте листа пшеницы
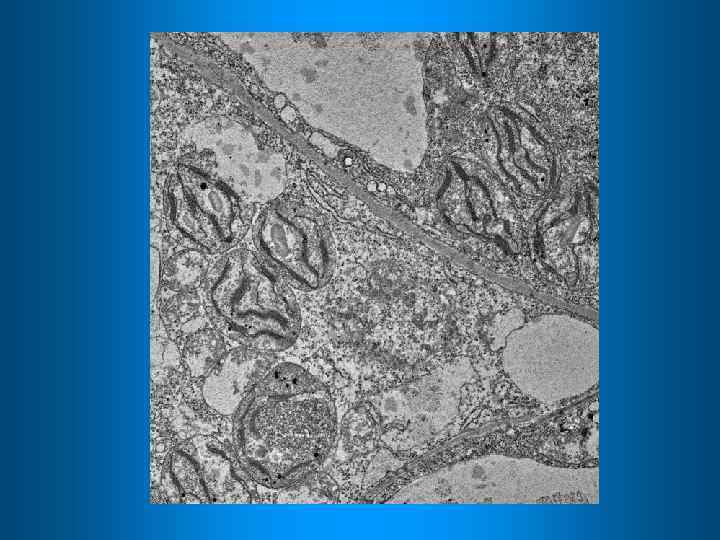
 Различные формы пластид способны к взаимопревращениям. Самые мелкие пластиды – пропластиды Пропластиды лейкопласты хлоропласты запасающие пластиды амилопласты липидопласты протеинопласты Хромопласты
Различные формы пластид способны к взаимопревращениям. Самые мелкие пластиды – пропластиды Пропластиды лейкопласты хлоропласты запасающие пластиды амилопласты липидопласты протеинопласты Хромопласты
 Недифференцированный лейкопласт запасающий лейкопласт хлоропласт хромопласт
Недифференцированный лейкопласт запасающий лейкопласт хлоропласт хромопласт
 АМИЛОПЛАСТ ХРОМОПЛАСТ Амилопласты риса
АМИЛОПЛАСТ ХРОМОПЛАСТ Амилопласты риса
 Хромопласты лепестков орхидей
Хромопласты лепестков орхидей