

Мутагенез, полиплоидия и гаплоидия.ppt
- Количество слайдов: 119
 Лекция 8. Мутационная селекция
Лекция 8. Мутационная селекция
 Задачи мутационной селекции 1. Обеспечение широкого спектра и частоты мутаций для получения исходного материала для отбора. 2. Получение мутантов для исправления отдельных недостатков сортов. 3. Увеличение рекомбинации генов и разрыв нежелательного сцепления генов. 4. Перенос фрагментов хромосом одного вида в хромосомы других видов при отделённой гибридизации. 5. Получение гомозиготных мутантов путём воздействия на гаплоиды излучениями и последующего удвоения у них числа хромосом.
Задачи мутационной селекции 1. Обеспечение широкого спектра и частоты мутаций для получения исходного материала для отбора. 2. Получение мутантов для исправления отдельных недостатков сортов. 3. Увеличение рекомбинации генов и разрыв нежелательного сцепления генов. 4. Перенос фрагментов хромосом одного вида в хромосомы других видов при отделённой гибридизации. 5. Получение гомозиготных мутантов путём воздействия на гаплоиды излучениями и последующего удвоения у них числа хромосом.
 Преимущества метода мутагенеза перед другими методами селекции 1. При улучшении одного или многих признаков существенно не перестраивается генотип улучшаемого сорта. 2. При мутагенезе возникает меньше гетерозиготных локусов в первом поколении, поэтому за более короткий срок можно получить константные, нерасщепляющиеся линии. 3. В результате мутаций у растений могут проявляться новые, ранее неизвестные, полезные признаки, для передачи которых гибридизация является не эффективной. 4. Действие мутагенов на генетический аппарат более упорядоченно, чем при спонтанном мутагенезе. Часть мутагенов совсем не вызывает хромосомных перестроек.
Преимущества метода мутагенеза перед другими методами селекции 1. При улучшении одного или многих признаков существенно не перестраивается генотип улучшаемого сорта. 2. При мутагенезе возникает меньше гетерозиготных локусов в первом поколении, поэтому за более короткий срок можно получить константные, нерасщепляющиеся линии. 3. В результате мутаций у растений могут проявляться новые, ранее неизвестные, полезные признаки, для передачи которых гибридизация является не эффективной. 4. Действие мутагенов на генетический аппарат более упорядоченно, чем при спонтанном мутагенезе. Часть мутагенов совсем не вызывает хромосомных перестроек.
 5. Мутагенезом можно вызывать взаимные транслокации хромосом у отдалённых гибридов, когда естественная рекомбинация не даёт желаемого эффекта. 6. При внутривидовой гибридизации частота рекомбинации тесно сцепленных генов может быть значительно повышена мутагенными факторами. 7. Устраняются нежелательные корреляции между признаками, поскольку хромосомные перестройки нарушают сцепление генов, определяющих коррелирующие признаки.
5. Мутагенезом можно вызывать взаимные транслокации хромосом у отдалённых гибридов, когда естественная рекомбинация не даёт желаемого эффекта. 6. При внутривидовой гибридизации частота рекомбинации тесно сцепленных генов может быть значительно повышена мутагенными факторами. 7. Устраняются нежелательные корреляции между признаками, поскольку хромосомные перестройки нарушают сцепление генов, определяющих коррелирующие признаки.
 Результаты облучения межвидовых гибридов F 1
Результаты облучения межвидовых гибридов F 1
 Классификация мутаций Мёллер предложил делить мутации по характеру изменения функционирования гена на гипоморфные (измененные аллели действуют в том же направлении, что и аллели дикого типа; синтезируется лишь меньше белкового продукта), аморфные (мутация выглядит, как полная потеря функции гена), антиморфные (мутантный признак изменяется, например, окраска зерна кукурузы меняется с пурпурного на бурый) и неоморфные.
Классификация мутаций Мёллер предложил делить мутации по характеру изменения функционирования гена на гипоморфные (измененные аллели действуют в том же направлении, что и аллели дикого типа; синтезируется лишь меньше белкового продукта), аморфные (мутация выглядит, как полная потеря функции гена), антиморфные (мутантный признак изменяется, например, окраска зерна кукурузы меняется с пурпурного на бурый) и неоморфные.
 Первичную мутацию иногда называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны.
Первичную мутацию иногда называют прямой мутацией, а мутацию, восстанавливающую исходную структуру гена, — обратной мутацией, или реверсией. Возврат к исходному фенотипу у мутантного организма вследствие восстановления функции мутантного гена нередко происходит не за счет истинной реверсии, а вследствие мутации в другой части того же самого гена или даже другого неаллельного гена. В этом случае возвратную мутацию называют супрессорной. Генетические механизмы, благодаря которым происходит супрессия мутантного фенотипа, весьма разнообразны.
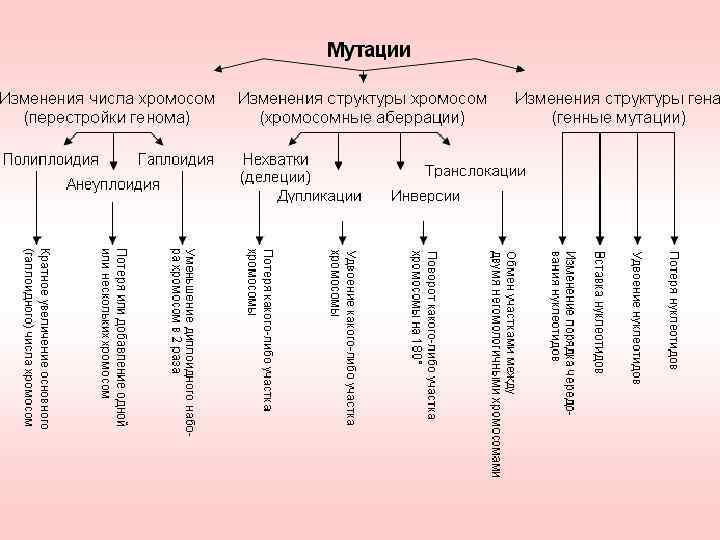
 Хромосомные мутации
Хромосомные мутации
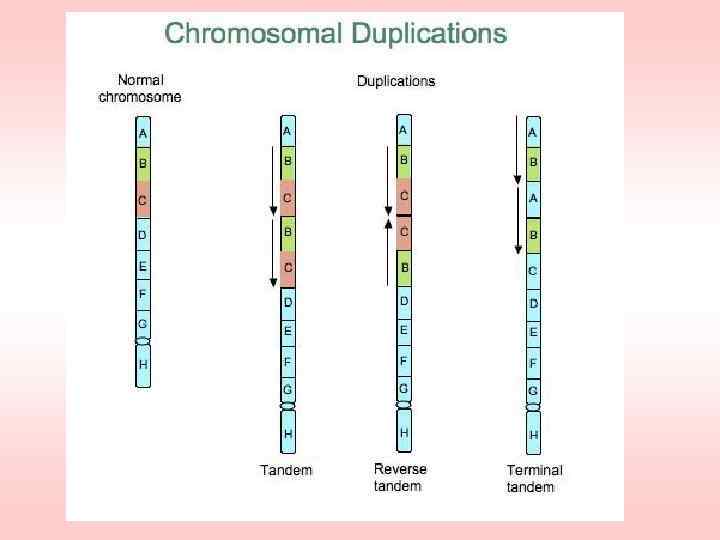
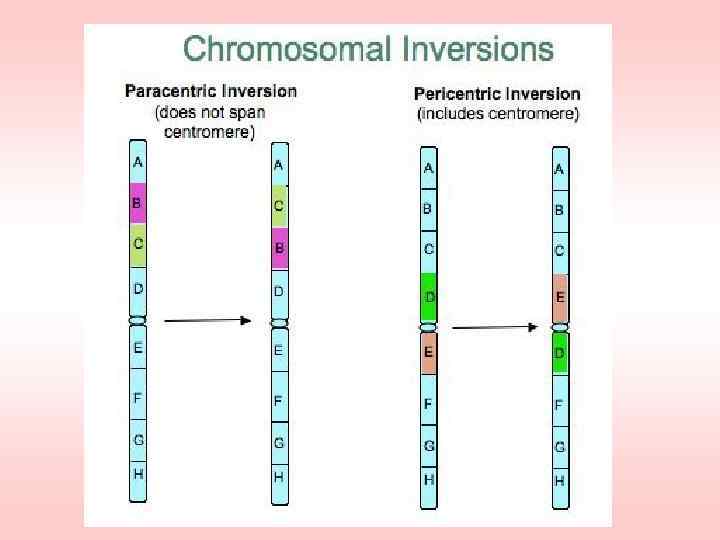
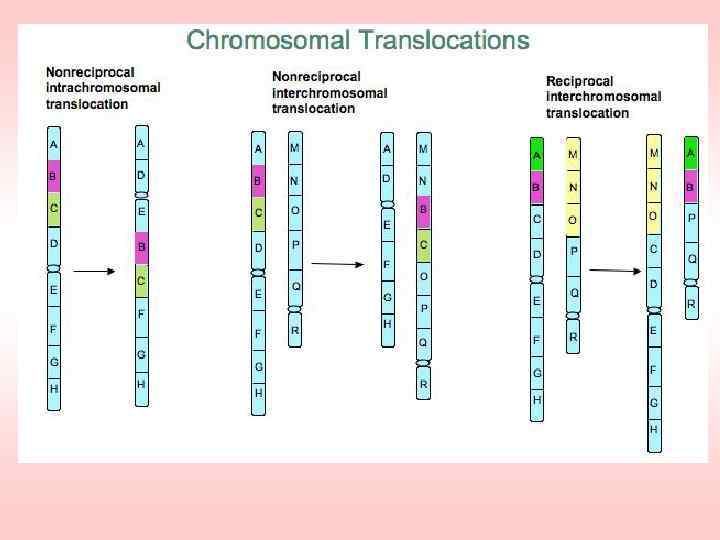
 Ионизирующие излучения –") Получение мутаций с помощью излучений Виды излучений, используемые в селекции: 1) Ионизирующие излучения – рентгеновские лучи, нейтроны, гамма лучи. 2) Неионизирующие излучения – ультрафиолетовые лучи, лазерное излучение.
Получение мутаций с помощью излучений Виды излучений, используемые в селекции: 1) Ионизирующие излучения – рентгеновские лучи, нейтроны, гамма лучи. 2) Неионизирующие излучения – ультрафиолетовые лучи, лазерное излучение.
 Источники ионизирующего излучения В природе ионизирующее излучение обычно генерируется в результате спонтанного радиоактивного распада радионуклидов, ядерных реакций (синтез и индуцированное деление ядер, захват протонов, нейтронов, альфа-частиц и др. ), а также при ускорении заряженных частиц в космосе. Искусственными источниками ионизирующего излучения являются искусственные радионуклиды (генерируют альфа-, бета- и гамма-излучения), ядерные реакторы (генерируют главным образом нейтронное и гаммаизлучение), радионуклидные нейтронные источники, ускорители элементарных частиц (генерируют потоки заряженных частиц, а также фотонное излучение), рентгеновские аппараты (генерируют рентгеновское излучение).
Источники ионизирующего излучения В природе ионизирующее излучение обычно генерируется в результате спонтанного радиоактивного распада радионуклидов, ядерных реакций (синтез и индуцированное деление ядер, захват протонов, нейтронов, альфа-частиц и др. ), а также при ускорении заряженных частиц в космосе. Искусственными источниками ионизирующего излучения являются искусственные радионуклиды (генерируют альфа-, бета- и гамма-излучения), ядерные реакторы (генерируют главным образом нейтронное и гаммаизлучение), радионуклидные нейтронные источники, ускорители элементарных частиц (генерируют потоки заряженных частиц, а также фотонное излучение), рентгеновские аппараты (генерируют рентгеновское излучение).
 Рентгеновские лучи - это, как и свет, электромагнитные волны, только с очень небольшой длиной волны.
Рентгеновские лучи - это, как и свет, электромагнитные волны, только с очень небольшой длиной волны.
 рентгеновскую трубку и вращающийся столик.") Ящик со свинцовой защитой, содержащий низковольтную (50 кв) рентгеновскую трубку и вращающийся столик.
Ящик со свинцовой защитой, содержащий низковольтную (50 кв) рентгеновскую трубку и вращающийся столик.
 Гамма-излучение также, как и рентгеновское излучение является электромагнитным излучением, но возникает оно при распаде радиоактивных изотопов, обычно 60 Со, 137 Cs или 226 Ra. Биологические объекты облучают гамма лучами двумя способами: 1) Острое облучение (кратковременное излучение из мощного источника) – контейнер с семенами, клубнями и др. опускают под воду на глубину 3 метра между восьмью стержнями из радиоактивного кобальта. 2) Хроническое облучение (длительное, но значительно более слабое) – обработка растений на гамма поле.
Гамма-излучение также, как и рентгеновское излучение является электромагнитным излучением, но возникает оно при распаде радиоактивных изотопов, обычно 60 Со, 137 Cs или 226 Ra. Биологические объекты облучают гамма лучами двумя способами: 1) Острое облучение (кратковременное излучение из мощного источника) – контейнер с семенами, клубнями и др. опускают под воду на глубину 3 метра между восьмью стержнями из радиоактивного кобальта. 2) Хроническое облучение (длительное, но значительно более слабое) – обработка растений на гамма поле.
 Схема гамма-установки для острого облучения: 1 – подъёмный механизм; 2 – камера с объектом; 3 – пульт автоматического управления; 4 – бак, заполненный водой; 5 – восемь стержней, набранных из радиоактивного кобальта (60 Со); 6 – пробка аварийного отсека; 7 – аварийный отсек.
Схема гамма-установки для острого облучения: 1 – подъёмный механизм; 2 – камера с объектом; 3 – пульт автоматического управления; 4 – бак, заполненный водой; 5 – восемь стержней, набранных из радиоактивного кобальта (60 Со); 6 – пробка аварийного отсека; 7 – аварийный отсек.
") Гамма поле института радиационной селекции (Япония)
Гамма поле института радиационной селекции (Япония)

 Экспериментальный комплекс «Гамма-поле» Молдавской опытной станции по эфиро-масличным культурам. 1987 -й год.
Экспериментальный комплекс «Гамма-поле» Молдавской опытной станции по эфиро-масличным культурам. 1987 -й год.
; 2") Схема размещения гамма-установки на гамма-поле: 1 – контейнер для хранения гамма-излучателя (60 Со); 2 и 4 – система подъёма и спуска излучателя; 3 – управление установкой; 5 – защитный контейнер.
Схема размещения гамма-установки на гамма-поле: 1 – контейнер для хранения гамма-излучателя (60 Со); 2 и 4 – система подъёма и спуска излучателя; 3 – управление установкой; 5 – защитный контейнер.
 Оптимальные дозы гамма- и рентгеновского излучения для сухих семян, дающие высокий выход хозяйственнополезных мутаций Культура Доза, Гр Пшеница яровая и озимая 50 -100 Горох кормовой 80 -120 Ячмень яровой Овёс 50 -100 Люпин 140 -160 70 -100 Бобы 30 -40 Кукуруза 50 -100 Фасоль 50 -70 Горох зерновой 50 -100 Соя 50 -80
Оптимальные дозы гамма- и рентгеновского излучения для сухих семян, дающие высокий выход хозяйственнополезных мутаций Культура Доза, Гр Пшеница яровая и озимая 50 -100 Горох кормовой 80 -120 Ячмень яровой Овёс 50 -100 Люпин 140 -160 70 -100 Бобы 30 -40 Кукуруза 50 -100 Фасоль 50 -70 Горох зерновой 50 -100 Соя 50 -80
 — электромагнитное излучение, занимающее диапазон между видимым и рентгеновским") Ультрафиолетовое излучение (ультрафиолет, УФ, UV) — электромагнитное излучение, занимающее диапазон между видимым и рентгеновским излучением. Источником ультрафиолетовых лучей являются ртутные, водородные, ксеноновые и др. газоразрядные лампы, окна которых (либо целиком колбы) изготовляют из прозрачных для ультрафиолетового излучения материалов (чаще из кварца). Для ультрафиолетовой области спектра разработаны также оптические квантовые генераторы (лазеры).
Ультрафиолетовое излучение (ультрафиолет, УФ, UV) — электромагнитное излучение, занимающее диапазон между видимым и рентгеновским излучением. Источником ультрафиолетовых лучей являются ртутные, водородные, ксеноновые и др. газоразрядные лампы, окна которых (либо целиком колбы) изготовляют из прозрачных для ультрафиолетового излучения материалов (чаще из кварца). Для ультрафиолетовой области спектра разработаны также оптические квантовые генераторы (лазеры).
 Проникающая способность ультрафиолетовых лучей очень мала. При УФ облучении происходит только возбуждение молекул. Поэтому УФ излучение можно использовать только для обработки пыльцевых зёрен. УФ излучение взывает высокую частоту мутаций в пределах длины волны, которая поглощается ДНК (260 -265 нм). Дозу ультрафиолетового излучения выражают в Дж/м кв.
Проникающая способность ультрафиолетовых лучей очень мала. При УФ облучении происходит только возбуждение молекул. Поэтому УФ излучение можно использовать только для обработки пыльцевых зёрен. УФ излучение взывает высокую частоту мутаций в пределах длины волны, которая поглощается ДНК (260 -265 нм). Дозу ультрафиолетового излучения выражают в Дж/м кв.
 Нейтроны образуются в результате некоторых ядерных реакций, в частности при делении ядер урана и плутония. Облучение нейтронами проводят с помощью ядерного реактора или циклотрона. При обработке семян быстрыми нейтронами оптимальная доза для пшеницы и ячменя составляет 2, 5 -7, 5 Гр, для кукурузы – 3 -5, для проса – 5 -10, для гороха – 2 -3 Гр. При использовании тепловых нейтронов оптимальная доза (н/см кв. ) равна: для пшеницы – 1 х 10 в 10 степени – 1 х 10 в 13 степени, для ячменя 1 х 10 в 9 степени – 1 х 10 в 11 степени, для кукурузы – 6 х 10 в 7 степени – 1 х 10 в 10 степени.
Нейтроны образуются в результате некоторых ядерных реакций, в частности при делении ядер урана и плутония. Облучение нейтронами проводят с помощью ядерного реактора или циклотрона. При обработке семян быстрыми нейтронами оптимальная доза для пшеницы и ячменя составляет 2, 5 -7, 5 Гр, для кукурузы – 3 -5, для проса – 5 -10, для гороха – 2 -3 Гр. При использовании тепловых нейтронов оптимальная доза (н/см кв. ) равна: для пшеницы – 1 х 10 в 10 степени – 1 х 10 в 13 степени, для ячменя 1 х 10 в 9 степени – 1 х 10 в 11 степени, для кукурузы – 6 х 10 в 7 степени – 1 х 10 в 10 степени.
 Циклотрон и ядерный реактор института ядерной физики в Санкт-Питербурге
Циклотрон и ядерный реактор института ядерной физики в Санкт-Питербурге
") При воздействии на семена и вегетирующие растения импульсным и непрерывным (в том числе низкоинтенсивным) лазерным излучением видимой области спектра можно получить высокий выход мутантных форм. С большой частотой возникают мутации с повышенной продуктивностью, не наблюдается летального эффекта у растений первого поколения. Например, из сорта мягкой пшеницы Ленинградка выделена серия ценных перспективных мутантов ЛМ 1, ЛМ-2, …, ЛМ-8, которые переданы в сортоиспытание или включены в гибридизацию. Из сорта Надя выделено более 20 ценных мутантов, характеризующиеся скороспелостью, низкорослостью, высокой продуктивностью и повышенным содержанием белка.
При воздействии на семена и вегетирующие растения импульсным и непрерывным (в том числе низкоинтенсивным) лазерным излучением видимой области спектра можно получить высокий выход мутантных форм. С большой частотой возникают мутации с повышенной продуктивностью, не наблюдается летального эффекта у растений первого поколения. Например, из сорта мягкой пшеницы Ленинградка выделена серия ценных перспективных мутантов ЛМ 1, ЛМ-2, …, ЛМ-8, которые переданы в сортоиспытание или включены в гибридизацию. Из сорта Надя выделено более 20 ценных мутантов, характеризующиеся скороспелостью, низкорослостью, высокой продуктивностью и повышенным содержанием белка.
 Работа гелий-неонового лазера
Работа гелий-неонового лазера
 Импульсный лазер
Импульсный лазер
 Поглощенная и экспозиционная доза Поглощенная доза показывает, какое количество энергии излучения поглощено в единице массы любого облучаемого вещества и определяется отношением поглощенной энергии ионизирующего излучения на массу вещества. За единицу измерения поглощенной дозы в системе СИ принят грэй (Гр). 1 Гр — это такая доза, при которой массе 1 кг передается энергия ионизирующего излучения 1 Дж. Внесистемной единицей поглощенной дозы является рад. 1 Гр=100 рад; 1 Р (рентген)=1 рад, или 1 к. Р (килорентген)=10 Гр. Рентген – экспозиционная доза, которая устанавливается по степени ионизации воздуха источником ионизирующего излучения.
Поглощенная и экспозиционная доза Поглощенная доза показывает, какое количество энергии излучения поглощено в единице массы любого облучаемого вещества и определяется отношением поглощенной энергии ионизирующего излучения на массу вещества. За единицу измерения поглощенной дозы в системе СИ принят грэй (Гр). 1 Гр — это такая доза, при которой массе 1 кг передается энергия ионизирующего излучения 1 Дж. Внесистемной единицей поглощенной дозы является рад. 1 Гр=100 рад; 1 Р (рентген)=1 рад, или 1 к. Р (килорентген)=10 Гр. Рентген – экспозиционная доза, которая устанавливается по степени ионизации воздуха источником ионизирующего излучения.
 Закономерности радиационного мутагенеза Частота появления мутаций зависит от дозы облучения. Существует положительная линейная зависимость между дозой облучения и частотой мутаций. Однако эта пропорциональность наблюдается в основном в интервале 100 %-ной выживаемости. С переходом в интервал до ЛД 0 -ЛД 100 выход мутаций уже не может возрастать пропорционально дозе, так как с повышением дозы будет увеличиваться гибель растений, относительное число жизнеспособных мутантов начинает падать. ЛД 0 -порог виживаемости, ЛД 100 – гибель всех растений.
Закономерности радиационного мутагенеза Частота появления мутаций зависит от дозы облучения. Существует положительная линейная зависимость между дозой облучения и частотой мутаций. Однако эта пропорциональность наблюдается в основном в интервале 100 %-ной выживаемости. С переходом в интервал до ЛД 0 -ЛД 100 выход мутаций уже не может возрастать пропорционально дозе, так как с повышением дозы будет увеличиваться гибель растений, относительное число жизнеспособных мутантов начинает падать. ЛД 0 -порог виживаемости, ЛД 100 – гибель всех растений.
 Факторы от которых зависит доза облучения 1. Видовая принадлежность растения. 2. Сортовая принадлежность (разные сорта одного вида с одинаковым набором хромосом могут иметь разную радиочувствительность). 3. Облучаемые органы (семена обрабатывают большей дозой, чем вегетативные органы) 4. Физиологическое состояние органов растения (покоящиеся семена обрабатывают большей дозой, чем прорастающие). 5. Плоидность (тетраплоиды гречихи более устойчивы к гамма-излучению, чем диплоиды).
Факторы от которых зависит доза облучения 1. Видовая принадлежность растения. 2. Сортовая принадлежность (разные сорта одного вида с одинаковым набором хромосом могут иметь разную радиочувствительность). 3. Облучаемые органы (семена обрабатывают большей дозой, чем вегетативные органы) 4. Физиологическое состояние органов растения (покоящиеся семена обрабатывают большей дозой, чем прорастающие). 5. Плоидность (тетраплоиды гречихи более устойчивы к гамма-излучению, чем диплоиды).
 Критическая доза радиации – доза, выше которой уже наблюдается значительное снижение жизнеспособности и плодовитости растений, а затем и их гибель. За критическую дозу принимают такую дозу, при которой выживают 30 -40% растений (ЛД 30 -40). При высоких дозах излучения, дающих сильный повреждающий эффект, доля хозяйственно-ценных мутаций меньше, чем при средних. Поэтому в селекционных целях рекомендуют дозы излучения в 1, 5 -2 раза ниже критических. Лучшими считаются те, которые лишь незначительно снижают всхожесть семян и мало угнетают рост растений. После воздействия рентгеновским и гамма излучением и быстрыми нейтронами семена можно сразу высевать, т. к. они не имеют наведённой радиации.
Критическая доза радиации – доза, выше которой уже наблюдается значительное снижение жизнеспособности и плодовитости растений, а затем и их гибель. За критическую дозу принимают такую дозу, при которой выживают 30 -40% растений (ЛД 30 -40). При высоких дозах излучения, дающих сильный повреждающий эффект, доля хозяйственно-ценных мутаций меньше, чем при средних. Поэтому в селекционных целях рекомендуют дозы излучения в 1, 5 -2 раза ниже критических. Лучшими считаются те, которые лишь незначительно снижают всхожесть семян и мало угнетают рост растений. После воздействия рентгеновским и гамма излучением и быстрыми нейтронами семена можно сразу высевать, т. к. они не имеют наведённой радиации.
 Способы обработки излучениями
Способы обработки излучениями
 Химический мутагенез Известно множество химических соединений, обладаюших мутагенной активностью: алкилирующие препараты, перекиси, гидроксиламины, аналоги оснований ДНК, соли тяжелых металлов, некоторые ароматические соединения (алкалоиды, лекарственные препараты, гербициды) и др. Большую группу химических мутагенов представляют алкилирующие соединения. В настоящее время в селекции широко применяют следующие мутагены: нитрозопроизводные мочевины — нитрозометилмочевина (НММ), нитрозоэтилмочевина (НЭМ), нитрозодиметилмочевина (НДММ); этиленимин (ЭИ), этилметансульфонат (ЭМС), диметилсульфат (ДМС), 1, 4 -бисдиазоацетилбутан (1, 4 -ДАБ), колхицин и др.
Химический мутагенез Известно множество химических соединений, обладаюших мутагенной активностью: алкилирующие препараты, перекиси, гидроксиламины, аналоги оснований ДНК, соли тяжелых металлов, некоторые ароматические соединения (алкалоиды, лекарственные препараты, гербициды) и др. Большую группу химических мутагенов представляют алкилирующие соединения. В настоящее время в селекции широко применяют следующие мутагены: нитрозопроизводные мочевины — нитрозометилмочевина (НММ), нитрозоэтилмочевина (НЭМ), нитрозодиметилмочевина (НДММ); этиленимин (ЭИ), этилметансульфонат (ЭМС), диметилсульфат (ДМС), 1, 4 -бисдиазоацетилбутан (1, 4 -ДАБ), колхицин и др.
 — желтый блестящий кристаллический порошок. Хорошо растворим в") 1, 4 -бисдиазоацетилбутан (1, 4 -ДАБ) — желтый блестящий кристаллический порошок. Хорошо растворим в воде и органических растворителях. Химический мутаген, очень ядовит. Диметилсульфат (ДМС) — бесцветная жидкость без запаха, химический мутаген. В воде плохо растворяется. Хорошо растворяется в спирте, эфире. Очень ядовита. Нитрозометилмочевина (НММ) — белый порошок, хорошо растворимый в воде, спирте, эфире; химический мутаген, разлагается при комнатной температуре, ядовит. Нитрозоэтилмочевина (НЭМ) — кремовато-белый порошок, химический мутаген. Хорошо растворимый в воде, спирте, эфире. Ядовит.
1, 4 -бисдиазоацетилбутан (1, 4 -ДАБ) — желтый блестящий кристаллический порошок. Хорошо растворим в воде и органических растворителях. Химический мутаген, очень ядовит. Диметилсульфат (ДМС) — бесцветная жидкость без запаха, химический мутаген. В воде плохо растворяется. Хорошо растворяется в спирте, эфире. Очень ядовита. Нитрозометилмочевина (НММ) — белый порошок, хорошо растворимый в воде, спирте, эфире; химический мутаген, разлагается при комнатной температуре, ядовит. Нитрозоэтилмочевина (НЭМ) — кремовато-белый порошок, химический мутаген. Хорошо растворимый в воде, спирте, эфире. Ядовит.
 — кремовато-белый порошок, в воде плохо растворим. При комнатной температуре разлагается со") Этилметансульфонат (ЭМС) — кремовато-белый порошок, в воде плохо растворим. При комнатной температуре разлагается со взрывом. Химический мутаген. Ядовит. Этиленимин, азиридин, бесцветная легкоподвижная жидкость с запахом аммиака. Этиленимин сильно ядовит; предельно допустимая концентрация паров Этиленимин в воздухе 0, 02 мг/м 3. В жидком состоянии обладает сильным кожным действием.
Этилметансульфонат (ЭМС) — кремовато-белый порошок, в воде плохо растворим. При комнатной температуре разлагается со взрывом. Химический мутаген. Ядовит. Этиленимин, азиридин, бесцветная легкоподвижная жидкость с запахом аммиака. Этиленимин сильно ядовит; предельно допустимая концентрация паров Этиленимин в воздухе 0, 02 мг/м 3. В жидком состоянии обладает сильным кожным действием.
 Концентрации химических мутагенов для обработки сухих семян Мутаген Этиленимин Концентрация водного раствора, % (по объёму) 0, 01 -0, 06 Этилметансульфонат 0, 1 -0, 5 Гиброксиламин 1, 5 -3, 0 1, 4 -Бисдиазоацетилбутан 0, 2 -0, 5 Диметилсульфат 0, 01 -0, 2 Нитрозоэтилмочевина 0, 01 -0, 05 Нитрозометилмочевина 0, 001 -0, 005
Концентрации химических мутагенов для обработки сухих семян Мутаген Этиленимин Концентрация водного раствора, % (по объёму) 0, 01 -0, 06 Этилметансульфонат 0, 1 -0, 5 Гиброксиламин 1, 5 -3, 0 1, 4 -Бисдиазоацетилбутан 0, 2 -0, 5 Диметилсульфат 0, 01 -0, 2 Нитрозоэтилмочевина 0, 01 -0, 05 Нитрозометилмочевина 0, 001 -0, 005
 Обрабатываемые органы и продолжительность обработки Химическими мутагенами можно обрабатывать сухие и проросшие семена, черенки, клубни, луковицы, инъецировать эти вещества в стебель растений перед вступлением их в генеративную фазу и т. д. Продолжительность обработки семян (экспозиция) варьирует от 3 до 18 часов.
Обрабатываемые органы и продолжительность обработки Химическими мутагенами можно обрабатывать сухие и проросшие семена, черенки, клубни, луковицы, инъецировать эти вещества в стебель растений перед вступлением их в генеративную фазу и т. д. Продолжительность обработки семян (экспозиция) варьирует от 3 до 18 часов.
 Методы обработки семян, проростков и завязей химическими мутагенами 1. Обработка семян парами химических мутагенов ( «обработка в газовой фазе» ). 2. Обработка клеток стеблевой меристемы у проростков. 3. Применение химических мутагенов вместе со стимуляторами роста. 4. Обработка завязей ячменя через 48– 72 ч после оплодотворения. 5. Совместное использование специфических ферментов и химических мутагенов. 6. Обработка методом вакуум-инфильтрации.
Методы обработки семян, проростков и завязей химическими мутагенами 1. Обработка семян парами химических мутагенов ( «обработка в газовой фазе» ). 2. Обработка клеток стеблевой меристемы у проростков. 3. Применение химических мутагенов вместе со стимуляторами роста. 4. Обработка завязей ячменя через 48– 72 ч после оплодотворения. 5. Совместное использование специфических ферментов и химических мутагенов. 6. Обработка методом вакуум-инфильтрации.
 Индуцирование и отбор плотноколосового эректоидного мутанта у ячменя при воздействии излучением на семена и растения перед их цветением (в кружках показаны исходные (А) и мутировавшие (а) гены): 1 – растение исходного сорта; 2 – мутировавшее растение М 1; 3 – неизменённое растение М 2; 4 – фенотипически не изменённое растение М 2; 5 – изменённое растение М 2
Индуцирование и отбор плотноколосового эректоидного мутанта у ячменя при воздействии излучением на семена и растения перед их цветением (в кружках показаны исходные (А) и мутировавшие (а) гены): 1 – растение исходного сорта; 2 – мутировавшее растение М 1; 3 – неизменённое растение М 2; 4 – фенотипически не изменённое растение М 2; 5 – изменённое растение М 2
 Ход селекционного процесса на основе мутагенеза
Ход селекционного процесса на основе мутагенеза
 Первый год: белые кружки – выжившие, неизменённые растения; заштрихованные кружки – выжившие, внешне не изменённые, но мутировавшие (гетерозиготные) растения; плюс (+) – растения не взошли или погибли позже. А – раздельное выращивание потомств всех выживших растений. Второй год: белые кружки – неизменённые растения; чёрные кружки – внешне изменённые, вероятно, мутировавшие растения. Б – выращивание изменённых растений для проверки константности возникших изменений. Третий год: чёрные кружки – изменения наследуются (мутанты); белые кружки – изменения не наследуются (в М 2 были отобраны модифицировавшие растения). Четвёртый и последующие годы: испытание мутантов на урожайность, устойчивость к полеганию и другие полезные признаки
Первый год: белые кружки – выжившие, неизменённые растения; заштрихованные кружки – выжившие, внешне не изменённые, но мутировавшие (гетерозиготные) растения; плюс (+) – растения не взошли или погибли позже. А – раздельное выращивание потомств всех выживших растений. Второй год: белые кружки – неизменённые растения; чёрные кружки – внешне изменённые, вероятно, мутировавшие растения. Б – выращивание изменённых растений для проверки константности возникших изменений. Третий год: чёрные кружки – изменения наследуются (мутанты); белые кружки – изменения не наследуются (в М 2 были отобраны модифицировавшие растения). Четвёртый и последующие годы: испытание мутантов на урожайность, устойчивость к полеганию и другие полезные признаки
 Питомник М 1 С учётом частичной гибели растений в М 1, количество семян для облучения необходимо брать в 2 – 2, 5 раза больше, чем требуется семей М 2. Облученные семена пшеницы и ячменя лучше высевать в теплице или парнике и выращивать там до фазы 3– 4 настоящих листьев, затем пересаживать в поле. Зернобобовые культуры плохо переносят пересадку, поэтому их сразу высевают в поле. Для контроля высевают необлученные семена. Первое мутантное поколение М 1 у зерновых культур (пшеницы, ячменя) сеют рядовым способом на высоком агрофоне с междурядьями 15 см и расстоянием между растениями – 1– 2 см. Зернобобовые (горох, вику, сою, фасоль) сеют широкорядно с междурядьями 45– 50 см и расстоянием в рядке 10– 15 см.
Питомник М 1 С учётом частичной гибели растений в М 1, количество семян для облучения необходимо брать в 2 – 2, 5 раза больше, чем требуется семей М 2. Облученные семена пшеницы и ячменя лучше высевать в теплице или парнике и выращивать там до фазы 3– 4 настоящих листьев, затем пересаживать в поле. Зернобобовые культуры плохо переносят пересадку, поэтому их сразу высевают в поле. Для контроля высевают необлученные семена. Первое мутантное поколение М 1 у зерновых культур (пшеницы, ячменя) сеют рядовым способом на высоком агрофоне с междурядьями 15 см и расстоянием между растениями – 1– 2 см. Зернобобовые (горох, вику, сою, фасоль) сеют широкорядно с междурядьями 45– 50 см и расстоянием в рядке 10– 15 см.
 Второй мутантный питомник Поскольку мутации возникают в отдельных клетках семени, то зародыш, а затем взрослое растение является химерой (одни колосья-мутантные, другие – нормальные), поэтому семена (20 шт. ) из каждого колоса растений М 1 (для зерновых) , ветки или растения М 1 (для зернобобовых) высевают в М 2 отдельными линиями. Средний объём анализируемого в М 2 материала должен составлять от 200 до 1000 линий; для зернобобовых в случае уборки М 1 по растениям – 300– 400 линий. В одной линии должно быть не менее 15 растений (у зерновых) или 25 (у бобовых). При этом в каждой линии можно выявить обычно 2– 5 мутантов.
Второй мутантный питомник Поскольку мутации возникают в отдельных клетках семени, то зародыш, а затем взрослое растение является химерой (одни колосья-мутантные, другие – нормальные), поэтому семена (20 шт. ) из каждого колоса растений М 1 (для зерновых) , ветки или растения М 1 (для зернобобовых) высевают в М 2 отдельными линиями. Средний объём анализируемого в М 2 материала должен составлять от 200 до 1000 линий; для зернобобовых в случае уборки М 1 по растениям – 300– 400 линий. В одной линии должно быть не менее 15 растений (у зерновых) или 25 (у бобовых). При этом в каждой линии можно выявить обычно 2– 5 мутантов.
 В М 2 выявляются рецессивные мутации. Доминантные и полудоминантные мутации (безостые, плотноколосые, спельтоиды у пшеницы, с антоциановым венчиком у гороха) выявляются уже в М 1, но они встречаются реже. Отбор изменённых форм по морфологическим признакам, устойчивости к болезням, скороспелости проводят в М 2 в течение всей вегетации. Мутанты по количественным признакам (элементы структуры урожая) выявляют после уборки путём лабораторного анализа. В М 2 обычно отбирают в зависимости от культуры от 1 до 3% потомств (линий), в которых предполагают наличие полезных мутаций.
В М 2 выявляются рецессивные мутации. Доминантные и полудоминантные мутации (безостые, плотноколосые, спельтоиды у пшеницы, с антоциановым венчиком у гороха) выявляются уже в М 1, но они встречаются реже. Отбор изменённых форм по морфологическим признакам, устойчивости к болезням, скороспелости проводят в М 2 в течение всей вегетации. Мутанты по количественным признакам (элементы структуры урожая) выявляют после уборки путём лабораторного анализа. В М 2 обычно отбирают в зависимости от культуры от 1 до 3% потомств (линий), в которых предполагают наличие полезных мутаций.
, поскольку") Питомник М 3 Семена от неизмененных форм высевают сплошным способом (не по линиям), поскольку часть мутантов может выщепиться и в М 3. Семена от изменённых форм, выделенных в М 2, высевают для получения потомств М 3 отдельными линиями. Для оценки на продуктивность в М 3 через 10– 20 номеров мутантных линий высевают семена исходного сорта и сорта-стандарта. Мутанты, выделившиеся по урожайности и по другим хозяйственно-ценным признакам, переводят в контрольный или селекционный питомник первого года (СП-1).
Питомник М 3 Семена от неизмененных форм высевают сплошным способом (не по линиям), поскольку часть мутантов может выщепиться и в М 3. Семена от изменённых форм, выделенных в М 2, высевают для получения потомств М 3 отдельными линиями. Для оценки на продуктивность в М 3 через 10– 20 номеров мутантных линий высевают семена исходного сорта и сорта-стандарта. Мутанты, выделившиеся по урожайности и по другим хозяйственно-ценным признакам, переводят в контрольный или селекционный питомник первого года (СП-1).
 Использование мутаций в сочетании с гибридизацией Возможны такие комбинации скрещиваний: мутант с исходным сортом; разные мутанты между собой в пределах одного и того же сорта; мутанты, выделенные у разных сортов, при воздействии разными мутагенами. Ценный материал для гибридизации представляют мутанты, у которых один-два важных признака изменены в положительную сторону, хотя по общей продуктивности они не превосходят исходный сорт. Например, у гороха часто встречаются низкорослые сильно ветвящиеся мутанты. При скрещивании их с исходным сортом, как показали опыты К. К. Сидоровой, удаётся получать высокорослые ветвящиеся растения, отличающиеся высокой продуктивностью.
Использование мутаций в сочетании с гибридизацией Возможны такие комбинации скрещиваний: мутант с исходным сортом; разные мутанты между собой в пределах одного и того же сорта; мутанты, выделенные у разных сортов, при воздействии разными мутагенами. Ценный материал для гибридизации представляют мутанты, у которых один-два важных признака изменены в положительную сторону, хотя по общей продуктивности они не превосходят исходный сорт. Например, у гороха часто встречаются низкорослые сильно ветвящиеся мутанты. При скрещивании их с исходным сортом, как показали опыты К. К. Сидоровой, удаётся получать высокорослые ветвящиеся растения, отличающиеся высокой продуктивностью.
 У пшеницы встречаются мутанты, устойчивые к полеганию, но с низкими хлебопекарными качествами или мутанты с очень упругой клейковиной, но сильно полегающие. В опытах И. В. Чёрного скрещивание таких мутантов между собой позволило получить формы, устойчивые к полеганию, с высокими хлебопекарными качествами. При использовании мутагенеза проводят «накопительные» скрещивания. Если в результате воздействия мутагенами появились аддитивно ценные мутации, то они наверняка возникли в разных растениях и максимальный эффект могут дать в том случае, когда окажутся в одной линии. Для объединения аддитивно ценных мутаций проводят накопительные скрещивания полученных после воздействия мутагенами линий по схеме: [(А х В) х (С х Д) х (Е х Ж) и т. д. ].
У пшеницы встречаются мутанты, устойчивые к полеганию, но с низкими хлебопекарными качествами или мутанты с очень упругой клейковиной, но сильно полегающие. В опытах И. В. Чёрного скрещивание таких мутантов между собой позволило получить формы, устойчивые к полеганию, с высокими хлебопекарными качествами. При использовании мутагенеза проводят «накопительные» скрещивания. Если в результате воздействия мутагенами появились аддитивно ценные мутации, то они наверняка возникли в разных растениях и максимальный эффект могут дать в том случае, когда окажутся в одной линии. Для объединения аддитивно ценных мутаций проводят накопительные скрещивания полученных после воздействия мутагенами линий по схеме: [(А х В) х (С х Д) х (Е х Ж) и т. д. ].
 Мутанты, выделенные в одних почвеннометеорологических условиях, могут оказаться более продуктивными в других условиях. Например, в опытах К. К. Сидоровой и Г. А. Дебелого компактный, более устойчивый к полеганию мутант гороха, выделенный из сорта Торсдаг, при испытании в Новосибирской области уступал по урожайности исходному сорту, а в условиях Подмосковья дал значительную прибавку урожая.
Мутанты, выделенные в одних почвеннометеорологических условиях, могут оказаться более продуктивными в других условиях. Например, в опытах К. К. Сидоровой и Г. А. Дебелого компактный, более устойчивый к полеганию мутант гороха, выделенный из сорта Торсдаг, при испытании в Новосибирской области уступал по урожайности исходному сорту, а в условиях Подмосковья дал значительную прибавку урожая.
 Получение направленных мутаций При решении одних задач селекции подходят химические мутагены, а для решения других – излучения. При работе с ячменём и озимой пшеницей было показано, что эректоидные формы (низкорослые, с прочной соломиной) чаще возникают при воздействии излучений с высокой линейной потерей энергии (ЛПЭ) – быстрые нейтроны, альфа-частицы. Устойчивые к заболеваниям формы часто возникают при обработке семян этиленимином в низких концентрациях, а также при обработке тепловыми нейтронами.
Получение направленных мутаций При решении одних задач селекции подходят химические мутагены, а для решения других – излучения. При работе с ячменём и озимой пшеницей было показано, что эректоидные формы (низкорослые, с прочной соломиной) чаще возникают при воздействии излучений с высокой линейной потерей энергии (ЛПЭ) – быстрые нейтроны, альфа-частицы. Устойчивые к заболеваниям формы часто возникают при обработке семян этиленимином в низких концентрациях, а также при обработке тепловыми нейтронами.
 Для преимущественного получения перестроек хромосом проводят облучение быстрыми нейтронами, а для выявления преимущественно точковых мутантов – обработку невысокими концентрациями этиленимина (0, 01– 0, 04%), или этилметансульфоната (0, 1– 0, 2%), или облучение гамма-лучами с дополнительной обработкой. Если излучения используют для получения транслокаций, вставок, разрыва сцепления между локусами, рекомендуется облучать растения во время мейоза.
Для преимущественного получения перестроек хромосом проводят облучение быстрыми нейтронами, а для выявления преимущественно точковых мутантов – обработку невысокими концентрациями этиленимина (0, 01– 0, 04%), или этилметансульфоната (0, 1– 0, 2%), или облучение гамма-лучами с дополнительной обработкой. Если излучения используют для получения транслокаций, вставок, разрыва сцепления между локусами, рекомендуется облучать растения во время мейоза.
 Типы полезных изменений, возникающие при использовании мутагенных факторов 1. Получение короткостебельных растений и мутантов с повышенной прочностью стебля у ячменя, пшеницы, овса, риса. 2. Создание форм, сочетающих урожайность и скороспелость. 3. Создание сортов пшеницы, ячменя и др. культур, устойчивых к болезням. 4. Улучшение хлебопекарных качеств сортов пшеницы, создание высокоолеиновых сортов подсолнечника. 5. Создание высокопродуктивных сортов пшеницы. 6. Получение безалкалоидных сортов кормового люпина с увеличенным количеством бобов на растении и высокой семенной продуктивностью.
Типы полезных изменений, возникающие при использовании мутагенных факторов 1. Получение короткостебельных растений и мутантов с повышенной прочностью стебля у ячменя, пшеницы, овса, риса. 2. Создание форм, сочетающих урожайность и скороспелость. 3. Создание сортов пшеницы, ячменя и др. культур, устойчивых к болезням. 4. Улучшение хлебопекарных качеств сортов пшеницы, создание высокоолеиновых сортов подсолнечника. 5. Создание высокопродуктивных сортов пшеницы. 6. Получение безалкалоидных сортов кормового люпина с увеличенным количеством бобов на растении и высокой семенной продуктивностью.
 Достижения мутационной селекции в Сибири Отбором из мутантной популяции сорта Мироновская 808, полученной с помощью ЭИ (0, 01%), создан сорт Омская озимая (1989). Сорт озимой пшеницы Сибирская Нива (1992) создан путём воздействия ЭИ (0, 1%) на сорт ППГ 186. Путём многократного индивидуального отбора из мутантных популяций созданы такие сорта яровой мягкой пшеницы, как Росинка (1997) – путем обработки Сибаковской 3 гамма-лучами; Росинка 2 (1999) – обработка Целинной 21 ЭИ; Славянка Сибири (2002) – обработка Лютесценс 65 НДММ. У позднеспелого высокоурожайного сорта Лютесценс 65 получено свыше 200 мутантов, которые отличаются сокращённым вегетационным периодом (до 5– 10 сут).
Достижения мутационной селекции в Сибири Отбором из мутантной популяции сорта Мироновская 808, полученной с помощью ЭИ (0, 01%), создан сорт Омская озимая (1989). Сорт озимой пшеницы Сибирская Нива (1992) создан путём воздействия ЭИ (0, 1%) на сорт ППГ 186. Путём многократного индивидуального отбора из мутантных популяций созданы такие сорта яровой мягкой пшеницы, как Росинка (1997) – путем обработки Сибаковской 3 гамма-лучами; Росинка 2 (1999) – обработка Целинной 21 ЭИ; Славянка Сибири (2002) – обработка Лютесценс 65 НДММ. У позднеспелого высокоурожайного сорта Лютесценс 65 получено свыше 200 мутантов, которые отличаются сокращённым вегетационным периодом (до 5– 10 сут).
 Лекция 9. Экспериментальная полиплоидия и гаплоидия Безвременник осенний – растение из семян и клубнелуковиц которого получают колхицин
Лекция 9. Экспериментальная полиплоидия и гаплоидия Безвременник осенний – растение из семян и клубнелуковиц которого получают колхицин
 Задачи экспериментальной полиплоидии в селекции 1. Повышение продуктивности. 2. Преодоление самонесовместимости. 3. Преодоление межродовой и межвидовой нескрещиваемости. 4. Восстановление плодовитости у отдалённых гибридов. 5. Закрепление гетерозиса.
Задачи экспериментальной полиплоидии в селекции 1. Повышение продуктивности. 2. Преодоление самонесовместимости. 3. Преодоление межродовой и межвидовой нескрещиваемости. 4. Восстановление плодовитости у отдалённых гибридов. 5. Закрепление гетерозиса.
 Требования к объектам полиплоидии 1. Растения с небольшим числом хромосом лучше реагируют на их удвоение, чем растения с высоким числом хромосом. 2. Перекрёстноопыляющиеся культуры более подходят для селекции на основе полиплоидии, чем самоопылители. У них скорее восстанавливается фертильность. 3. Чем более гетерозиготен материал, тем больше шансов получить выгодную новую комбинацию. Поэтому в качестве исходного материала для полиплоидизации целесообразно брать гибридный материал. 4. Растения, у которых используются вегетативные части, представляют более перспективный материал для полиплоидии, чем выращиваемые на семена.
Требования к объектам полиплоидии 1. Растения с небольшим числом хромосом лучше реагируют на их удвоение, чем растения с высоким числом хромосом. 2. Перекрёстноопыляющиеся культуры более подходят для селекции на основе полиплоидии, чем самоопылители. У них скорее восстанавливается фертильность. 3. Чем более гетерозиготен материал, тем больше шансов получить выгодную новую комбинацию. Поэтому в качестве исходного материала для полиплоидизации целесообразно брать гибридный материал. 4. Растения, у которых используются вегетативные части, представляют более перспективный материал для полиплоидии, чем выращиваемые на семена.
 Типы полиплоидов, используемые в селекции и методы их получения 1. Аллополиплоиды – возникают в результате кратного увеличения в клетках наборов хромосом одного и того же вида. АА АААА 2. Аутополиплоиды – получаются путём соединения в одном геноме хромосомных наборов разных видов и последующим удвоением числа хромосом или скрещиванием аутополиплоидов. 1) АА х ВВ = F 1 АВ ААВВ 2) АААА х ВВВВ = F 1 ААВВ 3. Сегментные аллополиплоиды. 4. Автоаллополиплоиды.
Типы полиплоидов, используемые в селекции и методы их получения 1. Аллополиплоиды – возникают в результате кратного увеличения в клетках наборов хромосом одного и того же вида. АА АААА 2. Аутополиплоиды – получаются путём соединения в одном геноме хромосомных наборов разных видов и последующим удвоением числа хромосом или скрещиванием аутополиплоидов. 1) АА х ВВ = F 1 АВ ААВВ 2) АААА х ВВВВ = F 1 ААВВ 3. Сегментные аллополиплоиды. 4. Автоаллополиплоиды.
 Происхождение различных типов полиплоидов
Происхождение различных типов полиплоидов
 Различия между аутополиплоидами и аллополиплоидами 1. Фертильность наиболее сильно снижается у аутополиплоидов, что связано с нарушениями процесса мейоза, особенно у культур, возделываемых на семена. У аллополиплоидов хромосомы каждого типа представлены парами, поэтому мейоз протекает нормально и плодовитость снижается меньше, чем у аутополиплоидов. 2. Морфологически аутополиплоид сходен с родительской формой, а аллополиплоид занимает промежуточное положение между родительскими видами и похож на гибрид между ними.
Различия между аутополиплоидами и аллополиплоидами 1. Фертильность наиболее сильно снижается у аутополиплоидов, что связано с нарушениями процесса мейоза, особенно у культур, возделываемых на семена. У аллополиплоидов хромосомы каждого типа представлены парами, поэтому мейоз протекает нормально и плодовитость снижается меньше, чем у аутополиплоидов. 2. Морфологически аутополиплоид сходен с родительской формой, а аллополиплоид занимает промежуточное положение между родительскими видами и похож на гибрид между ними.
 Получение капустно-редечного гибрида
Получение капустно-редечного гибрида
 Зерновые культуры у которых используются искусственно полученные тетраплоидные сорта 1. Рожь (Secale cereale L. )(4 x=28) 2. Гречиха (Fagopyrum esculentum ssp. vulgare St. )(4 x=32) Кормовые культуры 1. Кормовая свёкла (Beta vulgaris L. )(4 х=36) 2. Турнепс (Brassica rapa ssp. rapifera) (4 х=40) 3. Люцерна синяя (Medicago sativa L. ), Люцерна изменчивая (Medicago varia T. Martyn. )(4 х=32) 4. Клевер луговой (Trifolium pratense) (4 х=28), клевер гибридный (Trifolium hybridum L. )(4 х=32)
Зерновые культуры у которых используются искусственно полученные тетраплоидные сорта 1. Рожь (Secale cereale L. )(4 x=28) 2. Гречиха (Fagopyrum esculentum ssp. vulgare St. )(4 x=32) Кормовые культуры 1. Кормовая свёкла (Beta vulgaris L. )(4 х=36) 2. Турнепс (Brassica rapa ssp. rapifera) (4 х=40) 3. Люцерна синяя (Medicago sativa L. ), Люцерна изменчивая (Medicago varia T. Martyn. )(4 х=32) 4. Клевер луговой (Trifolium pratense) (4 х=28), клевер гибридный (Trifolium hybridum L. )(4 х=32)

 и три полиплоидных колоса ржи") Диплоидный (слева) и три полиплоидных колоса ржи
Диплоидный (слева) и три полиплоидных колоса ржи
 Проростки ржи: а – диплоидный; б – тетраплоидный.
Проростки ржи: а – диплоидный; б – тетраплоидный.
 Плоды гречихи: а – дипоидной, б – тетраплоидной.
Плоды гречихи: а – дипоидной, б – тетраплоидной.
") Клевер луговой (красный)
Клевер луговой (красный)
 Клевер гибридный
Клевер гибридный
 и тетраплоидного (справа) клевера") Соцветие диплоидного (слева) и тетраплоидного (справа) клевера
Соцветие диплоидного (слева) и тетраплоидного (справа) клевера

 Турнепс
Турнепс
 Люцерна синяя
Люцерна синяя
 Люцерна изменчивая
Люцерна изменчивая
 Положительные признаки аутотетраплоидов зерновых и кормовых культур 1. Повышенная продуктивность. 2. Устойчивость к полеганию за счёт прочного и толстого стебля. 3. Устойчивость к осыпанию (гречиха). 4. Устойчивость к болезням (рожь, свёкла) 5. Дружное созревание плодов (гречиха). 6. Крупные семена. 7. Повышенное содержание белка в зерне (гречиха). 8. Повышенный урожай зелёной массы (на 25 -86%) и быстрое отрастание после укоса (клевер).
Положительные признаки аутотетраплоидов зерновых и кормовых культур 1. Повышенная продуктивность. 2. Устойчивость к полеганию за счёт прочного и толстого стебля. 3. Устойчивость к осыпанию (гречиха). 4. Устойчивость к болезням (рожь, свёкла) 5. Дружное созревание плодов (гречиха). 6. Крупные семена. 7. Повышенное содержание белка в зерне (гречиха). 8. Повышенный урожай зелёной массы (на 25 -86%) и быстрое отрастание после укоса (клевер).
. 3. Большое содержание воды") Нежелательные признаки аутополиплоидов 1. Позднеспелость. 2. Снижение плодовитости (семенной продуктивности). 3. Большое содержание воды в вегетативной массе. 4. Более сложный характер наследования признаков по сравнению с диплоидами (расщепление в F 2 при моногибридном скрещивании – 35 : 1, вместо 3 : 1 – у диплоидов).
Нежелательные признаки аутополиплоидов 1. Позднеспелость. 2. Снижение плодовитости (семенной продуктивности). 3. Большое содержание воды в вегетативной массе. 4. Более сложный характер наследования признаков по сравнению с диплоидами (расщепление в F 2 при моногибридном скрещивании – 35 : 1, вместо 3 : 1 – у диплоидов).
 Методы создания тетраплоидных сортов ржи в Сиб. НИИСХ 1. Проводят гибридизацию тетраплоидных форм разного географического происхождения и последующий отбор. В качестве материнских форм используют сорта – Свеча, Белта, Пуховчанка, Шатиловская тетра и др. В качестве отцовских форм (тестера) – Тетра гибрид, Тетра короткая, Популяция 2. Например, сорт Сибирь (1999) создан в результате скрещивания трёх тетраплоидных сортов: местная репродукция Белта, Тетра короткая, Шатиловская тетра. 2. Создают тетраплоидные формы ржи на основе диплоидных гибридов первого поколения с целью закрепления гетерозисного эффекта переводом гибридов на полиплоидный уровень. Для удвоения хромосом у гибридов проводят обработку семян, проростков или колосьев раствором колхицина.
Методы создания тетраплоидных сортов ржи в Сиб. НИИСХ 1. Проводят гибридизацию тетраплоидных форм разного географического происхождения и последующий отбор. В качестве материнских форм используют сорта – Свеча, Белта, Пуховчанка, Шатиловская тетра и др. В качестве отцовских форм (тестера) – Тетра гибрид, Тетра короткая, Популяция 2. Например, сорт Сибирь (1999) создан в результате скрещивания трёх тетраплоидных сортов: местная репродукция Белта, Тетра короткая, Шатиловская тетра. 2. Создают тетраплоидные формы ржи на основе диплоидных гибридов первого поколения с целью закрепления гетерозисного эффекта переводом гибридов на полиплоидный уровень. Для удвоения хромосом у гибридов проводят обработку семян, проростков или колосьев раствором колхицина.
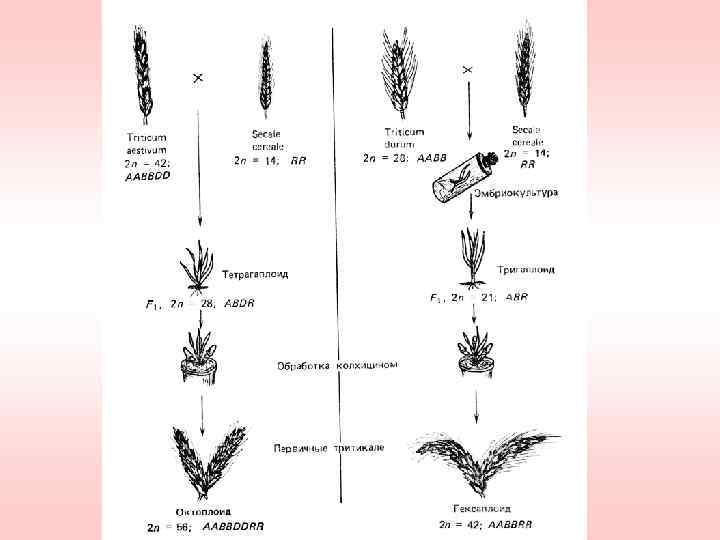
(2 n=56,") Аллополиплоиды, имеющие практическую ценность в растениеводстве 1. Тритикале (×Triticosecale Wittm. & A. Camus)(2 n=56, 42) 2. Рапс - В. napus (2 n=38)(листовая капуста-B. oleracea (4 х=36) х сурепица-B. campestris (4 х=40) ) 3. Брюква - В. napus (2 n=38)(листовая капуста. B. oleracea (4 х=36) х турнепс- B. campestris(4 х=40)) 4. Горчица сарептская – B. juncea(2 n=36) (горчица чёрная-B. nigra (4 х=32) х сурепица-B. campestris (4 х=40) )
Аллополиплоиды, имеющие практическую ценность в растениеводстве 1. Тритикале (×Triticosecale Wittm. & A. Camus)(2 n=56, 42) 2. Рапс - В. napus (2 n=38)(листовая капуста-B. oleracea (4 х=36) х сурепица-B. campestris (4 х=40) ) 3. Брюква - В. napus (2 n=38)(листовая капуста. B. oleracea (4 х=36) х турнепс- B. campestris(4 х=40)) 4. Горчица сарептская – B. juncea(2 n=36) (горчица чёрная-B. nigra (4 х=32) х сурепица-B. campestris (4 х=40) )
 Тритикале
Тритикале
 Родственные связи видов рода Brassica L.
Родственные связи видов рода Brassica L.
 Рапс
Рапс
 Горчица сарептская
Горчица сарептская
 Соцветие и корнеплод брюквы
Соцветие и корнеплод брюквы
 Создание триплоидных гибридов сахарной свёклы Триплоидные гибриды имеют большее содержание сахара в соке, большую массу корня и на 15 -20% больше выход сахара с единицы площади по сравнению с диплоидами, устойчивы к церкоспорозу. Для получения гибридных семян скрещивают диплоидные и тетраплоидные формы. Семенники диплоидов и тетраплоидов высаживают чередующимися рядами, и в результате естественного переопыления образуются гибридные триплоидные семена. Наиболее рентабельно размещение диплоидных и тетраплоидных форм в отношении 1 : 3 Например, 1 диплоидная многосемянная : 3 тетраплоидная односемянная = гибрид Белоцерковский поли 41
Создание триплоидных гибридов сахарной свёклы Триплоидные гибриды имеют большее содержание сахара в соке, большую массу корня и на 15 -20% больше выход сахара с единицы площади по сравнению с диплоидами, устойчивы к церкоспорозу. Для получения гибридных семян скрещивают диплоидные и тетраплоидные формы. Семенники диплоидов и тетраплоидов высаживают чередующимися рядами, и в результате естественного переопыления образуются гибридные триплоидные семена. Наиболее рентабельно размещение диплоидных и тетраплоидных форм в отношении 1 : 3 Например, 1 диплоидная многосемянная : 3 тетраплоидная односемянная = гибрид Белоцерковский поли 41
 Поскольку пыльцевые трубки диплоидных форм растут быстрее, чем тетраплоидных, то наблюдается тенденция к оплодотворению пыльцой диплоидных растений, поэтому на тетраплоидных растениях будут преобладать триплоидные семена. Большой практический интерес представляют гибриды между тетраплоидной сахарной и диплоидной кормовой свёклой, у которых повышен выход сахара с единицы площади (например, гибрид Тимирязевский 12).
Поскольку пыльцевые трубки диплоидных форм растут быстрее, чем тетраплоидных, то наблюдается тенденция к оплодотворению пыльцой диплоидных растений, поэтому на тетраплоидных растениях будут преобладать триплоидные семена. Большой практический интерес представляют гибриды между тетраплоидной сахарной и диплоидной кормовой свёклой, у которых повышен выход сахара с единицы площади (например, гибрид Тимирязевский 12).
 Триплоидный гибрид сахарной свёклы
Триплоидный гибрид сахарной свёклы
 Факторы, вызывающие полиплоидию 1. Физические – воздействия высокими температурами, ионизирующие излучения. 2. Механические – повреждения тканей, декапитация, центрифугирование. 3. Химические – колхицин, аппиоль, ауранция, закись азота, аценафтен, хлористый сангуинарин, гаммексан, линдан. 4. Генетические – использование генов, вызывающих образование диплоидных яйцеклеток (рецессивный ген elongate у кукурузы в гомозиготном состоянии вызывает образование диплоидных яйцеклеток), получение полиплоидов в результате отдалённых скрещиваний.
Факторы, вызывающие полиплоидию 1. Физические – воздействия высокими температурами, ионизирующие излучения. 2. Механические – повреждения тканей, декапитация, центрифугирование. 3. Химические – колхицин, аппиоль, ауранция, закись азота, аценафтен, хлористый сангуинарин, гаммексан, линдан. 4. Генетические – использование генов, вызывающих образование диплоидных яйцеклеток (рецессивный ген elongate у кукурузы в гомозиготном состоянии вызывает образование диплоидных яйцеклеток), получение полиплоидов в результате отдалённых скрещиваний.
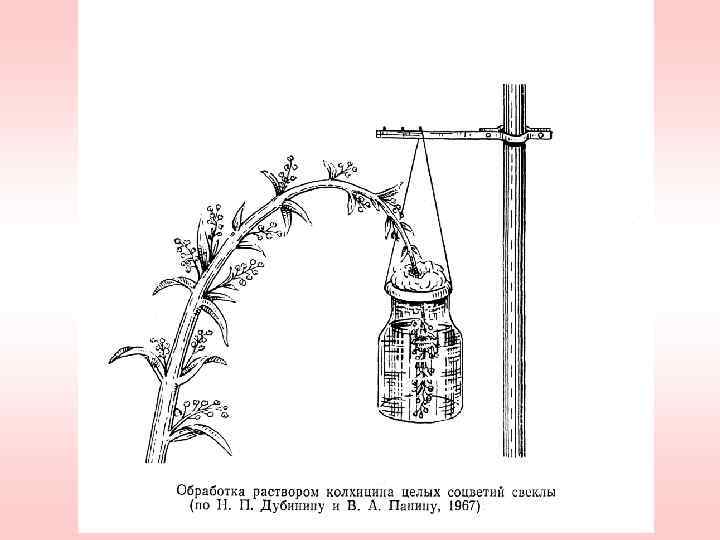
 используют") Использование колхицина для получения полиплоидов Колхицин (С 22 H 25 О 6 N) используют в виде водных растворов, ланолиновой пасты, в виде раствора в агар-агаре, глицерине, трагаканте. Готовят 1 -2%-ный раствор, а затем разбавляют до нужной концентрации. На свету колхицин разлагается. При обработке семян, проростков и корневой системы используют 0, 01 -0, 2%-ные растворы, а при воздейстии на стеблевые точки роста – 0, 52%-ные растворы. Продолжительность экспозиции варьирует от нескольких часов до нескольких суток.
Использование колхицина для получения полиплоидов Колхицин (С 22 H 25 О 6 N) используют в виде водных растворов, ланолиновой пасты, в виде раствора в агар-агаре, глицерине, трагаканте. Готовят 1 -2%-ный раствор, а затем разбавляют до нужной концентрации. На свету колхицин разлагается. При обработке семян, проростков и корневой системы используют 0, 01 -0, 2%-ные растворы, а при воздейстии на стеблевые точки роста – 0, 52%-ные растворы. Продолжительность экспозиции варьирует от нескольких часов до нескольких суток.
 Факторы влияющие на чувствительность к колхицину и условия, повышающие эффективность его действия Чувствительность к колхицину зависит от 1) периода развития культуры, 2) зоны меристемы (обрабатывать желательно меристематические ткани с максимальным количеством делящихся клеток), 3) обрабатываемых органов (корни более чувствительны, чем стеблевые точки роста), от температуры. Обработка семян и проростков колхицином более эффективна в условиях вакуума, при использовании переносчиков колхицина (диметилсульфооксид) и в комбинации со стимуляторами роста (гетероауксином). Температура во время обработки варьирует от 19 -21 (для ячменя) до 27 -30 градусов.
Факторы влияющие на чувствительность к колхицину и условия, повышающие эффективность его действия Чувствительность к колхицину зависит от 1) периода развития культуры, 2) зоны меристемы (обрабатывать желательно меристематические ткани с максимальным количеством делящихся клеток), 3) обрабатываемых органов (корни более чувствительны, чем стеблевые точки роста), от температуры. Обработка семян и проростков колхицином более эффективна в условиях вакуума, при использовании переносчиков колхицина (диметилсульфооксид) и в комбинации со стимуляторами роста (гетероауксином). Температура во время обработки варьирует от 19 -21 (для ячменя) до 27 -30 градусов.
 Методы получения полиплоидов с помощью колхицина 1. Колхицинирование семян. 2. Погружение проростков в водный раствор колхицина. 3. Обработка мелкосемянных культур. 4. Капельный метод. 5. Метод инъекций. 6. Метод CIMMYT. 7. Обработка корней. 8. Обработка взрослых растений. 9. Обработка путём погружения побега. 10. Обработка цветоносных побегов.
Методы получения полиплоидов с помощью колхицина 1. Колхицинирование семян. 2. Погружение проростков в водный раствор колхицина. 3. Обработка мелкосемянных культур. 4. Капельный метод. 5. Метод инъекций. 6. Метод CIMMYT. 7. Обработка корней. 8. Обработка взрослых растений. 9. Обработка путём погружения побега. 10. Обработка цветоносных побегов.
 Капельный метод
Капельный метод
 и в") Обработка проростков колхицином. Семена помещают на фильтровальную бумагу в чашки Петри (1) и в момент их наклёвывания чашки переворачивают дном вверх с тем. Чтобы отрастающие корешки в силу геотропизма росли вниз (2). При длине корешков около 0, 5 мм чашки возвращают в исходное положение, семена заливают раствором колхицина, корешки при этом не должны соприкасаться с раствором (3). Более крупные семена при переворачивании чашки придерживают металлической сеткой.
Обработка проростков колхицином. Семена помещают на фильтровальную бумагу в чашки Петри (1) и в момент их наклёвывания чашки переворачивают дном вверх с тем. Чтобы отрастающие корешки в силу геотропизма росли вниз (2). При длине корешков около 0, 5 мм чашки возвращают в исходное положение, семена заливают раствором колхицина, корешки при этом не должны соприкасаться с раствором (3). Более крупные семена при переворачивании чашки придерживают металлической сеткой.

 Морфологические и анатомические признаки полиплоидизации растения 1. Увеличены размеры вегетативных органов и изменена их форма. 2. Растения увеличиваются в ширину и уменьшаются в высоту. 3. Листья крупнее, шире и толще, имеют неправильную форму, бугристость и кожистость, окраска интенсивно зелёная. 4. Увеличена лигула – язычок у злаков. 5. Семядоли у проростков более крупные, мясистые, корешки отличаются булавовидными вздутиями, более короткий раздутый стебель. 6. Значительно увеличены размеры цветков и целых соцветий. 7. Возрастают размеры пыльцевых зёрен, а также плодов и семян.
Морфологические и анатомические признаки полиплоидизации растения 1. Увеличены размеры вегетативных органов и изменена их форма. 2. Растения увеличиваются в ширину и уменьшаются в высоту. 3. Листья крупнее, шире и толще, имеют неправильную форму, бугристость и кожистость, окраска интенсивно зелёная. 4. Увеличена лигула – язычок у злаков. 5. Семядоли у проростков более крупные, мясистые, корешки отличаются булавовидными вздутиями, более короткий раздутый стебель. 6. Значительно увеличены размеры цветков и целых соцветий. 7. Возрастают размеры пыльцевых зёрен, а также плодов и семян.
 8. Увеличены клетки эпидермиса и замыкающие клетки устьиц. 9. Увеличено количество пор в экзине (оболочке пыльцевых зёрен).
8. Увеличены клетки эпидермиса и замыкающие клетки устьиц. 9. Увеличено количество пор в экзине (оболочке пыльцевых зёрен).
 Микроскоп для подсчёта числа хромосом, изучения пыльцы и устьичного аппарата у полиплоидов
Микроскоп для подсчёта числа хромосом, изучения пыльцы и устьичного аппарата у полиплоидов
 и тетраплоидных (справа) растений сахарной свёклы сорта Рамонская 023") Пыльца диплоидных (слева) и тетраплоидных (справа) растений сахарной свёклы сорта Рамонская 023
Пыльца диплоидных (слева) и тетраплоидных (справа) растений сахарной свёклы сорта Рамонская 023
 и тетраплоидный (2 n=4 x=32) наборы хромосом у гречихи") Диплоидный (2 n=16) и тетраплоидный (2 n=4 x=32) наборы хромосом у гречихи
Диплоидный (2 n=16) и тетраплоидный (2 n=4 x=32) наборы хромосом у гречихи
; Б") Близнецовые растения из одной семяпочки: А - спонтанный тетраплоид сахарной свёклы (4 х=36); Б - диплоидное растение (2 х=18)
Близнецовые растения из одной семяпочки: А - спонтанный тетраплоид сахарной свёклы (4 х=36); Б - диплоидное растение (2 х=18)
 Замыкающие клетки устьиц и хлоропласты растений шелковицы: 1 – диплоидной, 2 – триплоидной, 3 – тетраплоидной.
Замыкающие клетки устьиц и хлоропласты растений шелковицы: 1 – диплоидной, 2 – триплоидной, 3 – тетраплоидной.
 Схема создания тетраплоидного сорта гречихи Год Этап селекционного процесса 1 -й Сорт А х Сорт В F 0 2 -й F 1 (Гибриды) оценка гетерозиса 3 -й Полиплоидизация гибридов (колхицинирование стеблевых точек роста проростков) F 1 C 0, отбор растений по числу пор в экзине и числу хромосом. 4 -й Отбор тетраплоидных растений по числу хромосом в С 1 и нормально протекающему мейозу в период бутонизации. Браковка растений с низкой семенной продуктивностью. 5 -й Выделенные в С 1 в период уборки высокопродуктивные растения высевают в С 2 на изолированных участках, проводят негативный отбор в пределах каждого потомства до цветения, в период уборки отбирают высокопродуктивные растения в высокоурожайных потомствах С 2.
Схема создания тетраплоидного сорта гречихи Год Этап селекционного процесса 1 -й Сорт А х Сорт В F 0 2 -й F 1 (Гибриды) оценка гетерозиса 3 -й Полиплоидизация гибридов (колхицинирование стеблевых точек роста проростков) F 1 C 0, отбор растений по числу пор в экзине и числу хромосом. 4 -й Отбор тетраплоидных растений по числу хромосом в С 1 и нормально протекающему мейозу в период бутонизации. Браковка растений с низкой семенной продуктивностью. 5 -й Выделенные в С 1 в период уборки высокопродуктивные растения высевают в С 2 на изолированных участках, проводят негативный отбор в пределах каждого потомства до цветения, в период уборки отбирают высокопродуктивные растения в высокоурожайных потомствах С 2.
 6 -й Высокопродуктианые растения С 2 объединяют в пределах каждого потомства и высевают на С 3 смежно с другими потомствами в питомнике направленного переопыления. До цветения удаляют слаборазвитые растения, а в период уборки отбирают высокопродуктивные. После анализа структуры урожая и качества зерна, потомства объединяют в популяцию. 7 -9 -й Сформированные таким образом популяции включают в испытательные питомники. Предварительное сортоиспытание С 4 -С 6 10 -12 Государственное сортоиспытание С 7 -С 9 -й 13 -й Районирование С 10 C 0 – растения, подвергшиеся колхицинированию. Образующиеся семена и полученные из них растения обозначают С 1; последующие поколения – С 2, С 3, С 4 и т. д.
6 -й Высокопродуктианые растения С 2 объединяют в пределах каждого потомства и высевают на С 3 смежно с другими потомствами в питомнике направленного переопыления. До цветения удаляют слаборазвитые растения, а в период уборки отбирают высокопродуктивные. После анализа структуры урожая и качества зерна, потомства объединяют в популяцию. 7 -9 -й Сформированные таким образом популяции включают в испытательные питомники. Предварительное сортоиспытание С 4 -С 6 10 -12 Государственное сортоиспытание С 7 -С 9 -й 13 -й Районирование С 10 C 0 – растения, подвергшиеся колхицинированию. Образующиеся семена и полученные из них растения обозначают С 1; последующие поколения – С 2, С 3, С 4 и т. д.
 Направления использования экспериментальной гаплоидии в селекции 1. Ускоренное получение гомозиготных линий для производства гетерозисных гибридов F 1. 2. Быстрый перевод самоопылённых линий кукурузы на стерильную основу. 3. Ускорение селекционного процесса у самоопыляющихся культур. 4. Увеличение вероятности отбора ценных комбинаций признаков у дигаплоидов гибридов F 2. 5. Интрогрессия отдельных признаков от диких тетраплоидных видов культурным, путём создания дигаплоидов и их скрещивания между собой.
Направления использования экспериментальной гаплоидии в селекции 1. Ускоренное получение гомозиготных линий для производства гетерозисных гибридов F 1. 2. Быстрый перевод самоопылённых линий кукурузы на стерильную основу. 3. Ускорение селекционного процесса у самоопыляющихся культур. 4. Увеличение вероятности отбора ценных комбинаций признаков у дигаплоидов гибридов F 2. 5. Интрогрессия отдельных признаков от диких тетраплоидных видов культурным, путём создания дигаплоидов и их скрещивания между собой.
 У кукурузы установлен андрогенез, когда после слияния ядра яйцеклетки с ядром спермия материнское ядро элиминируется и зародыш развивается только из ядра спермия; поэтому он гаплоиден и передает отцовские признаки.
У кукурузы установлен андрогенез, когда после слияния ядра яйцеклетки с ядром спермия материнское ядро элиминируется и зародыш развивается только из ядра спермия; поэтому он гаплоиден и передает отцовские признаки.
 Методы получения гаплоидов 1 метод. Отдалённая гибридизация - при отдаленной гибридизации некоторых видов установлено явление селективной элиминации хромосом одного из родителей на ранней стадии развития гибридного зародыша. Например, при скрещивании диплоидных ячменей Hordeum vulgare (культурный) и H. bulbosum (многолетний луковичный дикий) на стадии роста зародыша и эндосперма (через 5 дней после оплодотворения) происходит выпад хромосом дикого вида. Возникает гаплоид с набором хромосом H. vulgare. Через 15 суток после оплодотворения рост гибридного зародыша на материнском растении прекращается, но при культивировании in vitro из таких зародышей развиваются проростки.
Методы получения гаплоидов 1 метод. Отдалённая гибридизация - при отдаленной гибридизации некоторых видов установлено явление селективной элиминации хромосом одного из родителей на ранней стадии развития гибридного зародыша. Например, при скрещивании диплоидных ячменей Hordeum vulgare (культурный) и H. bulbosum (многолетний луковичный дикий) на стадии роста зародыша и эндосперма (через 5 дней после оплодотворения) происходит выпад хромосом дикого вида. Возникает гаплоид с набором хромосом H. vulgare. Через 15 суток после оплодотворения рост гибридного зародыша на материнском растении прекращается, но при культивировании in vitro из таких зародышей развиваются проростки.
 Частота и количество образовавшихся растений при этом способе очень высоки. Кроме того, растения-альбиносы не образуются. С помощью этого метода были выведены сорта Исток и Одесский-115 – за 4 года вместо 10 - 12 лет обычной селекции. В Канаде так были получены сорта Минго и Родео. Если в качестве опылителя использовать дикий ячмень, то можно индуцировать гаплоиды у ржи и пшеницы.
Частота и количество образовавшихся растений при этом способе очень высоки. Кроме того, растения-альбиносы не образуются. С помощью этого метода были выведены сорта Исток и Одесский-115 – за 4 года вместо 10 - 12 лет обычной селекции. В Канаде так были получены сорта Минго и Родео. Если в качестве опылителя использовать дикий ячмень, то можно индуцировать гаплоиды у ржи и пшеницы.
 2 метод. Выращивание гаплоидов на питательной среде из пыльников или пыльцы - из неоплодотворенных половых клеток с редуцированным набором хромосом можно регенерировать целые растения. Получение гаплоидных растений из изолированных пыльников может идти по двум направлениям: 1. Прямая регенерация соматических зародышей внутри пыльников из отдельных пыльцевых зерен формируются проэмбриональные структуры, которые при определенных условиях культивирования развиваются в эмбриоиды (зародышеподобные структуры), дающие начало гаплоидным растениям. 2. Косвенная - через каллусогенез - пыльца делится, клетки, возникшие в результате делений, быстро увеличиваются в размерах и, разрывая оболочку пыльцевого зерна, образуют каллус.
2 метод. Выращивание гаплоидов на питательной среде из пыльников или пыльцы - из неоплодотворенных половых клеток с редуцированным набором хромосом можно регенерировать целые растения. Получение гаплоидных растений из изолированных пыльников может идти по двум направлениям: 1. Прямая регенерация соматических зародышей внутри пыльников из отдельных пыльцевых зерен формируются проэмбриональные структуры, которые при определенных условиях культивирования развиваются в эмбриоиды (зародышеподобные структуры), дающие начало гаплоидным растениям. 2. Косвенная - через каллусогенез - пыльца делится, клетки, возникшие в результате делений, быстро увеличиваются в размерах и, разрывая оболочку пыльцевого зерна, образуют каллус.
 В результате дальнейшего морфогенеза из этих каллусных клеток регенерируют растения. При этом растения могут иметь разную степень плоидности - ди-, поли-, анеуплоидные. Последние часто стерильны, но после обработки растений колхицином происходит удвоение числа хромосом, в результате чего можно получить фертильные гомозиготы.
В результате дальнейшего морфогенеза из этих каллусных клеток регенерируют растения. При этом растения могут иметь разную степень плоидности - ди-, поли-, анеуплоидные. Последние часто стерильны, но после обработки растений колхицином происходит удвоение числа хромосом, в результате чего можно получить фертильные гомозиготы.
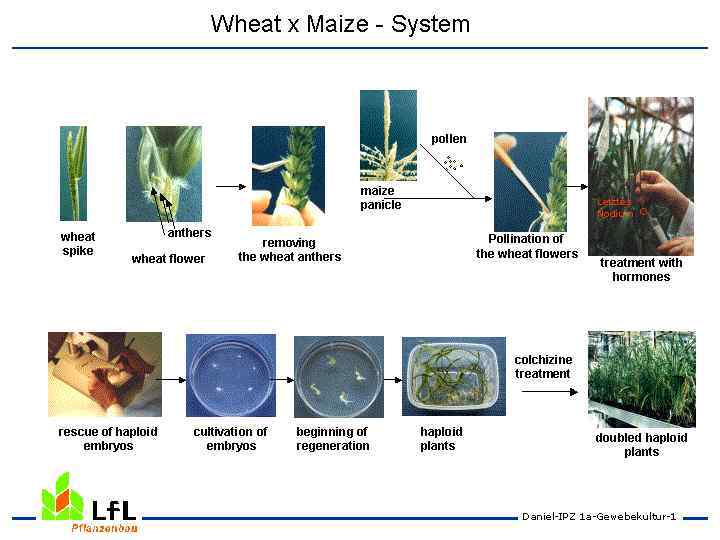
 Выращивание гаплоидов из пыльцевых зёрен
Выращивание гаплоидов из пыльцевых зёрен
 и дикого (справа) вида картофеля") Дигаплоиды культурного (слева) и дикого (справа) вида картофеля
Дигаплоиды культурного (слева) и дикого (справа) вида картофеля
 для вовлечения в селекцию") А. П. Ермишин, О. В. Маханько, Е. В. Воронкова (2005) для вовлечения в селекцию ценного генофонда диких диплоидных видов картофеля из Мексики получили дигаплоиды (2 n = 2 x = 24) тетраплоидных соматических гибридов (2 n = 4 x = 48) и провели последующее их беккроссирование дигаплоидами Solanum tuberosum. С помощью гаплопродюсера S. phureja Iv. P 35 было получено десять дигаплоидов соматических гибридов S. tuberosum + S. pinnatisectum, которые все скрещивались с фертильными дигаплоидами S. tuberosum с образованием выполненных жизнеспособных семян. Это дает возможность проведения эффективной интрогрессии селекционно-ценных генов дикого вида в селекционный материал на диплоидном уровне, что имеет ряд преимуществ по сравнению с осуществлением этой процедуры на тетраплоидном уровне.
А. П. Ермишин, О. В. Маханько, Е. В. Воронкова (2005) для вовлечения в селекцию ценного генофонда диких диплоидных видов картофеля из Мексики получили дигаплоиды (2 n = 2 x = 24) тетраплоидных соматических гибридов (2 n = 4 x = 48) и провели последующее их беккроссирование дигаплоидами Solanum tuberosum. С помощью гаплопродюсера S. phureja Iv. P 35 было получено десять дигаплоидов соматических гибридов S. tuberosum + S. pinnatisectum, которые все скрещивались с фертильными дигаплоидами S. tuberosum с образованием выполненных жизнеспособных семян. Это дает возможность проведения эффективной интрогрессии селекционно-ценных генов дикого вида в селекционный материал на диплоидном уровне, что имеет ряд преимуществ по сравнению с осуществлением этой процедуры на тетраплоидном уровне.
 Микрофотографии клеток корешка с метафазной пластинкой хромосом S. laplaticum (2 n=48 -слева, 2 n=24 – cправа)
Микрофотографии клеток корешка с метафазной пластинкой хромосом S. laplaticum (2 n=48 -слева, 2 n=24 – cправа)
 Выращивание гаплоидов на искусственной питательной среде
Выращивание гаплоидов на искусственной питательной среде
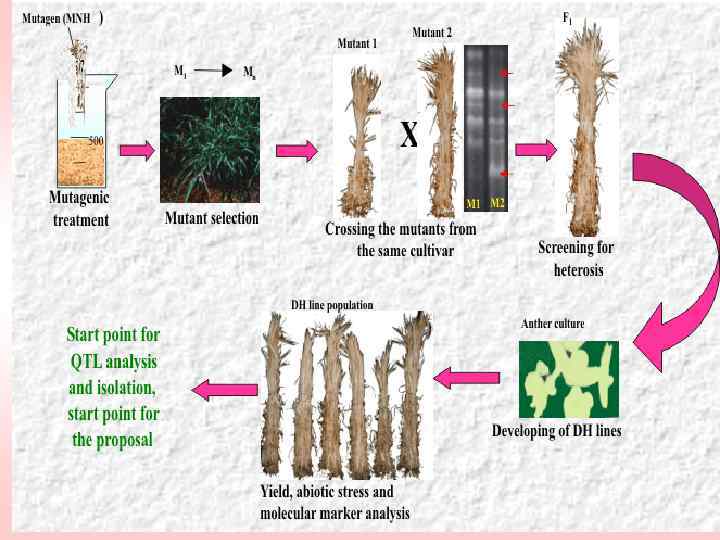
 Схема получения сорта ячменя БИОС-1 Лаборатория генетики растений ИОГен им. Н. И. Вавилова РАН
Схема получения сорта ячменя БИОС-1 Лаборатория генетики растений ИОГен им. Н. И. Вавилова РАН
 Технология получения гомозиготных линий злаков Технология разработана в лаборатории клеточной селекции ГНУ НИИСХ Юго-Востока. Руководитель д. б. н. Дьячук Т. И 1. Отбор донорных растений и культивирование пыльников на среде с 2, 4 - Д Сорт озимой тритикале Святозар 5. Размножение семян для последующего селекционного изучения 2. Получение гаплоидных новообразований (эмбриоидов и каллусов) 4. Удвоение числа хромосом колхицином у регенерантов 3. Регенерация гаплоидных растений ПРАКТИЧЕСКАЯ РЕАЛИЗАЦИЯ: создано более 15 тысяч гомозиготных линий пшеницы, ячменя, тритикале и проса. Создан допущенный к использованию сорт яровой мягкой пшеницы Саратовская 64. Передан на Государственное сортоиспытание сорт озимой гексаплоидной тритикале Святозар.
Технология получения гомозиготных линий злаков Технология разработана в лаборатории клеточной селекции ГНУ НИИСХ Юго-Востока. Руководитель д. б. н. Дьячук Т. И 1. Отбор донорных растений и культивирование пыльников на среде с 2, 4 - Д Сорт озимой тритикале Святозар 5. Размножение семян для последующего селекционного изучения 2. Получение гаплоидных новообразований (эмбриоидов и каллусов) 4. Удвоение числа хромосом колхицином у регенерантов 3. Регенерация гаплоидных растений ПРАКТИЧЕСКАЯ РЕАЛИЗАЦИЯ: создано более 15 тысяч гомозиготных линий пшеницы, ячменя, тритикале и проса. Создан допущенный к использованию сорт яровой мягкой пшеницы Саратовская 64. Передан на Государственное сортоиспытание сорт озимой гексаплоидной тритикале Святозар.