Лек 6 2014.ppt
- Количество слайдов: 51



Лекция 6 Взаимодействие клеток Система синтеза белка Пузырьковый транспорт

Плазматическая мембрана – граница клетки с окружающей средой Одна из функций клеточной мембраны – обеспечение взаимодействия клеток и формирование межклеточных контактов в тканях Взаимодействие клеток: Адгезия (прилипание, слипание) - селективный процесс, протекающий только между определенными типами клеток. В процессе адгезии во внеклеточном матриксе формируются примембранные структуры, образованные молекулами адгезии (рецепторы адгезии), которые взаимодействуют с прилежащими к плазмалемме белками цитоплазмы и цитоскелета. Селективная адгезия клеток обеспечивается трансмембранными белками - МОЛЕКУЛАМИ КЛЕТОЧНОЙ АДГЕЗИИ (cell adhesion molecules) Гомофильная и гетерофильная связь – между клетками одного, или разного типа. Формирование тканей и органов

Число молекул межклеточной адгезии очень велико. Молекулы клеточной адгезии в основном представлены членами следующих белковых суперсемейств: - иммуноглобулины - интегрины - лектины (в том числе - селектины) - протеогликаны -кадгерины. Выделяют кальций-зависимые и кальций-независимые типы адгезии. Кальций–независимая клеточная адгезия в основном обеспечивается иммуноглобулиноподобными белками, а также лектинами. Лектины - белки, быстро, избирательно и обратимо связывающиеся с сахарами.

Кадгерины – молекулы Са 2+ - зависимой адгезии Присутствуют практически во всех клетках позвоночных Трансмембранные гликопротеины Внутриклеточный домен связывается с белками цитоскелета Внеклеточный домен имеет Са-связывающий сайт Гомофильная связь: кадгерин одной клетки связывается с кадгерином другой клетки Морфологически оформленные межклеточные контакты
 Кадгерины В области и замыкающих комплексов, и десмосом мембраны")
Замыкающий комплекс (опоясывающий непрерывный контакт) Кадгерины В области и замыкающих комплексов, и десмосом мембраны не сливаются. Место контакта имеет особую структуру.
. Опоясывающий непрерывный контакт")
Замыкающие комплексы (пласты эпителия). Опоясывающий непрерывный контакт

Десмосома Дискретный контакт Кадгерины

Контакты клеток в эпителиоидной культуре MDCK

Эпителий почечного канальца лосося

Эпителий десны больного парадонтозом

В тканях: простой контакт – мембраны прилежат друг к другу и разделены пространством не менее 10 нм Нет дополнительных структур, межклеточное пространство заполнено гликокаликсом и внеклеточным матриксом, электроннопрозрачно Интердигитации в базальных отделах кишечного эпителия

Щелевидный контакт Коннексины Область протяженностью 0, 5 -3 мкм, где плазматические мембраны разделены промежутком 2 -3 нм, что придает структуре 7 -слойный вид после осмирования. Служат для прямой связи между цитоплазмой соседних клеток. Содержат специальные поровые структуры, переносящие молекулы до 1000 дальтон. В каждой клетке образуется полупора, полупоры соседних клеток формируют пору. Полупоры собираются в аппарате Гольджи и в составе пузырьков доставляются к плазмалемме.

Щелевидный контакт между клетками печени. Атомно-силовая микроскопия высокого разрешения Ультратонкий срез
")
Синапсы Специальный тип контактов между клетками нервной ткани Обеспечивают передачу нервного импульса (потенциала действия) с нервной клетки на другую нервную клетку.

Межклеточные контакты обеспечивают пространственную организацию тканей, поляризацию клеток, защиту внутренней среды организма. Опухолевые клетки не образуют межклеточныж контактов. Дестабилизация кадгерин-опосредованных контактов может быть обусловлена действием онкогенов, токсическими воздействиями, инфекционными агентами. Для стабильности межклеточных контактов важен кальций: в отсутствие кальция изменяется конформация кадгеринов и, в результате, они быстро разрушаются протеазами. 1 — простое межклеточное соединение; 2 — интердигитация; 3 — щелевидное соединение; 4 — замыкающий комплекс; 5 — десмосома

Система синтеза белка ДНК – матрица для синтеза и ДНК, и РНК - информационная (матричная), рибосомальная и транспортная. Транскрипция - перенос информации с ДНК на РНК, Или синтез и-РНК (м-РНК) в соответствии с ДНК-матрицей ТРАНСЛЯЦИЯ – синтез белков в соответствии с матрицей и-РНК Трансляция происходит на рибосомах Процесс трансляции очень консервативен Все РНК «читаются» с 5’-конца к 3’–концу Все белки синтезируются с аминогруппы к карбоксильному концу РИБОСОМА – «машина» для синтеза белка

Размеры рибосом: 20 - 24 нм Рибосома прокариот 50 s 30 s Рибосома эукариот 60 s 40 s Клетка E. Cоli содержит около 20 000 рибосом, Быстрорастущая клетка млекопитающих – около 10 000 рибосом

Нобелевская премия по химии 2009 г. Yonath A. E. , Steitz Th. A. , Ramakrishnan V. Схема строения бактериальной рибосомы. Рентгеноструктурный анализ. Молекулы р. РНК – оранжевого цвета Белки малой субъединицы – голубого цвета Белки большой субъединицы – зеленого Молекула антибиотика (красный цвет) связана с малой единицей. Фракция рибосом Напыление металлом

Схема синтеза белка на рибосомах Стадия элонгации Стадия терминации Полисома. Расстояние между рибосомами – 100– 200 нуклеотидов р. РНК обеспечивает сборку белковой цепи без участия белков. т. е, р. РНК обладает пептидил-трансферазной активностью Рибосома является рибозимом.
 РНК в ядре (м. РНК) 2 – соединение м.")
1 – синтез матричной (информационной) РНК в ядре (м. РНК) 2 – соединение м. РНК с субъединицами рибосомы 3 – транспортные РНК (т. РНК) соединяются с аминокислотами 4 – на рибосоме т. РНК, несущая АК, связывается с м. РНК (кодон и антикодон) 5 – формируется пептидная связь между АК – синтез белка
 и скоплением промежуточных")
Участок клетки с высоким содержанием свободных рибосом и полисом (красный контур) и скоплением промежуточных филаментов (синий контур)
 Гликозилирование –")
Визуализация синтеза белка на рибосомах Модификация синтезированных белковых молекул Протеолитическое расщепление (cleavage) Гликозилирование – модификация белков путем присоединения углеводов Модификация белков присоединением липидов

Первичная, вторичная, третичная и четвертичная структура белков Вторичная структура белков определяется водородными связями. Неупорядоченная спираль – при отсутствии водородных связей α спираль β слой Около 60% белков существуют в виде α- спирали, или β- слоя. Остальные – в виде неупорядоченных спиралей и изогнутых цепей в виде буквы U (шпилька).

Третичная структура белков. Определяется гидрофобными взаимодействиями между неполярными группами, и дисульфидными мостиками между аминокислотами. Третичная структура является наивысшим уровнем организации для мономерных белков, состоящих из одной цепи. Мультимерные белки состоят из двух, или более цепей, или субъединиц, соединенных нековалентными связями. Четвертичная структура белков. Методы изучения: - рентгеноструктурный анализ - электронно-микроскопическая томография Эпитопы – участки, на которые вырабатывается иммунный ответ. Кристаллы белков, выращенные в космосе
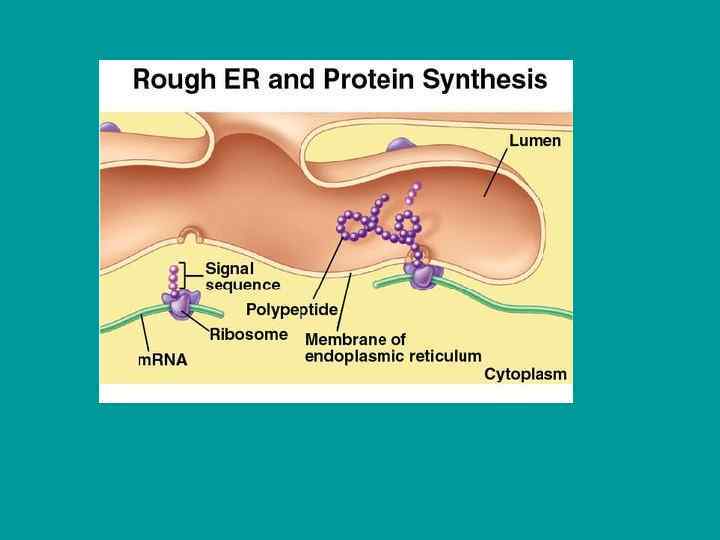
 Прикрепленные рибосомы (полисомы) Локализуются в цитозоле Локализуются на мембранах ЭПР Синтез")
Свободные рибосомы (полисомы) Прикрепленные рибосомы (полисомы) Локализуются в цитозоле Локализуются на мембранах ЭПР Синтез немембранных белков Синтез мембранных и немембранных белков ЭПР – эндоплазматический ретикулум = ЭПС – эндоплазматическая сеть
 (ЭПС, ЭПР) Расположена в цитоплазме. ЭПР – самая протяженная мембранная")
Эндоплазматическая сеть (эндоплазматический ретикулум) (ЭПС, ЭПР) Расположена в цитоплазме. ЭПР – самая протяженная мембранная структура клетки. Имеет единое замкнутое внутреннее пространство. Представляет собой сложную систему плоских и трубчатых элементов. Мембрана ЭПР – «обычная» трехслойная биологическая мембрана. В составе ЭПР выделяют часть, формирующую ядерную оболочку, структуры, локализованные в центре и на периферии клетки. ЭПР имеется у простейших, во всех клетках многоклеточных животных и растений Нет ЭПР в цитоплазме зрелых эритроцитов, в клетках сине-зеленых водорослей. Гладкий и шероховатый ЭПР - по наличию рибосом на мембране

Ширина просвета трубочек и цистерн ЭПР практически одинакова – 38 и 36 нм, соответственно Соотношение между тубулярной частью и цистернами зависит от типа клеток и их функционального состояния Оболочка ядра Ядро рибосомы Шероховатый ЭПР Гладкий ЭПР

Клетка Cos-7, экспрессирующая маркер мембран ЭПР - GFP-Sec 61 b. Видна ядерная оболочка и сеть структур, В - элементы ЭПР при большом увеличении.

Накопление пероксидазы хрена в ЭПР в процессе синтеза Видна ядерная оболочка. Ультратонкий срез

Шероховатый ЭПР

Шероховатый ЭПР

Гладкий ЭПР На мембране шероховатого ЭПР есть «гладкие» участки, это – не гладкий ЭПР. Гладкий ЭПР представлен трубочками, анастомозирующими между собой

ЭПР - один из универсальных клеточных органоидов выполняющих разносторонние функции. Главная функция шероховатого ЭПР - синтез и транслокация белков. Полость гладкого ЭПР является депо ионов кальция, а на её мембранах осуществляется синтез гликогена и липидов. ГЭПР развит в клетках, синтезирующих гликоген и липиды, например, в клетках сальных желез и коры надпочечника, в клетках печени, где происходит усиленный синтез гликогена. Появление новых методов визуализации клеточных структур заставляет критически анализировать данные о функциях ЭПР. Так, оказалось, что: -Нет доказательств разделения «классических» функций ШЭПС и ГЭПС. -Нет оснований говорить о разной роли трубочек и цистерн в жизни клетки. Очевидно, трубочки ЭПР– место образования транспортных везикул.

Почему ЭПР – единая структура, не распадающаяся на отдельные элементы даже во время деления клетки? Зачем нужны трубочки? Возможно, трубчатая форма обеспечивает проникновение элементов ЭПР в разные участки цитоплазмы. Новые данные: Установлены связи (контакты) ЭПР с другими мембранными органоидами и с плазмалеммой. Между мембраной ЭПР и мембраной органоида всегда сохраняется пространство около 30 нм. Слияния мембран разных органоидов нет. Контакты опосредуются особыми «привязывающими» белками (tethering proteins).
 ЭПР с другими мембранными органоидами и плазмалеммой")
Связь (контакт) ЭПР с другими мембранными органоидами и плазмалеммой

Предполагают, что тесная связь мембраны ЭПР и мембран различных органоидов обеспечивает прямой перенос молекул между структурами, в первую очередь – липидных. Считается, что контакт мембраны ЭПР и плазмалеммы обеспечивает перенос синтезированных в ЭПР сфинголипидов. В зоне контакта ЭПР и митохондрий реализуется, как минимум, две функции: синтез липидов и передача Ca 2+ -сигналов, Контакт мембраны ЭПР и других органоидов считают свидетельством «прямого» переноса молекул между органоидами, не связанного с формированием везикул. Механизмы пока не установлены.

Транспорт макромолекул в клетке: «пузырьковый» транспорт Клетка – сложная система взаимодействующих компартментов, ограниченных мембранами. Связь между мембранными компартментами реализуется посредством пузырькового транспорта. В одном пузырьке может переносится несколько разных лигандов. Пути переноса макромолекул в клетке с участием мембран: - формирование трубочек, которые «дорастают» до целевой мембраны (до 20% всего транспорта); - “Kiss-and-run” – пузырек-переносчик вступает в кратковременный контакт с целевой мембраной; - пузырек-переносчик сливается с целевой мембраной.
: А. формируются на")
Три типа «опушенных» пузырьков в клетке: 1. Сlathrin-coated vesicles (клатрин-опушенные пузырьки): А. формируются на плазмалемме Б. формируются на мембранах транс-сети АГ и переносят молекулы в лизосомы В. Формируются на мембранах эндосом 2 и 3. Non-clathrin-coated vesicles (COP-coated vesicles) 2. COP-I - пузырьки отпочковываются от АГ 3. COP-II - пузырьки отпочковываются от ЭПР. Могут иметь разные размеры (60 -90 нм), в зависимости от величины переносимой молекулы. COP - coat protein complex. Во всех типах пузырьков: Локализация молекул относительно мембраны неизменна. Есть сигнальные последовательности.

Фотосъемка живой клетки в течение двух минут Метка - СOP II Белок ts-O 45 -G – переносимая молекула

Основные стадии мембранного транспорта посредством «опушенных» везикул: 1. Почкование 2. Перемещение пузырька 3. «Причаливание» (связывание) с целевой мембраной 4. Слияние пузырька с целевой мембраной

Механизмы слияния мембранных пузырьков с «целевой» мембраной оценены Нобелевским комитетом Press Release 2013 -10 -07 The Nobel Assembly at Karolinska Institute has today decided to award The 2013 Nobel Prize in Physiology or Medicine jointly to James E. Rothman, Randy W. Schekman and Thomas C. Südhof for their discoveries of machinery regulating vesicle traffic, a major transport system in our cells
, локализованных в разных мембранах, в своеобразные")
Ключевой момент слияния мембран переплетение особых белков (SNARE), локализованных в разных мембранах, в своеобразные косы из четырех альфа-спиралей ( «застежки-молнии» ). Это переплетение дает энергию, для слияния липидных слоев, которые в норме достаточно сильно отталкиваются друг от друга из-за отрицательного заряда фосфатов мембран.

Те самые пузырьки… Синапс – основной объект лауреатов

COPII формирует пузырек В зависимости от молекул связывания, может формироваться трубочка Слияние с промежуточным компартментом АГ (с тубуло-ретикулярными кластерами) Пузырек становится «гладким»

Маннозо-6 -фосфат Лизосомальные белки Полость АГ Рецептор маннозо-6 фосфата транс-сеть цитозоль почкование адаптин Полость пузырька клатрин clathrin-coated vesicle Формирование «опушенного» пузырька, предназначенного для транспорта белка в лизосомы

Общие закономерности всех видов «пузырькового транспорта» : - Неизменная локализация молекул относительно мембраны. - Наличие сигнальных последовательностей. - Наличие неспецифического транспорта «навалом» (bulk-transport). - Перенос мембранных пузырьков регулируется ГТФазами (GTPases) семейств Rab, Arf, Arl и Sar, а также моно- и полифосфорилированными дериватами фосфатидилинозитола. -Сортировка молекул, на каждом этапе.

Химерный белок с пероксидазой хрена: формирование пузырьков на поверхности ЭПР и перемещение белка в аппарат Гольджи. «Пустые» пузырьки – возврат мембран из АГ в ЭПР «Опушенные» пузырьки в цитоплазме


Формирование транспортных везикул и трубочек на поверхности ЭПР происходит в строго определенных участках мембраны ЭПР, лишенных рибосом – в переходном, или промежуточном ЭПР (transitional ER). В клетках млекопитающих эти участки экспорта белков находятся вблизи цис-стороны аппарата Гольджи, и часто идентифицируются как «промежуточный компартмент между ЭПР и аппаратом Гольджи» , используется также название «везикуло-тубулярные кластеры» (vesicular tubular clusters).

Лек 6 2014.ppt