

Лекция 6 гистология.pptx
- Количество слайдов: 32
 Лекция 6 Нервная ткань 1. Гистогенез нервной ткани 2. Структурно-функциональная организация нервной ткани 3. Нервные волокна, их строение и классификация 4. Нервные окончания
Лекция 6 Нервная ткань 1. Гистогенез нервной ткани 2. Структурно-функциональная организация нервной ткани 3. Нервные волокна, их строение и классификация 4. Нервные окончания
 Нервная ткань – это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и передачи его. Нервная ткань состоит из нейронов (нейроцитов, собственно нервных клеток), обладающих способностью к выработке и проведению нервных импульсов, и клеток нейроглии, выполняющих ряд вспомогательных функций (опорную, трофическую, барьерную, секреторную и защитную).
Нервная ткань – это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и передачи его. Нервная ткань состоит из нейронов (нейроцитов, собственно нервных клеток), обладающих способностью к выработке и проведению нервных импульсов, и клеток нейроглии, выполняющих ряд вспомогательных функций (опорную, трофическую, барьерную, секреторную и защитную).
 развивается из дорсальной эктодермы Нервная ткань развивается из дорсальной") Гистогенез нервной ткани Нервная трубка(6) развивается из дорсальной эктодермы Нервная ткань развивается из дорсальной эктодермы. У 18 -дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, латеральные края которой приподнимаются, образуя нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки расширяется, образуя позднее головной мозг. Латеральные края продолжают подниматься и растут медиально, пока не встретятся и не сольются по средней линии в нервную трубку, которая отделяется от лежащей над ней эпидермальной эктодермы. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Часть клеток нервной пластинки не входит в состав ни нервной трубки, ни эпидермальной эктодермы, а образует скопления по бокам от нервной трубки, которые сливаются в рыхлый тяж, располагающийся между нервной трубкой и эпидермальной эктодермой, – это нервный гребень (или ганглиозная пластинка). Из нервной трубки в дальнейшем формируются нейроны и макроглия центральной нервной системы. Нервный гребень дает начало нейронам чувствительных и автономных ганглиев, клеткам мягкой мозговой и паутинной оболочек мозга и некоторым видам глии: нейролеммоцитам (шванновским клеткам), клеткам-сателлитам ганглиев. Из нервного гребня развиваются также клетки мозгового вещества надпочечников, меланоциты кожи, часть клеток APUD-системы, сенсорные клетки каротидных телец.
Гистогенез нервной ткани Нервная трубка(6) развивается из дорсальной эктодермы Нервная ткань развивается из дорсальной эктодермы. У 18 -дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, латеральные края которой приподнимаются, образуя нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки расширяется, образуя позднее головной мозг. Латеральные края продолжают подниматься и растут медиально, пока не встретятся и не сольются по средней линии в нервную трубку, которая отделяется от лежащей над ней эпидермальной эктодермы. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Часть клеток нервной пластинки не входит в состав ни нервной трубки, ни эпидермальной эктодермы, а образует скопления по бокам от нервной трубки, которые сливаются в рыхлый тяж, располагающийся между нервной трубкой и эпидермальной эктодермой, – это нервный гребень (или ганглиозная пластинка). Из нервной трубки в дальнейшем формируются нейроны и макроглия центральной нервной системы. Нервный гребень дает начало нейронам чувствительных и автономных ганглиев, клеткам мягкой мозговой и паутинной оболочек мозга и некоторым видам глии: нейролеммоцитам (шванновским клеткам), клеткам-сателлитам ганглиев. Из нервного гребня развиваются также клетки мозгового вещества надпочечников, меланоциты кожи, часть клеток APUD-системы, сенсорные клетки каротидных телец.
 Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных, или нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны: • внутренняя - вентрикулярная (или эпендимная) зона, • вокруг нее – субвентрикулярная зона, • затем промежуточная (или плащевая, или же мантийная, зона) и, наконец, • наружная - краевая (или маргинальная) зона нервной трубки.
Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных, или нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны: • внутренняя - вентрикулярная (или эпендимная) зона, • вокруг нее – субвентрикулярная зона, • затем промежуточная (или плащевая, или же мантийная, зона) и, наконец, • наружная - краевая (или маргинальная) зона нервной трубки.
, внутренняя, зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные (или матричные) клетки") Вентрикулярная (эпендимная), внутренняя, зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные (или матричные) клетки являются предшественниками нейронов и клеток макроглии. Субвентрикулярная зона состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая, или мантийная) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон – нейробластов и глиобластов. Нейробласты утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам. Способность к делению не утрачивают полностью и зрелые глиоциты. Новообразование нейронов прекращается в раннем постнатальном периоде. Поскольку число нейронов в головном мозге составляет примерно 1 триллион, очевидно, в среднем в течение всего пренатального периода в 1 мин формируется 2, 5 миллиона нейронов. Из клеток плащевого слоя образуются серое вещество спинного и часть серого вещества головного мозга. Маргинальная зона (или краевая вуаль) формируется из врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу. В некоторых областях головного мозга клетки плащевого слоя мигрируют дальше, образуя кортикальные пластинки — скопления клеток, из которых формируется кора большого мозга и мозжечка (т. е. серое вещество).
Вентрикулярная (эпендимная), внутренняя, зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные (или матричные) клетки являются предшественниками нейронов и клеток макроглии. Субвентрикулярная зона состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая, или мантийная) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон – нейробластов и глиобластов. Нейробласты утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам. Способность к делению не утрачивают полностью и зрелые глиоциты. Новообразование нейронов прекращается в раннем постнатальном периоде. Поскольку число нейронов в головном мозге составляет примерно 1 триллион, очевидно, в среднем в течение всего пренатального периода в 1 мин формируется 2, 5 миллиона нейронов. Из клеток плащевого слоя образуются серое вещество спинного и часть серого вещества головного мозга. Маргинальная зона (или краевая вуаль) формируется из врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу. В некоторых областях головного мозга клетки плащевого слоя мигрируют дальше, образуя кортикальные пластинки — скопления клеток, из которых формируется кора большого мозга и мозжечка (т. е. серое вещество).
 Структурно-функциональная организация нервной ткани Нервная ткань состоит из клеток – нейронов и межклеточного вещества – нейроглии. Нейроны (нейроциты) – клетки различных размеров (от 4 -5 мкм до 140 мкм). Их общее количество в нервной системе человека превышает 100 млрд. К рождению нейроны утрачивают способность к делению. Нейрон является морфологически и функционально самостоятельной единицей, но с помощью своих отростков осуществляет синаптический контакт с другими нейронами, образуя рефлекторные дуги – звенья цепи, из которой построена нервная система. В зависимости от функции в рефлекторной дуге различают три типа нейронов: афферентные, ассоциативные и эфферентные.
Структурно-функциональная организация нервной ткани Нервная ткань состоит из клеток – нейронов и межклеточного вещества – нейроглии. Нейроны (нейроциты) – клетки различных размеров (от 4 -5 мкм до 140 мкм). Их общее количество в нервной системе человека превышает 100 млрд. К рождению нейроны утрачивают способность к делению. Нейрон является морфологически и функционально самостоятельной единицей, но с помощью своих отростков осуществляет синаптический контакт с другими нейронами, образуя рефлекторные дуги – звенья цепи, из которой построена нервная система. В зависимости от функции в рефлекторной дуге различают три типа нейронов: афферентные, ассоциативные и эфферентные.
 нейроны генерируют нервные импульсы под влиянием изменений внешней или") Чувствительные или сенсорные (афферентные) нейроны генерируют нервные импульсы под влиянием изменений внешней или внутренней среды. Двигательные или эфферентные нейроны передают сигналы на рабочие органы. Ассоциативные (вставочные) нейроны осуществляют связь между нейронами, составляя 99, 98% от общего числа этих клеток. 1 – афферентный нейрон 2 – вставочный нейрон 3 – эфферентный нейрон 4 – нервные волокна 5 – волокна корковоспиномозгового пути
Чувствительные или сенсорные (афферентные) нейроны генерируют нервные импульсы под влиянием изменений внешней или внутренней среды. Двигательные или эфферентные нейроны передают сигналы на рабочие органы. Ассоциативные (вставочные) нейроны осуществляют связь между нейронами, составляя 99, 98% от общего числа этих клеток. 1 – афферентный нейрон 2 – вставочный нейрон 3 – эфферентный нейрон 4 – нервные волокна 5 – волокна корковоспиномозгового пути
 Нейроны отличаются большим разнообразием форм и размеров. Например, диаметр тел клеток-зерен коры мозжечка 4 -6 мкм, а гигантских пирамидных нейронов двигательной зоны коры большого мозга – 130 -150 мкм. Нейроны состоят из тела (или перикариона) и отростков: одного аксона и различного числа ветвящихся дендритов. По количеству отростков различают три типа нейронов: • униполярные, • биполярные • мультиполярные нейроны. Морфологические типы нейронов 1 - униполярный, 2 - биполярный, 3 - псевдоуниполярный, 4 - мультиполярный
Нейроны отличаются большим разнообразием форм и размеров. Например, диаметр тел клеток-зерен коры мозжечка 4 -6 мкм, а гигантских пирамидных нейронов двигательной зоны коры большого мозга – 130 -150 мкм. Нейроны состоят из тела (или перикариона) и отростков: одного аксона и различного числа ветвящихся дендритов. По количеству отростков различают три типа нейронов: • униполярные, • биполярные • мультиполярные нейроны. Морфологические типы нейронов 1 - униполярный, 2 - биполярный, 3 - псевдоуниполярный, 4 - мультиполярный
 Биохимическая классификация нейронов В зависимости от природы вырабатываемого нейромедиатора нейроны подразделяют на: • холинергические (медиатор ацетилхолин); • серотонинергические (серотонин); • дофаминергические (дофамин); • адрегергические (норадреналин, адреналин); • ГАМК-ергические (ГАМК); • пуринергические (АТФ); • пептидергические (энкефалины, эндорфины, холицистокенин, бомбезин).
Биохимическая классификация нейронов В зависимости от природы вырабатываемого нейромедиатора нейроны подразделяют на: • холинергические (медиатор ацетилхолин); • серотонинергические (серотонин); • дофаминергические (дофамин); • адрегергические (норадреналин, адреналин); • ГАМК-ергические (ГАМК); • пуринергические (АТФ); • пептидергические (энкефалины, эндорфины, холицистокенин, бомбезин).
 и отростков, обеспечивающих проведение нервных импульсов") Строение нейрона Нейрон состоит из клеточного тела (сомы) и отростков, обеспечивающих проведение нервных импульсов – дендритов, переносящих импульсы к телу нейрона, и аксона (нейрит), несущего импульс от тела нейрона. Подавляющее большинство нейронов человека содержит одно округлое светлое ядро, расположенное в центре клетки. Двуядерные и тем более многоядерные нейроны встречаются крайне редко. Плазмолемма нейрона является возбудимой мембраной, т. е. обладает способностью генерировать и проводить импульс. Ее интегральными белками являются белки, функционирующие как ионно-избирательные каналы, и рецепторные белки, вызывающие реакции нейронов на специфические стимулы. В нейроне мембранный потенциал покоя равен – 60 -70 м. В. Потенциал покоя создается за счет выведения Na+ из клетки. Большинство Na+- и К+-каналов при этом закрыты. Переход каналов из закрытого состояния в открытое регулируется мембранным потенциалом.
Строение нейрона Нейрон состоит из клеточного тела (сомы) и отростков, обеспечивающих проведение нервных импульсов – дендритов, переносящих импульсы к телу нейрона, и аксона (нейрит), несущего импульс от тела нейрона. Подавляющее большинство нейронов человека содержит одно округлое светлое ядро, расположенное в центре клетки. Двуядерные и тем более многоядерные нейроны встречаются крайне редко. Плазмолемма нейрона является возбудимой мембраной, т. е. обладает способностью генерировать и проводить импульс. Ее интегральными белками являются белки, функционирующие как ионно-избирательные каналы, и рецепторные белки, вызывающие реакции нейронов на специфические стимулы. В нейроне мембранный потенциал покоя равен – 60 -70 м. В. Потенциал покоя создается за счет выведения Na+ из клетки. Большинство Na+- и К+-каналов при этом закрыты. Переход каналов из закрытого состояния в открытое регулируется мембранным потенциалом.
 Тигроид в нервных клетках При окрашивании нервной ткани анилиновыми красителями в цитоплазме нейронов выявляется хроматофильная субстанция в виде базофильных глыбок и зерен различных размеров и форм (другие названия хроматофильной субстанции - тигроид, тельца Ниссля). Базофильные глыбки локализуются в перикарионах и дендритах нейронов, но никогда не обнаруживаются в аксонах и их конусовидных основаниях – аксональных холмиках. Базофилия глыбок объясняется высоким содержанием рибонуклеопротеидов. Каждая глыбка хроматофильной субстанции состоит из цистерн гранулярной эндоплазматической сети, свободных рибосом и полисом.
Тигроид в нервных клетках При окрашивании нервной ткани анилиновыми красителями в цитоплазме нейронов выявляется хроматофильная субстанция в виде базофильных глыбок и зерен различных размеров и форм (другие названия хроматофильной субстанции - тигроид, тельца Ниссля). Базофильные глыбки локализуются в перикарионах и дендритах нейронов, но никогда не обнаруживаются в аксонах и их конусовидных основаниях – аксональных холмиках. Базофилия глыбок объясняется высоким содержанием рибонуклеопротеидов. Каждая глыбка хроматофильной субстанции состоит из цистерн гранулярной эндоплазматической сети, свободных рибосом и полисом.
 Дендриты представляют собой истинные выпячивания тела клетки. Они содержат те же органеллы, что и тело клетки: глыбки хроматофильной субстанции (т. е. гранулярной эндоплазматической сети и полисом), митохондрии, большое количество нейротубул (или микротрубочек) и нейрофиламентов. За счет дендритов рецепторная поверхность нейрона увеличивается в 1000 и более раз. Аксон – это отросток, по которому импульс передается от тела клетки. Он содержит митохондрии, нейротубулы и нейрофиламенты, а также гладкую эндоплазматическую сеть. Возрастные изменения нейронов сопровождаются накоплением липофусцина, разрушением крист митохондрий. Липофусцин — «пигмент старения» – желто-бурого цвета липопротеидной природы, представляющий собой остаточные тельца (т. е. телолизосомы) с продуктами непереваренных структур.
Дендриты представляют собой истинные выпячивания тела клетки. Они содержат те же органеллы, что и тело клетки: глыбки хроматофильной субстанции (т. е. гранулярной эндоплазматической сети и полисом), митохондрии, большое количество нейротубул (или микротрубочек) и нейрофиламентов. За счет дендритов рецепторная поверхность нейрона увеличивается в 1000 и более раз. Аксон – это отросток, по которому импульс передается от тела клетки. Он содержит митохондрии, нейротубулы и нейрофиламенты, а также гладкую эндоплазматическую сеть. Возрастные изменения нейронов сопровождаются накоплением липофусцина, разрушением крист митохондрий. Липофусцин — «пигмент старения» – желто-бурого цвета липопротеидной природы, представляющий собой остаточные тельца (т. е. телолизосомы) с продуктами непереваренных структур.
 Из элементов цитоскелета в цитоплазме нейронов присутствуют нейрофиламенты и нейротубулы. Пучки нейрофиламентов на препаратах, импрегнированных серебром, видны в виде нитей – нейрофибрилл. Нейрофибриллы образуют сеть в теле нейрона, а в отростках расположены параллельно. Нейротубулы и нейрофиламенты участвуют в поддержании формы клеток, росте отростков и аксональном транспорте. Аксональный (точнее аксоплазматический) транспорт – это перемещение веществ от тела в отростки и от отростков в тело нейрона. Он направляется нейротубулами, а в транспорте участвуют белки – кинезин и динеин. Транспорт веществ от тела клетки в отростки называется прямым, или антероградным, транспорт веществ от отростков к телу – обратным, или ретроградным. Аксональный транспорт представлен двумя главными компонентами: быстрым компонентом (400 -2000 мм в сутки) и медленным (1 -2 мм в сутки). Обе транспортные системы присутствуют как в аксонах, так и в дендритах. Антероградная быстрая система проводит мембранные структуры, включая компоненты мембраны, митохондрии, пузырьки, содержащие пептиды, предшественники нейромедиаторов и другие белки. Ретроградная быстрая система проводит использованные материалы для деградации в лизосомах, распределения и рециркуляции и, возможно, факторы роста нервов.
Из элементов цитоскелета в цитоплазме нейронов присутствуют нейрофиламенты и нейротубулы. Пучки нейрофиламентов на препаратах, импрегнированных серебром, видны в виде нитей – нейрофибрилл. Нейрофибриллы образуют сеть в теле нейрона, а в отростках расположены параллельно. Нейротубулы и нейрофиламенты участвуют в поддержании формы клеток, росте отростков и аксональном транспорте. Аксональный (точнее аксоплазматический) транспорт – это перемещение веществ от тела в отростки и от отростков в тело нейрона. Он направляется нейротубулами, а в транспорте участвуют белки – кинезин и динеин. Транспорт веществ от тела клетки в отростки называется прямым, или антероградным, транспорт веществ от отростков к телу – обратным, или ретроградным. Аксональный транспорт представлен двумя главными компонентами: быстрым компонентом (400 -2000 мм в сутки) и медленным (1 -2 мм в сутки). Обе транспортные системы присутствуют как в аксонах, так и в дендритах. Антероградная быстрая система проводит мембранные структуры, включая компоненты мембраны, митохондрии, пузырьки, содержащие пептиды, предшественники нейромедиаторов и другие белки. Ретроградная быстрая система проводит использованные материалы для деградации в лизосомах, распределения и рециркуляции и, возможно, факторы роста нервов.
 Нейротубулы – органеллы, ответственные за быстрый транспорт, который называется также нейротубулозависимым. Каждая нейротубула содержит несколько путей, вдоль которых движутся различные частички. АТФ и ионы Са 2+ обеспечивают эти движения. На одной микротубуле пузырьки могут обгонять другие пузырьки, движущиеся в том же направлении. Два пузырька могут двигаться в противоположных направлениях одновременно по различным путям одной нейротубулы. Медленный транспорт – это антероградная система, проводящая белки и другие вещества для обновления и поддержания аксоплазмы зрелых нейронов и обеспечения аксоплазмой роста аксонов и дендритов при развитии и регенерации. Отдельной разновидностью нейронов являются секреторные нейроны. Способность синтезировать и секретировать биологически активные вещества, в частности нейромедиаторы, свойственна всем нейроцитам. Однако существуют нейроциты, специализированные преимущественно для выполнения этой функции, — секреторные нейроны, например клетки нейросекреторных ядер гипоталамической области головного мозга. В цитоплазме таких нейронов и в их аксонах находятся различной величины гранулы нейросекрета, содержащие белок, а в некоторых случаях липиды и полисахариды. Гранулы нейросекрета выводятся непосредственно в кровь (например, с помощью т. н. аксо-вазальных синапсов) или же в мозговую жидкость. Нейросекреты выполняют роль нейрорегуляторов, участвуя во взаимодействии нервной и гуморальной систем интеграции.
Нейротубулы – органеллы, ответственные за быстрый транспорт, который называется также нейротубулозависимым. Каждая нейротубула содержит несколько путей, вдоль которых движутся различные частички. АТФ и ионы Са 2+ обеспечивают эти движения. На одной микротубуле пузырьки могут обгонять другие пузырьки, движущиеся в том же направлении. Два пузырька могут двигаться в противоположных направлениях одновременно по различным путям одной нейротубулы. Медленный транспорт – это антероградная система, проводящая белки и другие вещества для обновления и поддержания аксоплазмы зрелых нейронов и обеспечения аксоплазмой роста аксонов и дендритов при развитии и регенерации. Отдельной разновидностью нейронов являются секреторные нейроны. Способность синтезировать и секретировать биологически активные вещества, в частности нейромедиаторы, свойственна всем нейроцитам. Однако существуют нейроциты, специализированные преимущественно для выполнения этой функции, — секреторные нейроны, например клетки нейросекреторных ядер гипоталамической области головного мозга. В цитоплазме таких нейронов и в их аксонах находятся различной величины гранулы нейросекрета, содержащие белок, а в некоторых случаях липиды и полисахариды. Гранулы нейросекрета выводятся непосредственно в кровь (например, с помощью т. н. аксо-вазальных синапсов) или же в мозговую жидкость. Нейросекреты выполняют роль нейрорегуляторов, участвуя во взаимодействии нервной и гуморальной систем интеграции.
 Нейроглия, ее структурно-функциональная организация Нейроглия выполняет следующие функции: опорную, трофическую, разграничительную, поддержание постоянства среды вокруг нейронов, защитную, секреторную. Различают глию центральной и периферической нервной системы.
Нейроглия, ее структурно-функциональная организация Нейроглия выполняет следующие функции: опорную, трофическую, разграничительную, поддержание постоянства среды вокруг нейронов, защитную, секреторную. Различают глию центральной и периферической нервной системы.
 Астроциты – клетки отростчатой формы, бедные органеллами. Они выполняют в основном опорную и трофическую функции. Различают два типа астроцитов - протоплазматические и волокнистые. Протоплазматические астроциты локализуются в сером веществе центральной нервной системы, а волокнистые астроциты преимущественно в белом веществе. Протоплазматические астроциты характеризуются короткими сильно ветвящимися отростками и светлым сферическим ядром.
Астроциты – клетки отростчатой формы, бедные органеллами. Они выполняют в основном опорную и трофическую функции. Различают два типа астроцитов - протоплазматические и волокнистые. Протоплазматические астроциты локализуются в сером веществе центральной нервной системы, а волокнистые астроциты преимущественно в белом веществе. Протоплазматические астроциты характеризуются короткими сильно ветвящимися отростками и светлым сферическим ядром.
 Олигодендроциты – имеют более мелкие по сравнению с астроцитами и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в миелиновых нервных волокнах, причем, в противоположность аналогичным клеткам периферической нервной системы – нейролеммоцитам, один олигодендроглиоцит может участвовать в миелинизации сразу нескольких аксонов.
Олигодендроциты – имеют более мелкие по сравнению с астроцитами и более интенсивно окрашивающиеся ядра. Их отростки немногочисленны. Олигодендроглиоциты присутствуют как в сером, так и в белом веществе. В сером веществе они локализуются вблизи перикарионов. В белом веществе их отростки образуют миелиновый слой в миелиновых нервных волокнах, причем, в противоположность аналогичным клеткам периферической нервной системы – нейролеммоцитам, один олигодендроглиоцит может участвовать в миелинизации сразу нескольких аксонов.
 Эпендимоциты выстилают желудочки головного мозга и центральный канал спинного мозга. Эти клетки цилиндрической формы. Они образуют слой типа эпителия, носящий название эпендимы. Между соседними клетками эпендимы имеются щелевидные соединения и пояски сцепления, но плотные соединения отсутствуют, так что цереброспинальная жидкость может проникать между эпендимоцитами в нервную ткань. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости.
Эпендимоциты выстилают желудочки головного мозга и центральный канал спинного мозга. Эти клетки цилиндрической формы. Они образуют слой типа эпителия, носящий название эпендимы. Между соседними клетками эпендимы имеются щелевидные соединения и пояски сцепления, но плотные соединения отсутствуют, так что цереброспинальная жидкость может проникать между эпендимоцитами в нервную ткань. Большинство эпендимоцитов имеют подвижные реснички, вызывающие ток цереброспинальной жидкости.
 Микроциты Микроглия представляет собой фагоцитирующие клетки, относящиеся к системе мононуклеарных фагоцитов и происходящие из стволовой кроветворной клетки (возможно, из премоноцитов красного костного мозга). Функция микроглии — защита от инфекции и повреждения, и удаление продуктов разрушения нервной ткани. Клетки микроглии характеризуются небольшими размерами, телами продолговатой формы. Их короткие отростки имеют на своей поверхности вторичные и третичные ответвления, что придает клеткам «колючий» вид.
Микроциты Микроглия представляет собой фагоцитирующие клетки, относящиеся к системе мононуклеарных фагоцитов и происходящие из стволовой кроветворной клетки (возможно, из премоноцитов красного костного мозга). Функция микроглии — защита от инфекции и повреждения, и удаление продуктов разрушения нервной ткани. Клетки микроглии характеризуются небольшими размерами, телами продолговатой формы. Их короткие отростки имеют на своей поверхности вторичные и третичные ответвления, что придает клеткам «колючий» вид.
 Периферическая глия Глия периферической нервной системы в отличие от макроглии центральной нервной системы происходит из нервного гребня. К периферической нейроглии относятся: нейролеммоциты (или шванновские клетки) и глиоциты ганглиев (или мантийные глиоциты). Нейролеммоциты Шванна формируют оболочки отростков нервных клеток в нервных волокнах периферической нервной системы. Мантийные глиоциты ганглиев окружают тела нейронов в нервных узлах и участвуют в обмене веществ этих нейронов. Нейролемоциты Глиоциты ганлглиев
Периферическая глия Глия периферической нервной системы в отличие от макроглии центральной нервной системы происходит из нервного гребня. К периферической нейроглии относятся: нейролеммоциты (или шванновские клетки) и глиоциты ганглиев (или мантийные глиоциты). Нейролеммоциты Шванна формируют оболочки отростков нервных клеток в нервных волокнах периферической нервной системы. Мантийные глиоциты ганглиев окружают тела нейронов в нервных узлах и участвуют в обмене веществ этих нейронов. Нейролемоциты Глиоциты ганлглиев
 Нервные волокна Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. По строению оболочек различают миелиновые и безмиелиновые нервные волокна. Безмиелиновое нервное волокно В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической – нейролеммоцитами Шванна. Миелиновое нервное волокно
Нервные волокна Отростки нервных клеток, покрытые оболочками, называются нервными волокнами. По строению оболочек различают миелиновые и безмиелиновые нервные волокна. Безмиелиновое нервное волокно В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической – нейролеммоцитами Шванна. Миелиновое нервное волокно
 Безмиелиновое нервное волокно Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану – мезаксон, на которой как бы подвешен осевой цилиндр.
Безмиелиновое нервное волокно Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану – мезаксон, на которой как бы подвешен осевой цилиндр.
 Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии – насечки миелина, или насечки Шмидта – Лантермана. Через определенные интервалы (1 -2 мм) видны участки волокна, лишенные миелинового слоя, – это т. н. узловатые перехваты, или перехваты Ранвье. В процессе миелинизации аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита – мезаксон. Мезаксон удлиняется, концентрически наслаивается (как бы накручивается) на осевой цилиндр и образует вокруг него плотную слоистую зону – миелиновый слой. Отсутствие миелинового слоя в области узловых перехватов объясняется тем, что в этом участке волокна кончается один нейролеммоцит и начинается другой. Осевой цилиндр в этом месте частично прикрыт интердигитирующими отростками нейролеммоцитов. Оболочка аксона (аксолемма) обладает в области перехвата значительной электронной плотностью.
Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии – насечки миелина, или насечки Шмидта – Лантермана. Через определенные интервалы (1 -2 мм) видны участки волокна, лишенные миелинового слоя, – это т. н. узловатые перехваты, или перехваты Ранвье. В процессе миелинизации аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита – мезаксон. Мезаксон удлиняется, концентрически наслаивается (как бы накручивается) на осевой цилиндр и образует вокруг него плотную слоистую зону – миелиновый слой. Отсутствие миелинового слоя в области узловых перехватов объясняется тем, что в этом участке волокна кончается один нейролеммоцит и начинается другой. Осевой цилиндр в этом месте частично прикрыт интердигитирующими отростками нейролеммоцитов. Оболочка аксона (аксолемма) обладает в области перехвата значительной электронной плотностью.
 Миелиновые и безмиелиновые нервные волокна
Миелиновые и безмиелиновые нервные волокна
 Нервные волокна заканчиваются концевыми аппаратами – нервными окончаниями. Различают три группы нервных окончаний: • межнейрональные синапсы, осуществляющие связь нейронов между собой; • эффекторные окончания (эффекторы), передающие нервный импульс на ткани рабочего органа (на мышечные или железистые клетки) • рецепторные (или Различные типы нервных окончаний в коже аффекторные, или чувствительные) окончания Нервные окончания
Нервные волокна заканчиваются концевыми аппаратами – нервными окончаниями. Различают три группы нервных окончаний: • межнейрональные синапсы, осуществляющие связь нейронов между собой; • эффекторные окончания (эффекторы), передающие нервный импульс на ткани рабочего органа (на мышечные или железистые клетки) • рецепторные (или Различные типы нервных окончаний в коже аффекторные, или чувствительные) окончания Нервные окончания
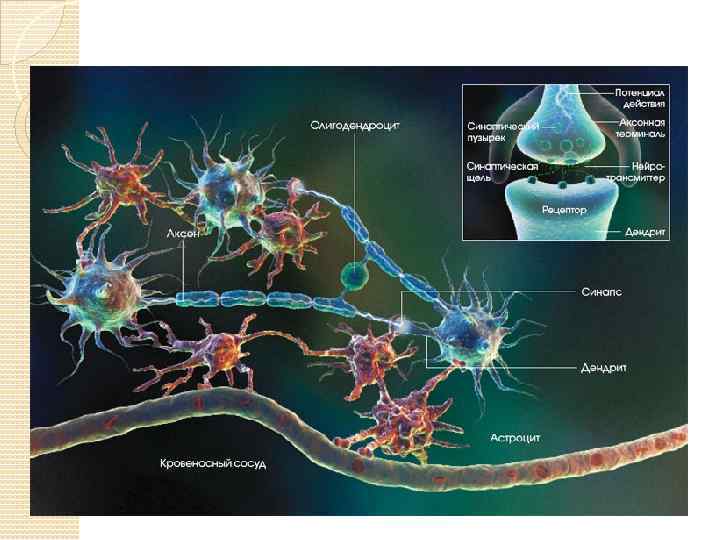
 Синапсы – это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры. В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы различают: аксо-дендритические, аксо-соматические, аксо-аксональные. В целом процессы в синапсе происходят в следующем порядке: 1. Волна деполяризации доходит до пресинаптической мембраны. 2. При этом открываются кальциевые каналы, и ионы Са 2+ входят в терминаль. 3. Повышение концентрации ионов Са 2+ в терминали вызывает экзоцитоз нейромедиатора, и медиатор попадает в синаптическую щель. 4. Далее, нейромедиатор диффундирует через синаптическую щель и связывается со специфическими рецепторными участками на постсинаптической мембране, что вызывает молекулярные изменения в постсинаптической мембране, приводящие к открытию ионных каналов и созданию постсинаптических потенциалов, обусловливающих реакции возбуждения или торможения.
Синапсы – это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры. В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы различают: аксо-дендритические, аксо-соматические, аксо-аксональные. В целом процессы в синапсе происходят в следующем порядке: 1. Волна деполяризации доходит до пресинаптической мембраны. 2. При этом открываются кальциевые каналы, и ионы Са 2+ входят в терминаль. 3. Повышение концентрации ионов Са 2+ в терминали вызывает экзоцитоз нейромедиатора, и медиатор попадает в синаптическую щель. 4. Далее, нейромедиатор диффундирует через синаптическую щель и связывается со специфическими рецепторными участками на постсинаптической мембране, что вызывает молекулярные изменения в постсинаптической мембране, приводящие к открытию ионных каналов и созданию постсинаптических потенциалов, обусловливающих реакции возбуждения или торможения.
 Эффекторные нервные окончания Двигательные нервные окончания – это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы. При их участии нервный импульс передается на ткани рабочих органов. Двигательные окончания в поперечнополосатых мышцах называются нервно-мышечными окончаниями. Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра нервного волокна и специализированного участка мышечного волокна. Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой и погружается в него, вовлекая за собой его плазмолемму и базальную мембрану. Мембрана мышечного волокна образует многочисленные складки, формирующие вторичные синаптические щели эффекторного окончания. В области окончания мышечное волокно не имеет типичной поперечной исчерченности и характеризуется обилием митохондрий, скоплением круглых или слегка овальных ядер.
Эффекторные нервные окончания Двигательные нервные окончания – это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы. При их участии нервный импульс передается на ткани рабочих органов. Двигательные окончания в поперечнополосатых мышцах называются нервно-мышечными окончаниями. Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра нервного волокна и специализированного участка мышечного волокна. Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой и погружается в него, вовлекая за собой его плазмолемму и базальную мембрану. Мембрана мышечного волокна образует многочисленные складки, формирующие вторичные синаптические щели эффекторного окончания. В области окончания мышечное волокно не имеет типичной поперечной исчерченности и характеризуется обилием митохондрий, скоплением круглых или слегка овальных ядер.
 Рецепторные нервные окончания Рецепторы – рассеяны по всему организму и воспринимают различные раздражения, как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов: экстерорецепторы и интерорецепторы. К экстерорецепторам (внешним) относятся: слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам (внутренним) относятся: висцерорецепторы (сигнализирующие о состоянии внутренних органов) и проприорецепторы (или рецепторы опорно-двигательного аппарата).
Рецепторные нервные окончания Рецепторы – рассеяны по всему организму и воспринимают различные раздражения, как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов: экстерорецепторы и интерорецепторы. К экстерорецепторам (внешним) относятся: слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам (внутренним) относятся: висцерорецепторы (сигнализирующие о состоянии внутренних органов) и проприорецепторы (или рецепторы опорно-двигательного аппарата).
 Свободные нервные окончания состоят только из конечных ветвлений осевого цилиндра Свободные нервные окончания обычно воспринимают холод, тепло и боль. Такие окончания характерны для эпителия.
Свободные нервные окончания состоят только из конечных ветвлений осевого цилиндра Свободные нервные окончания обычно воспринимают холод, тепло и боль. Такие окончания характерны для эпителия.
 Несвободные нервные окончания Несвободные, содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии. Несвободные окончания, кроме того, могут быть покрыты соединительнотканной капсулой, и тогда они называются инкапсулированными. Несвободные нервные окончания, не имеющие соединительнотканной капсулы, называются неинкапсулированными.
Несвободные нервные окончания Несвободные, содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии. Несвободные окончания, кроме того, могут быть покрыты соединительнотканной капсулой, и тогда они называются инкапсулированными. Несвободные нервные окончания, не имеющие соединительнотканной капсулы, называются неинкапсулированными.
 Рецепторные нервные окончания К инкапсулированным нервным окончаниям относятся также рецепторы мышц и сухожилий: это нервно-мышечные веретена и нервно-сухожильные веретена. Нервно-мышечные веретена являются сенсорными органами в скелетных мышцах, которые функционируют как рецептор на растяжение. Веретено состоит из нескольких исчерченных мышечных волокон — интрафузальных волокон, заключенных в растяжимую соединительнотканную капсулу. Между капсулой и интрафузальными волокнами имеется заполненное жидкостью пространство.
Рецепторные нервные окончания К инкапсулированным нервным окончаниям относятся также рецепторы мышц и сухожилий: это нервно-мышечные веретена и нервно-сухожильные веретена. Нервно-мышечные веретена являются сенсорными органами в скелетных мышцах, которые функционируют как рецептор на растяжение. Веретено состоит из нескольких исчерченных мышечных волокон — интрафузальных волокон, заключенных в растяжимую соединительнотканную капсулу. Между капсулой и интрафузальными волокнами имеется заполненное жидкостью пространство.