Лекция 6 (Летающие архозавры ).ppt
- Количество слайдов: 11



Лекция 6 Летающие архозавры
 рептилий. Долгое время считалось, что")
Птицы представляют собой наиболее высокоорганизованную эволюционную ветвь завроморфных (типичных) рептилий. Долгое время считалось, что наличие перьев является уникальным признаком птиц, но теперь известны оперенные динозавры без признаков способности к полету. Sinosauropteryx из раннего мела Китая, достигавший в длину 1 м , представлял собой изящного теропода, по общему плану теропода, строения весьма похожего на компсогната, и имел пухоподобный покров. Поскольку ничего похожего на маховые и рулевые перья у этого компсогната, динозавра не было, следует признать, что первоначальное возникновение перьев связано с нуждами терморегуляции. Вполне возможно, что перьевой покров имели многие мелкие динозавры, но условия сохранности ископаемого материала этого не демонстрируют. Хорошо сохранившиеся отпечатки кожи крупных динозавров (гадрозавров и цератопсов), свидетельствуют об отсутствии у них каких-либо теплооизолирующих покровов, что естественно для животных с таким соотношением поверхности тела и его объема, живущих в теплом климате, вне зависимости от того, эндотермные они или эктотермные. Первое животное, ставшее известным из числа «первоптиц» - Archaeopteryx из верхней юры Европы демонстрирует несколько иной план «первоптиц» строения. Скелет его достаточно типичен для мелких теропод, за исключением крупных передних конечностей, не уступающих по длине теропод, задним. Цевка из частично слитых плюсневых костей, как у цератозаврид. Череп очень похож на череп компсогната облегченной лицевой цератозаврид. частью, крупными глазницами и мозговой коробкой, что естественно при миниатюризации исходной модели (археоптерикс размером с ворону). По бокам хвоста располагались, как свидетельствуют отпечатки на тонкозернистом известняке, рулевые перья, а кисть и предплечья несли маховые перья с асимметричным опахалом, по абсолютной и относительной длине сопоставимые с аналогичными перьями сороки. Следов перьев, располагавшихся на других частях тела, не обнаружено. Это может говорить или о том, что перья могли возникнуть изначально и для выполнения аэродинамических функций (на то, что они ее выполняли, указывают их асимметричные опахала), или о том, что они были, но утратили связь с телом до фоссилизации образцов. Впрочем, эксперименты с трупами современных птиц показали, что у них туловищные перья при захоронении связи с туловищем не теряют. Грудина у археоптерикса, как у всех типичных динозавров, не окостеневала, что делало невозможным активный полет. Коракоид обычной для динозавров конфигурации, что делало невозможным антагонизм надкоракодной мышцы относительно грудной, характерный для современных птиц. Долгое время считалось, что предки птиц вели древесный образ жизни, а к полету перешли через планирование, увеличивающее длину прыжка с ветки на ветку. Это объясняло отсутствие у археоптерикса мощных грудных мышц, наряду с асимметрией опахал маховых перьев, а несомненную способность передних конечностей археоптерикса к хватанию, типичную для мелких теропод, связывали с передвижением среди древесной растительности. Из верхнего триаса Северной Америки известны гораздо хуже сохранившиеся остатки похожего на археоптерикса животного, отнесенного к роду Protoavis. При сходных размерах он имел еще больше общего с птицами: верхняя скуловая дуга утрачена, квадратная кость подвижна, Protoavis. череп кинетичен, шейные позвонки гетероцельные. Отпечатки перьев не сохранились, но на заднем крае локтевой кости имеются ямочки кинетичен, гетероцельные. для их прикрепления. Поскольку во времена господства «древесной» гипотезы (начало 20 в. ) были известны лишь сильно специализированные в иных направлениях динозавры, происхождение птиц связывали непосредственно с текодонтами. Но теперь известно, что все общие для этих двух групп признаки являются примитивными для архозавров в целом, а специализированные признаки птиц встречаются лишь у динозавров. Строение задних конечностей археоптерикса ясно указывает на адаптацию к быстрой наземной локомоции. Признаки древесной специализации в них отсутствуют, хотя их строение не препятствует передвижению по веткам, так же как не препятствует этому строение ног курицы. У планирующих животных, таких как летяги, удлинены проксимальные отделы конечностей, а у археоптерикса и современных птиц – дистальные. «Приземление» на тонкие ветви требует крайне высоко развитой способности к полету и маневренности, свойственных только наиболее продвинутым группам современных птиц, а значит, первые птицы вели преимущественно наземный образ жизни.


Подробное изучение во второй половине 20 в. дромеозаврид и их большое анатомическое сходство с птицами позволило предположить, что полет возник из способа ловли добычи, включающего «апплодирующие» движения передних конечностей. Согласно этой теории, мелкие «апплодирующие» динозавры, похожие на дромеозаврид, ловили передними лапами насекомых, а зачаточные крылья увеличивали их ловчую поверхность, что дромеозаврид, важно при захвате мелкой подвижной добычи. Движения, использовавшиеся мелкими насекомоядными динозаврами при захвате добычи, сопоставимы со взмахами крыльев в полете у птиц, и по мере удлинения маховых перьев первое могло постепенно переходить во второе на фоне прыжков за насекомыми. Последнее предположение очень важно, так как позволяет обосновать преимущества каждой последующей эволюционной стадии по сравнению с предыдущей, иллюстрируя плавный и естественный переход. (Очень часто ученые придумывают гипотетические промежуточные стадии, связывающие одну реальную форму с другой, не считаясь с тем, что это должны были быть не только жизнеспособные организмы, но и наделенные явными преимуществами по сравнению со своими предками). Это нисколько не противоречит и аэродинамической функции передних конечностей и хвоста, так как при прыжках за добычей даже небольшая подъемная сила могла существенно увеличивать маневренность и устойчивость. (Достаточно вспомнить, как используют крылья маневрирующие на бегу куры и даже страусы). Protoarchaeopteryx и Caudipteryx из раннего мела Китая блестяще подтвердили эту гипотезу. Эти тероподы имели в длину около 1 м, и обладали рулевыми перьями на заднем конце относительного недлинного хвоста, а их маховые перья были явно малы для полета, но достаточны для повышения эффективности ловли насекомых и маневрирования на бегу. Скорее всего, «древесная» и «наземная» гипотезы происхождения птиц вообще не должны противоречить другу. Происхождение птиц в своих истоках, по-видимому, связано с миниатюризацией исходной модели теропод, что давало им возможность поспорить с теропод, млекопитающими за экологическую нишу мелких насекомоядных животных. Но исходный план строения бипедальных динозавров – это план строения достаточно крупных (по крайней мере, с индюка) животных, приспособленных к передвижению по ровному субстрату. (Насколько двуногий план строения неудобен для резких подъемов и спусков, знает каждый человек, сравнивавший свои физические возможности в этой области, скажем, с собачьими или козьими). Таким образом, мелким динозаврам сильно мешали передвигаться любые поваленные стволы, кочки, ямы и тому подобное, что должно было стимулировать у них способность прыгать и цепляться за негоризонтальный субстрат. При мелких размерах тела различия между наземным и древесным образом жизни просто стираются. Здесь же следует вспомнить, что и дромеозавриды, сходные с птицами по своей анатомии и наверняка имевшие с ним и недавних общих предков также, судя по всему, дромеозавриды, прекрасно прыгали и могли цепляться на негоризонтальном субстрате (своих жертвах) передними конечностями. Можно предположить, что первые птицы были наземными насекомоядными животными, способными, тем не менее, запрыгивать и взлетать на деревья, то есть вели образ жизни, похожий на образ жизни современных дроздов. Важной находкой начала 21 в. стал Microraptor из нижнего мела Китая. Размером он такой же, как археоптерикс, а общий его план строения соответствует таковому у дромеозаврид, лишь череп у него более короткий и высокий, чем у крупных представителей семейства. Найденный дромеозаврид, образец включает отпечатки перьев в их естественном положении. На передних конечностях перья такие же, как у археоптерикса; рулевые перья крепятся лишь к задней половине длинного хвоста, но, что самое удивительное, голень также несла перья, похожие на второстепенные перья птиц, причем они были они ненамного меньше располагавшихся на предплечье. Эта находка сразу же заставила вспомнить о гипотезе перехода предков птиц к полету через древесную планирующую стадию, так как «ножные крылья» могли бы быть полезны при планировании, но никак не при машущем полете, и тем более не при сбивании взлетающих с земли насекомых. Реальному же использованию их для планирования коренным образом препятствует тот факт, что ни у какой птицы и ни у какого динозавра задние конечности не могут быть развернуты в стороны даже хотя бы примерно в горизонтальной плоскости. Остается лишь предположить, что эти перья могли использоваться для маневрирования на бегу или в прыжке, или выполняли сигнальную функцию, или развивались лишь потому, что не были вредны, будучи индуцированными к развитию теми же генами, что определяли развитие перьев на передних конечностях.
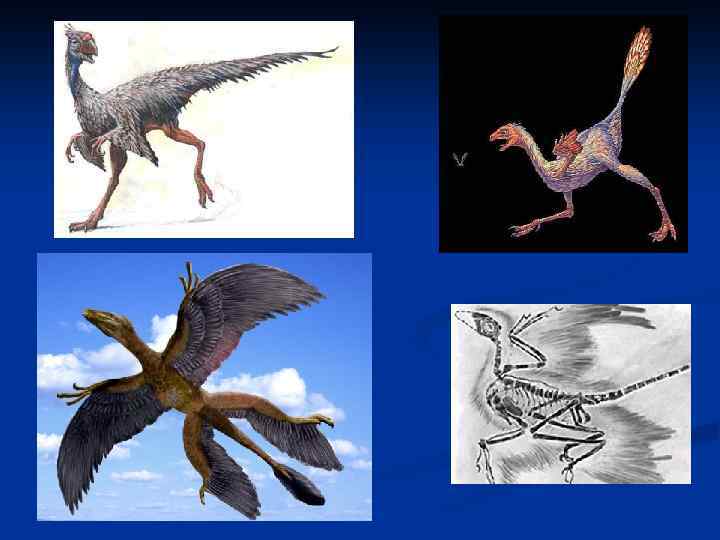

Почему же у предков птиц появились перья на хвосте? Потому, что при мелких размерах соотношение массы тела и его поверхности делает более удобным маневрирование путем отклонения воздушных потоков, а не путем смещения центра тяжести (как это делали с помощью хвоста крупные динозавры). Обособленное положение среди «первоптиц» занимают Alvaressaurus и Mononicuc из позднего мела Монголии. Это изящные тероподы «первоптиц» длиной около 1 м с длинным хвостом и необычным строением очень коротких, но сильных передних конечностей, обладавших лишь одним толстым и острым когтем. Примечательна окостеневшая грудина. Были ли у них перья, не известно, как не известно и то, прошли ли их необычные передние лапы стадию птичьих крыльев в своем эволюционном развитии, или нет. Что касается назначения этих конечностей, то они могли использоваться для выковыривания насекомых из гнилых бревен или термитников. Еще одно необычное животное, наделенное некоторыми птичьими признаками, при явной неспособности летать – двухметровый Avimimus также из позднего мела Монголии. На локтевой кости у него обозначены места крепления второстепенных перьев, а пястные кости слиты в «пряжку» . Вообще говоря, птичьи особенности появлялись в истории динозавров неоднократно и независимо. Любой динозавр имеет в своем строении те или иные из них (см. Лекцию 7), и даже вполне птичьего уровня организации достигли независимым путем, по крайней мере, две группы. Настоящих же птиц отличает окостеневающая у взрослых особей грудина с килем (или он утрачивается вторично), соединение коракоидов с грудиной суставом и изменение их конфигурации, позволяющее надкоракоидной мышце поднимать плечо, а не опускать, окостеневающие стернальные части ребер и сильно укороченный хвост. Отряд Enantiornithiformes считается более близко родственным археоптериксу, чем современным птицам, хотя представители его обладают всеми вышеназванными птичьими особенностями. Способность к активному полету, похоже, приобретена у них независимым путем. Увеличение объема грудных мышц достигнуто не столько за счет увеличения высоты киля, сколько за счет удлинения коракоидов. Хвост коракоидов. укорочен, но пигостиль в привычном его виде отсутствует. Энанциорнисы образовали в мелу ряд разнообразных форм, экологически конвергентных современным птицам. Среди них были и насекомоядные, и зерноядные с клювами, как у вьюрковых. Все остальные как ископаемые, так и современные птицы относятся к монофилетической группе веерохвостых. Два меловых отряда интересны сохранением у них зубов. Зубы располагались на зубной и верхнечелюстной костях, но отсутствовали на предчелюстной, как у предчелюстной, продвинутых птиц. Hesperornithiformes из Америки – высокоспециализированные, крупные (1 м), нелетающие ныряющие птицы. Их крылья сильно редуцированы, грудины без киля, а кости не пневматичны. Особенности захоронения свидетельствуют о колониальном образе жизни. пневматичны. Ichthyornithiformes также представлены в мелководных морских отложениях Северной Америки. Они имели хорошо развитый летный аппарат и по образу жизни, видимо, соответствовали современным чайкам. Из современных отрядов из мела известны только два: буревестникообразные (притом мелкие, не конкурировавшие с последними птерозаврами) и ржанкообразные. Высказывалось мнение, что палеогнатическое нёбо современных бескилевых птиц может быть следствием общей эмбрионализации всего организма взрослой особи в связи с эмбрионализацией строения летного аппарата. Но теперь известно, что в мелу и палеоцене существовала богатая фауна палеогнатических птиц, несомненно способных к полету. Укороченным хвостом эти птицы напоминают как бескилевых, так и современные формы вроде перепелов, у которых быстрый взлет на небольшое расстояние является в основном лишь способом бегства от наземных хищников. (Следует отметить, что тинамуобразные также сочетают палеогнатическое нёбо со способностью летать).


Несколько отрядов бескилевых птиц вымерли совсем недавно. Dinornithiformes обитали на Новой Зеландии, известны только с плейстоцена и исчезли в середине 2 -го тысячелетия н. э. У них были короткие цевки с четырьмя пальцами на них и неуклюжее телосложение, так как в отсутствие крупных наземных хищников им не было нужды быстро бегать. Кости крыла и плечевого пояса были полностью редуцированы. Существовало множество родов и видов этих птиц разного размера: от животных размером с индюка до Dinornis, достигавшего 3 м в высоту. Dinornis, Остатки динорнисообразных (моа) широко представлены в кухонных отходах аборигенов Новой Зеландии и в ямах в пещерах, куда эти (моа) неуклюжие животные иногда провались. В последних случаях иногда обеспечивалась высокая сохранность остатков (известны мумифицированные мышцы, связки, сухожилия и кожа с перьями разной окраски у разных видов: бурой, белой или поперечно-полосатой). Моа питались растительной пищей, и конкурентов у них среди млекопитающих не было. Среди современных птиц ближайшими родственниками моа выглядят киви. Aepyornithiformes известны из плейстоцена и голоцена (также до середины второго тысячелетия н. э. ) Мадагаскара. У них также были четырехпалые ноги, по росту они уступали моа, но имели более плотное телосложение и могли достигать массы 500 кг. Рудименты крыльев у моа, них сохранялись. Находки яиц этих птиц - самых крупных яиц в истории животных (объем 9 л) в болотах - заставляет думать, что эпиорнисы вели образ жизни гигантских пастушковых. Гигантские нелетающие птицы возникли и в отряде курообразных. Михиранги из плейстоцена Австралии соперничали по величине со курообразных. страусами. Страусообразные – единственные современные бескилевые, происхождение которых можно связать с неогнатическими птицами. Ergilornithidae из олигоцена Евразии представляют собой крупных нелетающих журавлеобразных с сильно редуцированным блоком второго пальца на цевке, что иллюстрирует возможный переход к двупалости у страусов. Известны и другие гигантские нелетающие журавлеобразные. Phorusrhacidae, или Phororhacidae, известны из олигоцена и плиоцена Америки. Они относятся к подотряду кариам и журавлеобразные. Phorusrhacidae, Phororhacidae, достигали в высоту до 3 м, а их огромные (до 0, 5 м длиной) черепа были снабжены мощным крючковатым клювом, похожим на клюв орла. Эти птицы были в целом довольно изящно сложенными, быстро бегающими хищниками. Diatrymidae из палеоцена и эоцена Северной Америки и Евразии иногда выделяются в собственный отряд Diatrymiformes. Они также имели крупные головы с крючковатыми клювами, но Diatrymiformes. их ноги короче и толще. Высказывалось предположение, что они не были хищниками, а своими клювами выкапывали из земли клубни, но это сомнительно. Семейства Graculavidae и Presbyornithidae, известные из позднего мела по эоцен, связывают между собой отряды ржанкообразных и Presbyornithidae, гусеобразных. У пресбиорниса куличье телосложение, но череп утки. От подобного рода птиц наверняка возникли и фламингообразные, так гусеобразных. фламингообразные, же, как и утиные, специализировавшиеся к фильтрации пищи из воды, но путем иной перестройки клюва. Современное семейство Cathartidae, формально относимое к соколообразным, в действительности, более родственно аистам. Ранние Cathartidae, соколообразным, представители близкого к катартидам семейства Teratornithidae, известного с миоцена по плейстоцен Америки, еще более похожи на аистов Teratornithidae, своими относительно длинными ногами. К тераторнитидам относится самое тяжелое из известных летающих животных и самая крупная летающая птица – Argentavis из миоцена с размахом крыльев до 7, 5 м и массой до 120 кг. Следует отметить, что эти величины гораздо больше теоретически рассчитанных предельных величин для летающих позвоночных. У тераторнитид ветви нижней челюсти не срастались в симфизе, что позволяло им заглатывать крупные пищевые объекты. Несколько видов тераторнитид представлены огромным количеством особей, наряду с другими хищными птицами, из плейстоценовых асфальтовых озер юга Северной Америки. В этих озерах, стремясь к воде, увязали многочисленные млекопитающие, а полакомиться их трупами прилетали и птицы, и если во время еды они дрались, то тоже часто падали в асфальт. Среди семейств отряда пеликанообразных раньше всех (с эоцена) известны ископаемые фрегатовые, причем они очень мало отличаются от фрегатовые, современного рода. Семейство Pelagornithidae включало гигантских птиц – парителей с размахом крыльев 5 – 6 м и общими пропорциями олуш. Длинные клювы несли множество острых зубоподобных костных выростов, а подвижность плечевого сустава была ограничено так, что крыло не могло подниматься выше горизонтального положения. Это делало пелагорнитид не способными к длительному машущему полету. Эти птицы, известные с эоцена по миоцен, жили в Европе, Северной и Южной Америки, Антарктиде, Новой Зеландии и Японии.


Plotopteridae, нелетающие ныряющие птицы из олигоцена и миоцена североамериканского и японского побережья Тихого океана, достигали Plotopteridae, в длину 2 м. Их ноги, несомненно, использовались для гребли, так как пропорции пальцев похожи на бакланьи, но сохранение киля указывает и на гребную функцию крыльев, как у пингвинов. Вымирание этих птиц могло быть связано с бурной радиацией тюленей и дельфинов. Этим же можно объяснить и отсутствие в современной фауне таких крупных пингвинов, как миоценовый представитель этого отряда, достигавший в высоту 1, 5 м. Представители отряда Pterosauria перешли к летающему образу жизни раньше, чем птицы. Строение их летного аппарата совершенно иное. Грудина, как у птиц, несла киль, но лишь в передней части грудины киль представлял собою костный вырост, а сзади он, видимо, продолжался в виде хряща. Коракоид и лопатка сращены, что у птиц бывает очень редко. Ключицы отсутствуют. Локтевая и лучевая кости неподвижны друг относительно друга, в противоположность этим костям у птиц; у их дистального конца крепился птероид – маленькая уникальная для этого отряда кость, поддерживавшая переднюю крыловую перепонку. Линия сгибания запястного сустава проходила между проксимальной и дистальной костями запястья, или несколькими аналогичными костями у примитивных представителей. Четвертая пястная кость сильно увеличена и создавала опору для удлиненного пальца, к которому крепилась летательная перепонка. Судя по отпечаткам перепонки, она содержала коллагеновые волокна, располагавшиеся в тех же направлениях, что и пальцы летучих мышей и перья птиц. Некоторые образцы показывают, что птерозавры были, видимо, покрыты тонкой короткой шерстью. Первая, вторая и третья пястные кости были маленькие, как и соответствующие им пальцы, снабженные острыми когтями и использовавшиеся, видимо, при наземной локомоции. Пятый палец и пятая пястная кость бесследно утрачены. Образцы с сохранившимися отпечатками кожи показывают, что задние конечности в летательную перепонку у большинства родов не включались, зато у полуводных форм сами несли плавательную перепонку между пальцами. Кости птерозавров полые и, судя по отверстиям, пневматизировались воздушными мешками. Сходное с птицами преобразование дыхательной системы свидетельствует и о сходном уровне метаболизма. Строение головного мозга, известное по отливкам мозговой полости, очень похоже на птичье. Череп птерозавров всегда очень крупный (по крайней мере, не меньше туловища), представляющий собой легкий каркас вокруг крупных отверстий. Квадратная кость подвижна. Положение задних конечностей птерозавров полностью улучшенное, головка бедреной кости отогнута медиально. Вертлужная впадина не медиально. прободена; имеется предлобковая кость. Малая берцовая кость редуцирована до стерженька, слитого с большой берцовой и не доходящего прободена; до ее дистального конца. Имеется мезотарсальный (внутризаплюсневый) сустав. Дистальные заплюсневые кости и плюсневые кости (внутризаплюсневый) сохраняют полную самостоятельность. Птерозавры подразделяются на два подотряда. Rhamphorhynchoidea существовали с позднего триаса по позднюю юру. Они имели длинных хвост, поддерживаемый окостеневающими сухожилиями, которые развивались как сильно удлиненные отростки пре– и постзигапофизов и гемапофизов, как у дромеозаврид. На конце гемапофизов, дромеозаврид. хвоста находилась кожистая рулевая лопасть. Грудные позвонки сохраняли самостоятельность. Четвертая пястная кость и четвертый палец кисти удлинены не очень сильно, то есть крылья рамфоринхоидов никогда не были очень длинными. Задние конечности относительно длинные, пятый палец на них удлинен и на ранних стадиях эволюции мог поддерживать отдельную перепонку, если он только не выполнял какой-либо другой функции. Pterodactyloidea существовали с поздней юры по поздний мел. Хвост у них очень короткий, и в компенсацию этого на голова часто бывали гребни, которые могли выполнять функцию руления. Ноги в среднем короче, а крылья иногда гораздо длиннее, чем у рамфоринхоидов. Имеется спинная кость, образованная сросшимися грудными позвонками; с ней сочленялся конец лопатки. Многие птеродактилоиды утрачивали зубы. В позднем мелу встречаются одни лишь крупные птерозавры (до 12 м в размахе крыльев). Видимо, это связано с конкуренцией со стороны птиц, которые долго не образовывали крупных форм. Летный аппарат птерозавров имел коренной недостаток по сравнению с таковым у птиц и летучих мышей: перепонка поддерживалась лишь одним пальцем и один ее разрыв лишал животное способности летать; кроме того, такое крыло всегда однообразно по форме, тогда как у птиц и летучих мышей благодаря разной относительной длине отдельных перьев или пальцев возможны разные варианты конфигурации крыльев в зависимости от образа жизни.

Подробности происхождения птерозавров неизвестны, так как промежуточных форм не найдено. Один их признак – сохранение крючковидной пятой плюсневой кости – отражает более примитивный уровень, чем уровень развития лагозухид и динозавров. Вместе с тем, полностью улучшенное положение задних конечностей и наличие в них внутризаплюсневого сустава указывает на более близкое родство с лагозухидами, чем с любой другой хорошо известной группой текодонтов. Каким образом птерозавры приобрели способность летать, гораздо менее понятно, чем в случае с птицами. У птерозавров, как у птиц, нет признаков сходства в строении скелета с планирующими позвоночными наподобие летяг, однако гораздо меньшая степень специализации задних конечностей к бегу, чем у динозавров и птиц, делает маловероятным переход к полету через использование передних конечностей для увеличения маневренности и устойчивости прыжках или для захвата подвижной добычи.
Лекция 6 (Летающие архозавры ).ppt