

Lektsia_5_Kholodovoy_stress_i_proch.pptx
- Количество слайдов: 22
 Лекция 5. Адаптация к низким температурам и ее молекулярные механизмы. Взаимодействия растений с бактериями и грибами: выработка защитных реакций. Подготовила Вера Абрамова 2 курс магистратуры Специальность «Биотехнология»
Лекция 5. Адаптация к низким температурам и ее молекулярные механизмы. Взаимодействия растений с бактериями и грибами: выработка защитных реакций. Подготовила Вера Абрамова 2 курс магистратуры Специальность «Биотехнология»
 Адаптация к низким температурам и ее молекулярные механизмы
Адаптация к низким температурам и ее молекулярные механизмы
 Сенсоры низкой температуры • Адаптация к холодовому стрессу является очень важным свойством растений. Возможные сенсоры низкой температуры в клетке: • 1)снижение подвижности мембран; • 2) интенсивность работы светособирающих систем хлоропластов не снижается, но снижена интенсивность работы цикла Кальвина.
Сенсоры низкой температуры • Адаптация к холодовому стрессу является очень важным свойством растений. Возможные сенсоры низкой температуры в клетке: • 1)снижение подвижности мембран; • 2) интенсивность работы светособирающих систем хлоропластов не снижается, но снижена интенсивность работы цикла Кальвина.
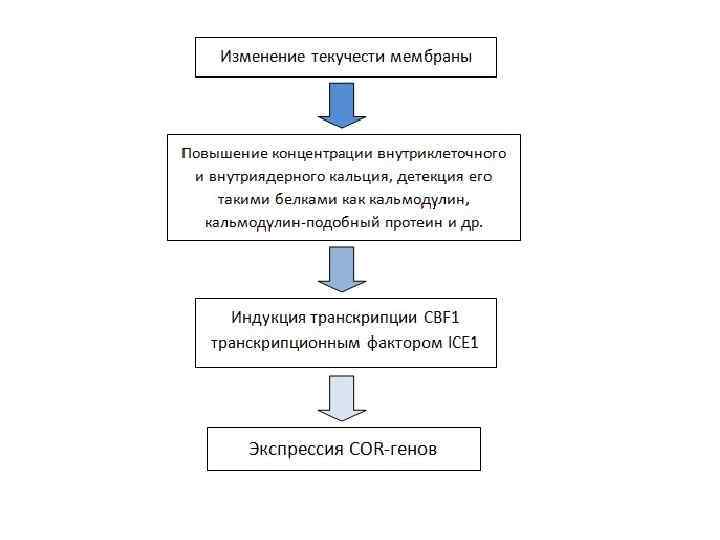
 Общий вид холодового ответа • Мембранная ригидификация вызывает накопление Са 2+ в цитоплазме, а также в ядре. В дальнейшем преобразовании этого сигнала задействованы Са 2+ сенсоры – Ca. M (calmodulin), CML (calmodulin-like), CDPK (Ca 2+ dependent protein kinase) и другие. • В результате повышается экспрессия генов COR cold responsive genes. Это происходит за счет активности транскрипционного фактора CBF- C repeat binding factors. • CBF 1/DREB 1 связываются в промоторной области COR-генов с последовательностью A/GCCGAC.
Общий вид холодового ответа • Мембранная ригидификация вызывает накопление Са 2+ в цитоплазме, а также в ядре. В дальнейшем преобразовании этого сигнала задействованы Са 2+ сенсоры – Ca. M (calmodulin), CML (calmodulin-like), CDPK (Ca 2+ dependent protein kinase) и другие. • В результате повышается экспрессия генов COR cold responsive genes. Это происходит за счет активности транскрипционного фактора CBF- C repeat binding factors. • CBF 1/DREB 1 связываются в промоторной области COR-генов с последовательностью A/GCCGAC.
 • Фермент Е 1 активирует С-конец убиквитина, используя энергию АТФ, аденилированием и переносом тиоэфирной связи с цистеина на E 1. Фермент Е 1 переносит убиквитин к транспортному ферменту Е 2, который при помощи Е 3 лигазы присоединяет его к субстрату.
• Фермент Е 1 активирует С-конец убиквитина, используя энергию АТФ, аденилированием и переносом тиоэфирной связи с цистеина на E 1. Фермент Е 1 переносит убиквитин к транспортному ферменту Е 2, который при помощи Е 3 лигазы присоединяет его к субстрату.
 u(biquitin-related) mo(difier)") SUMO=s(mall) u(biquitin-related) mo(difier)
SUMO=s(mall) u(biquitin-related) mo(difier)
 Деградация протеинов в протеасоме
Деградация протеинов в протеасоме
 Регуляция холодового ответа • Выше CBF по иерархии находится ICE 1 - inducer of CBF expression. Количество ICE 1 также регулируется. Наиболее хорошо описаны посттрансляционные модификации, прямо влияющие на его стабильность. • Убиквитинилирование, опосредованное HOS 1 приводит к деградации ICE 1 в протеасоме, сумоилирование, опосредованное SIZ 1, противостоит первой модификации и приводит к стабилизации уровня ICE 1.
Регуляция холодового ответа • Выше CBF по иерархии находится ICE 1 - inducer of CBF expression. Количество ICE 1 также регулируется. Наиболее хорошо описаны посттрансляционные модификации, прямо влияющие на его стабильность. • Убиквитинилирование, опосредованное HOS 1 приводит к деградации ICE 1 в протеасоме, сумоилирование, опосредованное SIZ 1, противостоит первой модификации и приводит к стабилизации уровня ICE 1.
 Посттрансляционная регуляция ICE 1
Посттрансляционная регуляция ICE 1
 Изменения уровня гормонов растений при холодовом стрессе • Уровень абсцизовой кислоты повышается • Уровень гиббереловой кислоты снижается. При холоде повышается уровень катаболизирующего гиббереллин фермента – ГА 2 -оксидазы, и снижается уровень вовлеченного в синтез ферменты – ГА 20 оксидазы. • Уровень салициловой кислоты повышается
Изменения уровня гормонов растений при холодовом стрессе • Уровень абсцизовой кислоты повышается • Уровень гиббереловой кислоты снижается. При холоде повышается уровень катаболизирующего гиббереллин фермента – ГА 2 -оксидазы, и снижается уровень вовлеченного в синтез ферменты – ГА 20 оксидазы. • Уровень салициловой кислоты повышается
 Индукция десатураз • В ответе растений на холодовой стресс важную роль играет индукция экспрессии десатураз – необходимо увеличить количество жирных кислот с ненасыщенными связями, что увеличит текучесть мембран при холоде
Индукция десатураз • В ответе растений на холодовой стресс важную роль играет индукция экспрессии десатураз – необходимо увеличить количество жирных кислот с ненасыщенными связями, что увеличит текучесть мембран при холоде
 Взаимодействия растений с бактериями и грибами: выработка защитных реакций
Взаимодействия растений с бактериями и грибами: выработка защитных реакций
 Отличия иммунитета растений и животных?
Отличия иммунитета растений и животных?
 • Растения не имеют в отличие от животных, специализированных клеток иммунной системы, в т. ч. адаптивную иммунную систему. Растения полагаются на неспецифический иммунитет каждой клетки и на системный ответ. • Типы ответа растения на патогены • PTI- PAMP triggered immunity • ETI-Effector-triggered immunity
• Растения не имеют в отличие от животных, специализированных клеток иммунной системы, в т. ч. адаптивную иммунную систему. Растения полагаются на неспецифический иммунитет каждой клетки и на системный ответ. • Типы ответа растения на патогены • PTI- PAMP triggered immunity • ETI-Effector-triggered immunity
 составляют первый уровень неспецифического иммунитета") PAMP-triggered immunity • Ассоциированные с патогенами молекулярные паттерны (РАМР) составляют первый уровень неспецифического иммунитета растений, который называется запущенный РАМР иммунитет (PTI- PAMP triggered immunity). Генетические и биохимические исследования идентифицировали паттернузнающие рецепторы PRR, вовлеченные в узнавание бактерий, грибов, оомицетов. • У животных ключевую роль в неспецифическом ответе играют Toll-like receptors (TLRs), разные виды которых экспрессируются на плазматической мембране или являются внутриклеточными рецепторами. PRR имеют структурные сходства с Toll-рецепторами дрозофилы и млекопитающих. • Запускаемый РАМР иммунный ответ растений представляет первую линию защиты и называется PTI.
PAMP-triggered immunity • Ассоциированные с патогенами молекулярные паттерны (РАМР) составляют первый уровень неспецифического иммунитета растений, который называется запущенный РАМР иммунитет (PTI- PAMP triggered immunity). Генетические и биохимические исследования идентифицировали паттернузнающие рецепторы PRR, вовлеченные в узнавание бактерий, грибов, оомицетов. • У животных ключевую роль в неспецифическом ответе играют Toll-like receptors (TLRs), разные виды которых экспрессируются на плазматической мембране или являются внутриклеточными рецепторами. PRR имеют структурные сходства с Toll-рецепторами дрозофилы и млекопитающих. • Запускаемый РАМР иммунный ответ растений представляет первую линию защиты и называется PTI.
 распознает главный эпитоп флагеллина fgl 22.") Рецепторы PTI • FLAGELLIN SENSING 2 (FLS 2) распознает главный эпитоп флагеллина fgl 22. Рецептор расположен на плазматической мембране и является важным компонентом иммунитета растений против патогенных бактерий. FLS 2 имеет характерную для трансмембранных киназ структуру с рецепторным эктодоменом, состоящим из лейцин богатых повторов (LRR) и активного внутриклеточного киназного домена. При связывании fgl 2 FLS 2 димеризуется с регуляторной киназой LRR-рецептора, BRI 1 ассоциированной киназой 1, которая в свою очередь запускает сигнальный каскад.
Рецепторы PTI • FLAGELLIN SENSING 2 (FLS 2) распознает главный эпитоп флагеллина fgl 22. Рецептор расположен на плазматической мембране и является важным компонентом иммунитета растений против патогенных бактерий. FLS 2 имеет характерную для трансмембранных киназ структуру с рецепторным эктодоменом, состоящим из лейцин богатых повторов (LRR) и активного внутриклеточного киназного домена. При связывании fgl 2 FLS 2 димеризуется с регуляторной киназой LRR-рецептора, BRI 1 ассоциированной киназой 1, которая в свою очередь запускает сигнальный каскад.
 • Другим самым охарактеризованным лигандом для рецепторов иммунитета растений является фактор") Рецепторы PTI (продолжение) • Другим самым охарактеризованным лигандом для рецепторов иммунитета растений является фактор трансляции EF-Tu бактерий. Лигандом является консервативный N-концевой ацетилированный участок. EFTu имеет все характеристики РАМР: имеется в большом количестве, необходим для выживания патогена. Как внутриклеточный EF-Tu становится видимым для растений, неясно. • GBP (glucan binding protein) бобовых, имеющий сайт связывания для 1, 6 -β- и 1, 3 -β-разветвленных гликозидов, представленных в клеточной стенке оомицета Phytophtora. Сигнальной трансдукции, следующей за связыванием лиганда, не отмечено. Связывающий хитин протеин CEBi. P был идентифицирован в рисе.
Рецепторы PTI (продолжение) • Другим самым охарактеризованным лигандом для рецепторов иммунитета растений является фактор трансляции EF-Tu бактерий. Лигандом является консервативный N-концевой ацетилированный участок. EFTu имеет все характеристики РАМР: имеется в большом количестве, необходим для выживания патогена. Как внутриклеточный EF-Tu становится видимым для растений, неясно. • GBP (glucan binding protein) бобовых, имеющий сайт связывания для 1, 6 -β- и 1, 3 -β-разветвленных гликозидов, представленных в клеточной стенке оомицета Phytophtora. Сигнальной трансдукции, следующей за связыванием лиганда, не отмечено. Связывающий хитин протеин CEBi. P был идентифицирован в рисе.
 ETI- effector triggered immunity • Чтобы избежать PTI, патогены могут запускать эффеторные молекулы внутрь клетки, как например, поступает Ps. syringae. Внутриклеточными рецепторами растений являются R-протеины. Запускаемые ими ответ называется ETI-effector triggered immunity или опосредованная R-генами устойчивость. Сверхчувствительная реакция есть ключевой признак ETI. Он приводит к гибели клетки и продукции антимикробных молекул – гидролитических энзимов хитиназы и β-1, 3 - глюканазы в окружающей ткани. • R-протеины состоят из вариабельного N-конца, нуклеотидсвязывающего сайта и лейцин-богатого повтора. • Активирование R-протеинов начинается, когда свои протеины оказываются затронуты или модифицированы эффекторами патогена. RIN 4 -RPMI-interacting protein – самый изученный целевой белок для защиты R-протеинами. При попадании эффекторов патогена в клетку (Avr), RIN 4 фосфорилируется (т. е. , видоизменяется) и активирует R-протеины.
ETI- effector triggered immunity • Чтобы избежать PTI, патогены могут запускать эффеторные молекулы внутрь клетки, как например, поступает Ps. syringae. Внутриклеточными рецепторами растений являются R-протеины. Запускаемые ими ответ называется ETI-effector triggered immunity или опосредованная R-генами устойчивость. Сверхчувствительная реакция есть ключевой признак ETI. Он приводит к гибели клетки и продукции антимикробных молекул – гидролитических энзимов хитиназы и β-1, 3 - глюканазы в окружающей ткани. • R-протеины состоят из вариабельного N-конца, нуклеотидсвязывающего сайта и лейцин-богатого повтора. • Активирование R-протеинов начинается, когда свои протеины оказываются затронуты или модифицированы эффекторами патогена. RIN 4 -RPMI-interacting protein – самый изученный целевой белок для защиты R-протеинами. При попадании эффекторов патогена в клетку (Avr), RIN 4 фосфорилируется (т. е. , видоизменяется) и активирует R-протеины.
 Реакция сверхчувствительности
Реакция сверхчувствительности
 Механизмы борьбы растений с инфекцией грибами • Защита растений против грибов заключается в основном в изменении концентрации метаболитов, энзимов внутри и вне клеток. • Важную роль играет реакция сверхчувствительности и респираторный взрыв. • При инфекции грибом в растении поизводятся нерпротеиновые субстанции –алкалоиды, сапонины, дигидрофенолы, танины, хиноны и ароматические аминокислоты. • Среди протеиновых компонентов защиты важны экстесины (богаты гидроксипролином), которые составляют основу для образования клеточной стенки в новом месте, глицин-богатые протеины и PR-протеины. Среди PR протеинов большую часть занимают β 1, 3 -глюканазы, пероксидаза, каталаза.
Механизмы борьбы растений с инфекцией грибами • Защита растений против грибов заключается в основном в изменении концентрации метаболитов, энзимов внутри и вне клеток. • Важную роль играет реакция сверхчувствительности и респираторный взрыв. • При инфекции грибом в растении поизводятся нерпротеиновые субстанции –алкалоиды, сапонины, дигидрофенолы, танины, хиноны и ароматические аминокислоты. • Среди протеиновых компонентов защиты важны экстесины (богаты гидроксипролином), которые составляют основу для образования клеточной стенки в новом месте, глицин-богатые протеины и PR-протеины. Среди PR протеинов большую часть занимают β 1, 3 -глюканазы, пероксидаза, каталаза.
 Источники • • • Холодовой ответ Steven Penfield. Temperature perception and signal transduction in plants//New Phytologist (2008) 179: 615– 628. http: //www. cellbiol. ru/book/kletka/proteasomy http: //biomolecula. ru/content/1226/ http: //www. news. leiden. edu/news/wrestling-with-sumo-proteins. html http: //www. labome. ru/method/Proteasome-Inhibitors. html Иммунитет растений Zipfel C. Pattern-recognition receptors in plant innate immunity// Curr. Opin. Immunol. 2008 Feb; 20(1): 10 -6 http: //en. wikipedia. org/wiki/Plant_disease_resistance#Effector_triggered_immunity http: //www. mnf. uni-tuebingen. de/fachbereiche/zentren/zmbp/plant-biochemistry/researchgroups/felix. html Robatzek S. and Wirthmueller L. Mapping FLS 2 function to structure: LRRs, kinase and its working bits//Protoplasma 2013 Jun; 250(3): 671 -81 http: //www. apsnet. org/edcenter/illglossary/Article%20 Images/Forms/Disp. Form. aspx? ID=519
Источники • • • Холодовой ответ Steven Penfield. Temperature perception and signal transduction in plants//New Phytologist (2008) 179: 615– 628. http: //www. cellbiol. ru/book/kletka/proteasomy http: //biomolecula. ru/content/1226/ http: //www. news. leiden. edu/news/wrestling-with-sumo-proteins. html http: //www. labome. ru/method/Proteasome-Inhibitors. html Иммунитет растений Zipfel C. Pattern-recognition receptors in plant innate immunity// Curr. Opin. Immunol. 2008 Feb; 20(1): 10 -6 http: //en. wikipedia. org/wiki/Plant_disease_resistance#Effector_triggered_immunity http: //www. mnf. uni-tuebingen. de/fachbereiche/zentren/zmbp/plant-biochemistry/researchgroups/felix. html Robatzek S. and Wirthmueller L. Mapping FLS 2 function to structure: LRRs, kinase and its working bits//Protoplasma 2013 Jun; 250(3): 671 -81 http: //www. apsnet. org/edcenter/illglossary/Article%20 Images/Forms/Disp. Form. aspx? ID=519