

Лекция 4.ppt_Биология
- Количество слайдов: 14
 Лекция 4 Ископаемые пресмыкающиеся
Лекция 4 Ископаемые пресмыкающиеся
 Рептилии – весьма неопределенное понятие, отражающее не степень родства, а уровень организации. Рептилии – это такие амниоты, которые не птицы и не звери. Но как узнать, относится ли ископаемое животное к амниотам? Из палеозоя никаких яиц неизвестно. Формально к примитивным рептилиям относили формы с хорошо развитыми, возникшими из плевроцентров телами позвонков (без хорды внутри), с маленькими (или отсутствующими) интерцентрами и единственным затылочным мыщелком. Строение тела этих животных, таких как Seimouria и Diadectes, и высокая степень окостенения суставных поверхностей их костей, говорит о наземном образе жизни. Но теперь Diadectes, для животных подобного типа, относящихся к антракозаврам, известны водные личинки. Ввиду отсутствия возможности причислить этих антракозаврам, животных к амниотам, их, а также их потомков иногда относят к классу Parareptilia. Но нужно помнить, что амниотического яйца могло не Parareptilia. быть и у представителей других филетических линий, взрослые особи которых в конце карбона, подобно сеймуриаморфам и диадектидам, уже имели рептилийный облик. Парарептилии по строению черепа анапсиды. У сеймурии был всего один, как у амфибий, крестцовый анапсиды. позвонок, и зубной аппарат хищника; в длину она достигала 50 см. Достигавший в длину 2 м диадект был растительноядным. Важнейший признак, по которому отделяют бесспорных амниот от сомнительных – присутствие поперечного выступа крыловидной кости, на котором крепилась крыловидная мышца, развивающая максимальное усилие при широко раскрытом рте. У амфибий крыловидная мышца недифференцирована. Древнейшие животные, соответствующие этому критерию – Hylonomus и Paleothyris из карбона, недифференцирована. относящиеся к семейству Protorothyridae отряда Captorhinida. Они сохранились не в обычных отложениях каменноугольных болот, как Captorhinida. большинство четвероногих сходного возраста, а внутри вертикально стоящих пней гигантских папоротникообразных растений. Это говорит об их истинно наземном образе жизни. По внешнему виду эти животные были похожи на мелких современных ящериц. Шейных позвонков четыре, крестцовых два. От животных с анатомией проторотирид можно вывести и всех остальных амниот, и монофилия амниот признается многими авторами. Millerosauria – подотряд капторинид из средней и верхней перми Южной Африки. Они независимо от современных диапсид приобрели боковое височное окно, а наличие ушной вырезки свидетельствует и о наличии барабанной перепонки. По-видимому, миллерозавры – первые амниоты, приобретшие эту структуру, но независимо от других, так как у миллерозавров она крепилась на заднем крае чешуйчатой и квадратноскуловой костей, а не на квадратной. Миллерозавры также были мелкими ящерицеподобными животными. Видимо, настоящие ящерицы их и вытеснили. Подотряд Procolophonoidea известны с верхней перми по верхний триас и широко распространены по свету. Крестцовых ребер три. Могла существовать барабанная перепонка, устроенная по типу миллерозавров. Зубы мелкие, колышковидные, приспособленные к растительной миллерозавров. колышковидные, пище. Триасовые представители крупные, а щечная область черепа скульптурирована у них длинными защитными отростками. Подотряд Pareiasaroidea – самые крупные капториниды из средней и верхней перми Африки, Западной Европы, России и Китая. Они достигали в длину трех метров. Их короткие и толстые лапы, в отличие от других примитивных амниот, были поставлены почти вертикально, чтобы поддерживать массивное туловище. Подобно млекопитающим, лопатка длинная, а лобковая и седалищная кости таза мелкие и повернутые назад позади удлиненной подвздошной. Такая конфигурация, возможно, облегчала использование мышц, двигавших заднюю ногу вперед-назад без отведения ее в сторону. Череп короткий, массивный, расширенный, скульптурированный, особенно в щечной скульптурированный, области. Зубы листовидные, приспособленные к растительной пище. В коже туловища залегали мелкие костные пластинки. Парейазавры были многочисленны, образовывали большие стада, пасшиеся в заросших растительностью болотах, и являлись излюбленной добычей крупных хищников того времени. Исчезновение их в конце перми было, видимо, связано с конкуренцией со стороны растительноядных терапсид. Mesosauria – подотряд капторинид, представляющий древнейших вторичноводных амниот из перми Южной Африки и Южной Америки. капторинид, Рыло сильно удлинено; зубы очень длинные и тонкие и, возможно, служили фильтровальным аппаратом для питания мелкими ракообразными. Шея длиннее, чем у примитивных капторинид, и включает 10 позвонков. Хвост длинный, уплощенный с боков, капторинид, приспособленный к гребле. Хвостовые позвонки, как у сухопутных капторинид, несут признаки способности автотомии. Стопа длинная, капторинид, веслообразная.
Рептилии – весьма неопределенное понятие, отражающее не степень родства, а уровень организации. Рептилии – это такие амниоты, которые не птицы и не звери. Но как узнать, относится ли ископаемое животное к амниотам? Из палеозоя никаких яиц неизвестно. Формально к примитивным рептилиям относили формы с хорошо развитыми, возникшими из плевроцентров телами позвонков (без хорды внутри), с маленькими (или отсутствующими) интерцентрами и единственным затылочным мыщелком. Строение тела этих животных, таких как Seimouria и Diadectes, и высокая степень окостенения суставных поверхностей их костей, говорит о наземном образе жизни. Но теперь Diadectes, для животных подобного типа, относящихся к антракозаврам, известны водные личинки. Ввиду отсутствия возможности причислить этих антракозаврам, животных к амниотам, их, а также их потомков иногда относят к классу Parareptilia. Но нужно помнить, что амниотического яйца могло не Parareptilia. быть и у представителей других филетических линий, взрослые особи которых в конце карбона, подобно сеймуриаморфам и диадектидам, уже имели рептилийный облик. Парарептилии по строению черепа анапсиды. У сеймурии был всего один, как у амфибий, крестцовый анапсиды. позвонок, и зубной аппарат хищника; в длину она достигала 50 см. Достигавший в длину 2 м диадект был растительноядным. Важнейший признак, по которому отделяют бесспорных амниот от сомнительных – присутствие поперечного выступа крыловидной кости, на котором крепилась крыловидная мышца, развивающая максимальное усилие при широко раскрытом рте. У амфибий крыловидная мышца недифференцирована. Древнейшие животные, соответствующие этому критерию – Hylonomus и Paleothyris из карбона, недифференцирована. относящиеся к семейству Protorothyridae отряда Captorhinida. Они сохранились не в обычных отложениях каменноугольных болот, как Captorhinida. большинство четвероногих сходного возраста, а внутри вертикально стоящих пней гигантских папоротникообразных растений. Это говорит об их истинно наземном образе жизни. По внешнему виду эти животные были похожи на мелких современных ящериц. Шейных позвонков четыре, крестцовых два. От животных с анатомией проторотирид можно вывести и всех остальных амниот, и монофилия амниот признается многими авторами. Millerosauria – подотряд капторинид из средней и верхней перми Южной Африки. Они независимо от современных диапсид приобрели боковое височное окно, а наличие ушной вырезки свидетельствует и о наличии барабанной перепонки. По-видимому, миллерозавры – первые амниоты, приобретшие эту структуру, но независимо от других, так как у миллерозавров она крепилась на заднем крае чешуйчатой и квадратноскуловой костей, а не на квадратной. Миллерозавры также были мелкими ящерицеподобными животными. Видимо, настоящие ящерицы их и вытеснили. Подотряд Procolophonoidea известны с верхней перми по верхний триас и широко распространены по свету. Крестцовых ребер три. Могла существовать барабанная перепонка, устроенная по типу миллерозавров. Зубы мелкие, колышковидные, приспособленные к растительной миллерозавров. колышковидные, пище. Триасовые представители крупные, а щечная область черепа скульптурирована у них длинными защитными отростками. Подотряд Pareiasaroidea – самые крупные капториниды из средней и верхней перми Африки, Западной Европы, России и Китая. Они достигали в длину трех метров. Их короткие и толстые лапы, в отличие от других примитивных амниот, были поставлены почти вертикально, чтобы поддерживать массивное туловище. Подобно млекопитающим, лопатка длинная, а лобковая и седалищная кости таза мелкие и повернутые назад позади удлиненной подвздошной. Такая конфигурация, возможно, облегчала использование мышц, двигавших заднюю ногу вперед-назад без отведения ее в сторону. Череп короткий, массивный, расширенный, скульптурированный, особенно в щечной скульптурированный, области. Зубы листовидные, приспособленные к растительной пище. В коже туловища залегали мелкие костные пластинки. Парейазавры были многочисленны, образовывали большие стада, пасшиеся в заросших растительностью болотах, и являлись излюбленной добычей крупных хищников того времени. Исчезновение их в конце перми было, видимо, связано с конкуренцией со стороны растительноядных терапсид. Mesosauria – подотряд капторинид, представляющий древнейших вторичноводных амниот из перми Южной Африки и Южной Америки. капторинид, Рыло сильно удлинено; зубы очень длинные и тонкие и, возможно, служили фильтровальным аппаратом для питания мелкими ракообразными. Шея длиннее, чем у примитивных капторинид, и включает 10 позвонков. Хвост длинный, уплощенный с боков, капторинид, приспособленный к гребле. Хвостовые позвонки, как у сухопутных капторинид, несут признаки способности автотомии. Стопа длинная, капторинид, веслообразная.

 Анапсиды, известные в настоящее время – черепахи. Отдельные систематики, считая за их предков антракозавров семейства Discosauriscidae, Анапсиды, Discosauriscidae, относят и самих черепах к парарептилиям. Предки черепах достигли раптилийного уровня организации лишь в середине перми. Таким парарептилиям. перми. образом, это самая молодая группа современных пресмыкающихся. Древнейшая известная черепаха – Proganochelys из позднего триаса Германии. Длина ее – 1 м. Голова, ноги и хвост не убирались под панцырь и были защищены шипами. Краевые зубы уже утрачены, а небные еще имелись. Носовые кости сохранились, а пинеальное отверстие в крыше черепа уже исчезло. Наружный аддуктор проходил напрямую от нижней челюсти к черепу, тогда как у скрытошейных черепах он перекидывается через передний выступ слуховой капсулы, а у бокошейных – через отросток крыловидной кости. Эти преобразования связаны с увеличением объема ушной полости. Лопатка проганохелиса еще не приобрела типичной для современных видов двуветвистой формы, в плечевом поясе сохраняются клейтрум, ключица и межключица. Конструкция панциря похожа на таковую клейтрум, межключица. современных черепах, но включает утраченные позже костные пластинки. Крестцовый позвонок один (у современных черепах два). Проганохелис относится к подотряду Amphychelydia, просуществовавшему до эоцена. Amphychelydia, Подотряд Pleurodira был в прошлом широко распространен. В то время как все ныне живущие бокошейные черепахи – пресноводные, к вымершим относятся и высокотелые сухопутные формы, и морские. К подотряду Cryptodira, богато представленному в современной фауне, относится семейство Meiolaniidae из плейстоцена Австралии. Они Cryptodira, достигали размеров коровы, имели рога, и вели наземный образ жизни. По размерам это не предел – представители современного рода Testudo в плиоцене у подножья Гималаев достигали веса 1 тонны. Очень крупными были и некоторые морские скрытошейные. Меловой скрытошейные. Archelon с ластообразными конечностями имел в длину 4 м и ажурную, как у современной кожистой черепахи, костную основу панциря. Eunotosaurus из средней перми Южной Африки относится к отдельному подотряду капторинид и имеет некоторые общие с черепахами особенности. Ребра его сильно расширены, как у черепах, а туловищных позвонков всего 10. Но следов кожного панциря нет, а плечевой пояс находился снаружи от ребер. Наличие пластиновидного затылка, по-видимому, исключало развитие аддукторной челюстной мускулатуры черепашьего типа. У черепах аддукторы челюсти распространяются назад и на затылок через задневисочные вырезки (функциональное подобие височных окон). Древнейший известный диапсид – Petrolacosaurus из карбона Канзаса, изящное ящерицеподобное, 20 – сантиметровое насекомоядное ящерицеподобное, существо. У него нет никаких признаков присутствия барабанной перепонки, тогда как ранние капториноморфы уже обладали ушной вырезкой. Таким образом, барабанная перепонка диапсид и анапсид возникла независимо. Близкий его родственник – нижнепермский Araeoscelis без нижнего височного окна. Возможно, оно закрылось вторично для укрепления черепа при более мощном укусе. Отряд Araeoscelida, к которому относятся оба рода, можно считать сестринской группой всех более поздних диапсид. Тому же уровню радиации Araeoscelida, диапсид. принадлежит Coelurosauravus из верхней перми Мадагаскара, чьи ребра использовались для планирования как у современного Draco, а Draco, нижняя скуловая дуга была утрачена; и водный отряд Thalattosauria из триаса Швейцарии и Северной Америки. Представителем последних является двухметровый Askeptosaurus с удлиненными телом, хвостом и шеей и смещенными далеко назад ноздрями. Ноги его короткие, но не преобразованы в плавники. Большинство остальных диапсид можно отнести к двум инфраклассам: Lepidosauromorpha и Archosauromorpha. инфраклассам: Archosauromorpha. У лепидозавроморф важный элемент локомоции – боковые изгибы тела. В связи с этим грудина у них хорошо развита, а коракоиды соединены с ней подвижно, увеличивая длину шага передних лап. Древнейшие лепидозавры Youngina и Thadeosaurus относятся к верхнепермскому отряду Eosuchia. Позвонки у них амфицельные; сохранен клейтрум, утраченный более прогрессивными лепидозаврами. Eosuchia. амфицельные; клейтрум, лепидозаврами. Помимо наземных, были и водные эозухии с веслоподобным хвостом, такие как Hovasaurus из верхней перми Мадагаскара. Семейство Champsosauridae, известное с мела по эоцен, примитивными признаками напоминает эозухий, но по специализированным сходно с Champsosauridae, эозухий, крокодилами: у них длинное, узкое рыло с ноздрями на конце верхней стороны; скуловые дуги раздвинуты в стороны, чтобы обеспечить крепление мощной челюстной мускулатуры.
Анапсиды, известные в настоящее время – черепахи. Отдельные систематики, считая за их предков антракозавров семейства Discosauriscidae, Анапсиды, Discosauriscidae, относят и самих черепах к парарептилиям. Предки черепах достигли раптилийного уровня организации лишь в середине перми. Таким парарептилиям. перми. образом, это самая молодая группа современных пресмыкающихся. Древнейшая известная черепаха – Proganochelys из позднего триаса Германии. Длина ее – 1 м. Голова, ноги и хвост не убирались под панцырь и были защищены шипами. Краевые зубы уже утрачены, а небные еще имелись. Носовые кости сохранились, а пинеальное отверстие в крыше черепа уже исчезло. Наружный аддуктор проходил напрямую от нижней челюсти к черепу, тогда как у скрытошейных черепах он перекидывается через передний выступ слуховой капсулы, а у бокошейных – через отросток крыловидной кости. Эти преобразования связаны с увеличением объема ушной полости. Лопатка проганохелиса еще не приобрела типичной для современных видов двуветвистой формы, в плечевом поясе сохраняются клейтрум, ключица и межключица. Конструкция панциря похожа на таковую клейтрум, межключица. современных черепах, но включает утраченные позже костные пластинки. Крестцовый позвонок один (у современных черепах два). Проганохелис относится к подотряду Amphychelydia, просуществовавшему до эоцена. Amphychelydia, Подотряд Pleurodira был в прошлом широко распространен. В то время как все ныне живущие бокошейные черепахи – пресноводные, к вымершим относятся и высокотелые сухопутные формы, и морские. К подотряду Cryptodira, богато представленному в современной фауне, относится семейство Meiolaniidae из плейстоцена Австралии. Они Cryptodira, достигали размеров коровы, имели рога, и вели наземный образ жизни. По размерам это не предел – представители современного рода Testudo в плиоцене у подножья Гималаев достигали веса 1 тонны. Очень крупными были и некоторые морские скрытошейные. Меловой скрытошейные. Archelon с ластообразными конечностями имел в длину 4 м и ажурную, как у современной кожистой черепахи, костную основу панциря. Eunotosaurus из средней перми Южной Африки относится к отдельному подотряду капторинид и имеет некоторые общие с черепахами особенности. Ребра его сильно расширены, как у черепах, а туловищных позвонков всего 10. Но следов кожного панциря нет, а плечевой пояс находился снаружи от ребер. Наличие пластиновидного затылка, по-видимому, исключало развитие аддукторной челюстной мускулатуры черепашьего типа. У черепах аддукторы челюсти распространяются назад и на затылок через задневисочные вырезки (функциональное подобие височных окон). Древнейший известный диапсид – Petrolacosaurus из карбона Канзаса, изящное ящерицеподобное, 20 – сантиметровое насекомоядное ящерицеподобное, существо. У него нет никаких признаков присутствия барабанной перепонки, тогда как ранние капториноморфы уже обладали ушной вырезкой. Таким образом, барабанная перепонка диапсид и анапсид возникла независимо. Близкий его родственник – нижнепермский Araeoscelis без нижнего височного окна. Возможно, оно закрылось вторично для укрепления черепа при более мощном укусе. Отряд Araeoscelida, к которому относятся оба рода, можно считать сестринской группой всех более поздних диапсид. Тому же уровню радиации Araeoscelida, диапсид. принадлежит Coelurosauravus из верхней перми Мадагаскара, чьи ребра использовались для планирования как у современного Draco, а Draco, нижняя скуловая дуга была утрачена; и водный отряд Thalattosauria из триаса Швейцарии и Северной Америки. Представителем последних является двухметровый Askeptosaurus с удлиненными телом, хвостом и шеей и смещенными далеко назад ноздрями. Ноги его короткие, но не преобразованы в плавники. Большинство остальных диапсид можно отнести к двум инфраклассам: Lepidosauromorpha и Archosauromorpha. инфраклассам: Archosauromorpha. У лепидозавроморф важный элемент локомоции – боковые изгибы тела. В связи с этим грудина у них хорошо развита, а коракоиды соединены с ней подвижно, увеличивая длину шага передних лап. Древнейшие лепидозавры Youngina и Thadeosaurus относятся к верхнепермскому отряду Eosuchia. Позвонки у них амфицельные; сохранен клейтрум, утраченный более прогрессивными лепидозаврами. Eosuchia. амфицельные; клейтрум, лепидозаврами. Помимо наземных, были и водные эозухии с веслоподобным хвостом, такие как Hovasaurus из верхней перми Мадагаскара. Семейство Champsosauridae, известное с мела по эоцен, примитивными признаками напоминает эозухий, но по специализированным сходно с Champsosauridae, эозухий, крокодилами: у них длинное, узкое рыло с ноздрями на конце верхней стороны; скуловые дуги раздвинуты в стороны, чтобы обеспечить крепление мощной челюстной мускулатуры.

 Отряды Чешуйчатые и Клинозубые образуют надотряд Lepidosauria в инфраклассе Lepidosauromorpha. У лепидозавров тонкое, Lepidosauromorpha. палочковидное стремя, позволяющее передавать колебания барабанной перепонки. Правда, у гаттерии сама перепонка отсутствует, но наверняка вторично. Редукция барабанной перепонки произошла также у змей и некоторых роющих ящериц. Стопа чешуйчатых и клинозубых имеет сходное строение: таранная, пяточная и центральная кости срастаются в одну (называемую также таранной), прочно прикрепленную к голени и подвижно сочленяющуюся с четвертой дистальной заплюсневой, а сухожилие икроножной мышцы крепится к заплюсневой, пятой плюсневой кости, имеющей крючковидную форму. Дивергенция чешуйчатых и клинозубых была связана с различиями в питании. У клинозубых аддукторная камера черепа крупная, что обеспечивает сильный медленный укус. Зубов немного и они акродонтные, незамещающиеся. Квадратная кость неподвижна, как у эозухий. У акродонтные, незамещающиеся. эозухий. ящериц аддукторная камера короткая, что обеспечивает быстрый, но слабый укус; этой быстроте и точности прицеливания, важной при захвате мелкой подвижной добычи, способствует подвижность квадратной кости, а зубы чаще плевродонтные и замещающиеся. Клинозубые адаптировались к оппортунистическому питанию, а ящерицы – к потреблению насекомых. Это и привело к тому, что в настоящее время существует лишь один вид клинозубых и более 3500 видов ящериц. Древнейшее клинозубое – Planocephalosaurus – известно из верхнего триаса Европы; оно совершенно сходно с гаттерией. С кайнозоя эта группа претерпела упадок. К клинозубым относится также водное семейство Pleurosauridae с уменьшенными ногами и неполной нижней скуловой дугой, известное с ранней юры до раннего мела. Ящерицы в лице южноафриканской Paliguana известны с верхней перми. Уже у нее утрачена нижняя скуловая дуга ради увеличения перми. подвижности квадратной кости. Появление стрептостилии связано с увеличением силы действия крыловидной мышцы при почти сомкнутых челюстях, что важно при охоте на мелкую добычу. Сочетание более эффективных слуха, челюстной механики и локомоции позволило ящерицам занять экологические ниши, ранее принадлежавшие саламандроподобным амфибиям, мелким анапсидам и эозухиям. Позвонки у эозухиям. ящериц процельные, кроме гекконов, у которых они вторично амфицельны. Самые прогрессивные ящерицы – надсемейство Varanoidea – процельные, гекконов, амфицельны. послужили предками змей, а также двух водных семейств, имеющий стрептогнатический внутричелюстной сустав: Aigialosauridae из верхней юры среднего мела имели малоспециализированные конечности и длину не более 1 м. Mosasauridae имели ластообразные гиперфалангические конечности (связь задних с позвоночником утрачена) и низкий хвостовой плавник из отростков верхних и нижних дуг позвонков. Задние туловищные ребра утрачены. Шея короткая для вараноидов (семь позвонков). У некоторых барабанная перепонка обызвествленная, видимо, чтобы выдерживать давление воды при нырянии. Зубы текодонтные – в лунках; у обызвествленная, рода Globidens круглые, для раскусывания аммонитов. Мозазавры достигали в длину 10 м. Беременные самки не обнаружены, а молодь редко встречается со взрослыми, - значит, мозазавры откладывали яйца на суше, что при их размерах и конструкции было непросто. Змеи произошли от вараноидов, отличаясь от них отсутствием грудины и конечности и наличием дополнительных суставных поверхностей вараноидов, позвонков – зигосфена спереди и зигантра сзади. Главный вопрос – были ли предки змей роющими или водными. Общие признаки с роющими ящерицами – удлинение тела, утрата ног, барабанной перепонки и полости среднего уха, сращение век, отсутствие склеральных колец. Но череп роющих ящериц и амфисбен монолитный и прочный, а у змей – ажурный. Даже у ранних змей высоко развит кинетизм черепа, при роющей жизни не выгодный. Утрата змеями барабанной перепонки может объясняться именно крайней подвижность квадратной кости, что делает невозможным ее прикрепление (равно как и сохранение скуловых дуг). Изменение глаза можно связать с ночным образом жизни и ползаньем (в том числе под водой) среди густой растительности. Недавно стали известны два верхнемеловых водных вараноида из Израиля, длинных и безногих: Ophiomorphus и Pachyrachis, годящихся на роль предков змей. Дальнейший успех змей Pachyrachis, был связан с радиацией млекопитающих, служивших им пищей. Известные с третичного периода амфисбены ничего не говорят о происхождении этой группы сверх известного от современных видов. Вероятнее всего, это группа равного змеям ранга, но их конкретные предки из числа ящериц неясны. Пресмыкающиеся часто и с легкостью возвращались к водному образу жизни – животному с низкой интенсивностью метаболизма не сложно задерживать дыхание. Судя по современным морским игуанам, питание и движение в воде не требует у рептилий структурной и физиологической адаптации. Водная локомоция требует у них ¾ энергетических затрат от сухопутной.
Отряды Чешуйчатые и Клинозубые образуют надотряд Lepidosauria в инфраклассе Lepidosauromorpha. У лепидозавров тонкое, Lepidosauromorpha. палочковидное стремя, позволяющее передавать колебания барабанной перепонки. Правда, у гаттерии сама перепонка отсутствует, но наверняка вторично. Редукция барабанной перепонки произошла также у змей и некоторых роющих ящериц. Стопа чешуйчатых и клинозубых имеет сходное строение: таранная, пяточная и центральная кости срастаются в одну (называемую также таранной), прочно прикрепленную к голени и подвижно сочленяющуюся с четвертой дистальной заплюсневой, а сухожилие икроножной мышцы крепится к заплюсневой, пятой плюсневой кости, имеющей крючковидную форму. Дивергенция чешуйчатых и клинозубых была связана с различиями в питании. У клинозубых аддукторная камера черепа крупная, что обеспечивает сильный медленный укус. Зубов немного и они акродонтные, незамещающиеся. Квадратная кость неподвижна, как у эозухий. У акродонтные, незамещающиеся. эозухий. ящериц аддукторная камера короткая, что обеспечивает быстрый, но слабый укус; этой быстроте и точности прицеливания, важной при захвате мелкой подвижной добычи, способствует подвижность квадратной кости, а зубы чаще плевродонтные и замещающиеся. Клинозубые адаптировались к оппортунистическому питанию, а ящерицы – к потреблению насекомых. Это и привело к тому, что в настоящее время существует лишь один вид клинозубых и более 3500 видов ящериц. Древнейшее клинозубое – Planocephalosaurus – известно из верхнего триаса Европы; оно совершенно сходно с гаттерией. С кайнозоя эта группа претерпела упадок. К клинозубым относится также водное семейство Pleurosauridae с уменьшенными ногами и неполной нижней скуловой дугой, известное с ранней юры до раннего мела. Ящерицы в лице южноафриканской Paliguana известны с верхней перми. Уже у нее утрачена нижняя скуловая дуга ради увеличения перми. подвижности квадратной кости. Появление стрептостилии связано с увеличением силы действия крыловидной мышцы при почти сомкнутых челюстях, что важно при охоте на мелкую добычу. Сочетание более эффективных слуха, челюстной механики и локомоции позволило ящерицам занять экологические ниши, ранее принадлежавшие саламандроподобным амфибиям, мелким анапсидам и эозухиям. Позвонки у эозухиям. ящериц процельные, кроме гекконов, у которых они вторично амфицельны. Самые прогрессивные ящерицы – надсемейство Varanoidea – процельные, гекконов, амфицельны. послужили предками змей, а также двух водных семейств, имеющий стрептогнатический внутричелюстной сустав: Aigialosauridae из верхней юры среднего мела имели малоспециализированные конечности и длину не более 1 м. Mosasauridae имели ластообразные гиперфалангические конечности (связь задних с позвоночником утрачена) и низкий хвостовой плавник из отростков верхних и нижних дуг позвонков. Задние туловищные ребра утрачены. Шея короткая для вараноидов (семь позвонков). У некоторых барабанная перепонка обызвествленная, видимо, чтобы выдерживать давление воды при нырянии. Зубы текодонтные – в лунках; у обызвествленная, рода Globidens круглые, для раскусывания аммонитов. Мозазавры достигали в длину 10 м. Беременные самки не обнаружены, а молодь редко встречается со взрослыми, - значит, мозазавры откладывали яйца на суше, что при их размерах и конструкции было непросто. Змеи произошли от вараноидов, отличаясь от них отсутствием грудины и конечности и наличием дополнительных суставных поверхностей вараноидов, позвонков – зигосфена спереди и зигантра сзади. Главный вопрос – были ли предки змей роющими или водными. Общие признаки с роющими ящерицами – удлинение тела, утрата ног, барабанной перепонки и полости среднего уха, сращение век, отсутствие склеральных колец. Но череп роющих ящериц и амфисбен монолитный и прочный, а у змей – ажурный. Даже у ранних змей высоко развит кинетизм черепа, при роющей жизни не выгодный. Утрата змеями барабанной перепонки может объясняться именно крайней подвижность квадратной кости, что делает невозможным ее прикрепление (равно как и сохранение скуловых дуг). Изменение глаза можно связать с ночным образом жизни и ползаньем (в том числе под водой) среди густой растительности. Недавно стали известны два верхнемеловых водных вараноида из Израиля, длинных и безногих: Ophiomorphus и Pachyrachis, годящихся на роль предков змей. Дальнейший успех змей Pachyrachis, был связан с радиацией млекопитающих, служивших им пищей. Известные с третичного периода амфисбены ничего не говорят о происхождении этой группы сверх известного от современных видов. Вероятнее всего, это группа равного змеям ранга, но их конкретные предки из числа ящериц неясны. Пресмыкающиеся часто и с легкостью возвращались к водному образу жизни – животному с низкой интенсивностью метаболизма не сложно задерживать дыхание. Судя по современным морским игуанам, питание и движение в воде не требует у рептилий структурной и физиологической адаптации. Водная локомоция требует у них ¾ энергетических затрат от сухопутной.
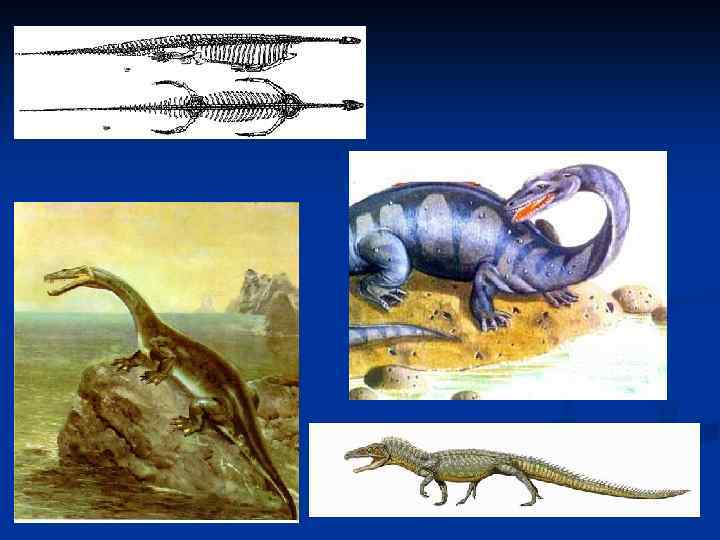
 Инфракласс Sauropterygia характеризуется эвриапсидным черепомм. Самый примитивный его представитель – Claudiosaurus из верхней черепомм. перми Мадагаскара – демонстрирует, что эвриапсидный череп произошел от диапсидного путем утраты нижней скуловой дуги. Хвост у этого животного длинный и не веслообразный, в отличие от увеличенных задних конечностей. Шея длинная, голова маленькая, грудина хрящевая, что возможно, связано с утратой важности чередующихся движений передних конечностей, характерных для наземной локомоции лепидозавров. По строению нёба этот род похож на нотозавров и плезиозавров. лепидозавров. Отряд Nothosauria, известный в основном из среднего триаса Европы и Китая, демонстрирует следующую стадию адаптации к жизни в воде. Nothosauria, Общий план строения конечностей еще не изменен, но форма суставов мало пригодна для передвижения по суше, а сами они содержат много хряща. Нотозавры имели длину от 20 см до 4 м; у всех длинная шея, маленькая голова и средней длины хвост. Большая протяженность плечевого пояса продвинутых нотозавров указывает на гребки передних конечностей вперед-назад. Тазовый пояс меньше и, вероятно, задние конечности использовались в основном для руления. У примитивных форм конечности мелкие, а хвост уплощен, что говорит о уплощен, крокодилообразной локомоции. Представители отряда Plesiosauria крупнее нотоозавров; их передние и задние конечности более сходны между собой, ластообразны и нотоозавров; гиперфалангичны, основная подвижность их – лишь в сочленении с поясами. Огромная передне-задняя протяженность поясов конечностей гиперфалангичны, говорит о мощных гребках вперед-назад, как у морского льва, а не о подводном полете по образцу пингвинов и морских черепах. Хвост короткий и, видимо, служил рулем. Подотряд Plesiosauroidea характеризовался длинной шеей и мелкой головой, а подотряд Pliosauroidea – короткой шеей и большой головой. Плезиозавры известны с триаса по поздний мел. Возможно, они родственны нотозаврам. Напротив, отряд Placodontia – самостоятельная группа. Они известны из прибрежных морских триасовых отложений Европы, Северной Африки и Западной Азии. Предки их неизвестны, и уже ранние представители сильно специализированы. Череп также эвриапсидный, а по эвриапсидный, форме очень похож на среднее между черепами ящерицы и старой лошади. Премаксиллярные зубы торчали вперед и, видимо, служили для откусывания от субстрата двустворчатых моллюсков, а максиллярные и нёбные зубы были большими, толстыми и дробили их раковины. Нижняя челюсть с венечным и угловым отростком. Таков череп Placodus, а у Placochelys передних зубов не было (их заменял роговой клюв). Placodus, Хвост длинный, гребной, конечности неспециализированы для плавания. Placochelys и Henodus имели широкое, плоское тело, покрытое многочисленными мелкими дермальными косточками, не сопоставимыми с крупными пластинами у черепах. Своеобразие Henodus заключается также в т ом, что височное окно у него закрыто. ом, Представители отряда Ichthyosauria – самые специализированные из морских рептилий. Тело у них рыбообразное, гипоцеркальный хвостовой плавник, а конечности – плавники, не годные для передвижения по земле. Таз не сочленялся с позвоночником. Челюсти длинные и узкие. Позвонки короткие, амфицельные, невральные дуги отделены от тел хрящом, поперечных отростков нет. Зубы крепились не в амфицельные, ячейках, а в общей борозде. Судя по отпечаткам, имелся кожный спинной плавник. Ихтиозавры известны с нижнего триаса по нижний мел. Происхождение их неясно. Nanchagosaurus из триаса Китая имел предглазничное окно и нижнее височное окно, что позволяет отнести его к архозаврам, но ихтиозавроподобная форма его тела может быть результатом конвергенции. Триасовые ихтиозавры, в отличие от юрских и меловых, имели длинные хвосты с низким плавником, что иллюстрирует переход к тунцевидному типу плавания у продвинутых ихтиозавров от угревидного. У поздних ихтиозавров кости конечностей настолько изменены, что образуют непрерывный ряд табличек, причем кости предплечья и голени схожи с фалангами пальцев, только крупнее. Число пальцев колебалось от трех (узкоплавничные ихтиозавры) до девяти (узкоплавничные (широкоплавничные), число фаланг сильно увеличено. Барабанной перепонки не было, стремя массивное. Некоторые ихтиозавры широкоплавничные), утрачивали зубы. Ихтиозавры были живородящими; их детеныши рождались хвостом вперед, как у китов, о чем свидетельствуют находки скелетов беременных и рожающих самок. Вымирание ихтиозавров было, вероятно, связано с распространением в мелу акул современного типа. Ихтиозавры были слишком рыбоподобны, чтобы конкурировать с настоящими рыбами.
Инфракласс Sauropterygia характеризуется эвриапсидным черепомм. Самый примитивный его представитель – Claudiosaurus из верхней черепомм. перми Мадагаскара – демонстрирует, что эвриапсидный череп произошел от диапсидного путем утраты нижней скуловой дуги. Хвост у этого животного длинный и не веслообразный, в отличие от увеличенных задних конечностей. Шея длинная, голова маленькая, грудина хрящевая, что возможно, связано с утратой важности чередующихся движений передних конечностей, характерных для наземной локомоции лепидозавров. По строению нёба этот род похож на нотозавров и плезиозавров. лепидозавров. Отряд Nothosauria, известный в основном из среднего триаса Европы и Китая, демонстрирует следующую стадию адаптации к жизни в воде. Nothosauria, Общий план строения конечностей еще не изменен, но форма суставов мало пригодна для передвижения по суше, а сами они содержат много хряща. Нотозавры имели длину от 20 см до 4 м; у всех длинная шея, маленькая голова и средней длины хвост. Большая протяженность плечевого пояса продвинутых нотозавров указывает на гребки передних конечностей вперед-назад. Тазовый пояс меньше и, вероятно, задние конечности использовались в основном для руления. У примитивных форм конечности мелкие, а хвост уплощен, что говорит о уплощен, крокодилообразной локомоции. Представители отряда Plesiosauria крупнее нотоозавров; их передние и задние конечности более сходны между собой, ластообразны и нотоозавров; гиперфалангичны, основная подвижность их – лишь в сочленении с поясами. Огромная передне-задняя протяженность поясов конечностей гиперфалангичны, говорит о мощных гребках вперед-назад, как у морского льва, а не о подводном полете по образцу пингвинов и морских черепах. Хвост короткий и, видимо, служил рулем. Подотряд Plesiosauroidea характеризовался длинной шеей и мелкой головой, а подотряд Pliosauroidea – короткой шеей и большой головой. Плезиозавры известны с триаса по поздний мел. Возможно, они родственны нотозаврам. Напротив, отряд Placodontia – самостоятельная группа. Они известны из прибрежных морских триасовых отложений Европы, Северной Африки и Западной Азии. Предки их неизвестны, и уже ранние представители сильно специализированы. Череп также эвриапсидный, а по эвриапсидный, форме очень похож на среднее между черепами ящерицы и старой лошади. Премаксиллярные зубы торчали вперед и, видимо, служили для откусывания от субстрата двустворчатых моллюсков, а максиллярные и нёбные зубы были большими, толстыми и дробили их раковины. Нижняя челюсть с венечным и угловым отростком. Таков череп Placodus, а у Placochelys передних зубов не было (их заменял роговой клюв). Placodus, Хвост длинный, гребной, конечности неспециализированы для плавания. Placochelys и Henodus имели широкое, плоское тело, покрытое многочисленными мелкими дермальными косточками, не сопоставимыми с крупными пластинами у черепах. Своеобразие Henodus заключается также в т ом, что височное окно у него закрыто. ом, Представители отряда Ichthyosauria – самые специализированные из морских рептилий. Тело у них рыбообразное, гипоцеркальный хвостовой плавник, а конечности – плавники, не годные для передвижения по земле. Таз не сочленялся с позвоночником. Челюсти длинные и узкие. Позвонки короткие, амфицельные, невральные дуги отделены от тел хрящом, поперечных отростков нет. Зубы крепились не в амфицельные, ячейках, а в общей борозде. Судя по отпечаткам, имелся кожный спинной плавник. Ихтиозавры известны с нижнего триаса по нижний мел. Происхождение их неясно. Nanchagosaurus из триаса Китая имел предглазничное окно и нижнее височное окно, что позволяет отнести его к архозаврам, но ихтиозавроподобная форма его тела может быть результатом конвергенции. Триасовые ихтиозавры, в отличие от юрских и меловых, имели длинные хвосты с низким плавником, что иллюстрирует переход к тунцевидному типу плавания у продвинутых ихтиозавров от угревидного. У поздних ихтиозавров кости конечностей настолько изменены, что образуют непрерывный ряд табличек, причем кости предплечья и голени схожи с фалангами пальцев, только крупнее. Число пальцев колебалось от трех (узкоплавничные ихтиозавры) до девяти (узкоплавничные (широкоплавничные), число фаланг сильно увеличено. Барабанной перепонки не было, стремя массивное. Некоторые ихтиозавры широкоплавничные), утрачивали зубы. Ихтиозавры были живородящими; их детеныши рождались хвостом вперед, как у китов, о чем свидетельствуют находки скелетов беременных и рожающих самок. Вымирание ихтиозавров было, вероятно, связано с распространением в мелу акул современного типа. Ихтиозавры были слишком рыбоподобны, чтобы конкурировать с настоящими рыбами.

 Архозавроморфам в той или иной степени свойственна тенденция к движению на выпрямленных в парасагиттальной плоскости конечностях. Грудина описанного для лепидозавроморф типа, облегчающая движения передних ног враскорячку, у них не встречается. У всех враскорячку, архозавроморф длинное, тонкое стремя, наводящее на мысль о наличии барабанной перепонки. Зубы расположены в ячейках (текодонтное состояние). Предчелюстная кость продолжается назад за ноздри, отделяя от них верхнечелюстную. Кроме самих архозавров, к архозавроморфам относятся еще три отряда ранних диапсид: Protorosauria, Trilophosauria и Rhynchosauria. диапсид: Protorosauria, Rhynchosauria. Проторозавры известны с верхней перми по нижнюю юру. Даже у самых ранних представителей шея длиной с туловище, что обусловлено смещением назад плечевого пояса и удлинением отдельных шейных позвонков. У Tanystropheus из среднего триаса Европы шея в четыре раза длиннее тела (не считая хвоста); ноги короткие, со слабо окостеневшим скелетом. Скорее всего, это животное жило в воде, так как на суше оно не смогло бы поддерживать равновесие ни с поднятой вверх шеей, ни с вытянутой вперед. У молодых особей зубы были трехбугорчатые, как у примитивных млекопитающих. Sharovipteryx из верхнего триаса Средней Азии имел очень длинные задние ноги, трехбугорчатые, поддерживавшие летательную перепонку. Это единственный случай преимущественного использования позвоночными задних конечностей для планирования. Трилофозавры существовали с нижнего по верхний триас. Передние концы челюстей беззубые (возможно, они были покрыты роговым клювом), а щечные зубы вытянуты в поперечном направлении и образуют острые режущие поверхности, приспособленные для перетирания растительной пищи. Нижнее височное окно закрыто ради укрепления черепа в соответствии с мощными зубами. Ринхозавры встречались также на протяжение всего триаса. У продвинутых ринхозавров короткий и очень широкий череп с непарной срединной ноздрей и безубыми концами челюстей, несшими роговой клюв. Верхнечелюстная и задняя часть зубной кости несут множество рядов мелких перетирающих зубов. Эти зубы не сменялись регулярно, а добавлялись спереди назад по мере роста челюсти. Верхние и нижние стирались друг о друга, а когда они снашивались до основания, костные поверхности смыкались. Аддукторная камера очень объемистая. Похоже, ринхозавры питались высокоабразивной растительной пищей. Размеры и пропорции тела ринхозавров свиноподобны. Задние конечности полностью улучшенного типа (двигающиеся в вертикальной плоскости), а заплюсна образует одноосный сустав для движения стопы вперед – назад. Передние ноги остались более раздвинутыми. Ринхозавры были обычными и многочисленными стадными животными. Собственно архозавры характеризуются наличием особого отверстия спереди от глазницы – предглазничного окна. Исходной группой архозавров был отряд Thecodontia. Это название дано им не совсем справедливо, так как текодонтные зубы присущи отнюдь не Thecodontia. только текодонтам. Известно несколько семейств этого отряда. Proterosuchidae – примитивнейшие из текодонтов. Известны они с верхней перми по нижний триас. Пиниальное отверствие сохранилось, мандибулярное окно отсутствует. Предчелюстные кости загибались вниз характерным крючком, способствующим удержанию жертвы. Запястье и заплюсна окостеневали слабо, но основная линия сгибания в заплюсне проходила между костями проксимального (таранной и пяточной) и дистального рядов; пальцев на ногах было пять. Ноги занимали, в отличие от всех остальных архозавров, примитивное растопыренное положение (неулучшенное). Головка бедренной кости расположена непосредственно на ее конце, а не отклонена вбок. В соответствии с горизонтальным расположением бедра лобковая и седалищная кости, на которых расположены двигающие бедро мышцы, расположены почти горизонтально. Размеры тела были крупные, до 4 м. По-видимому, протерозухиды вели полуводный образ жизни. Erythrosuchidae из нижнего триаса были похожи на протерозухид: тоже с плохо окостеневшими запястьем и заплюсной, протерозухид: мезотарсальным суставом и крючковидной верхней челюстью. Отдельные их представители были еще крупнее (череп достигал в длину 1 м). Головка бедренной кости еще не повернута внутрь, но вертлужная впадина ориентирована более вентрально, чем у протерозухид, а таз начал вентрально, протерозухид, превращаться в трехлучевой, что указывает на более вертикальное положение задних ног. трехлучевой, Euparkeriidae из нижнего триаса – мелкие (меньше 1 м), изящные животные с задними ногами в полтора раза длиннее передних, что, возможно, свидетельствует о факультативной бипедальности (не облигатной, так как суставные поверхности и конфигурация бедренной кости и заплюсны неспециализированы для двуногого передвижения). Заплюсна окостеневает не сильно (видимо, из-за мелких размеров тела), голеностопный сустав мезотарсальный. Когда линия сгибания проходит между проксимальным и дистальным рядами костей мезотарсальный. заплюсны, это обеспечивает симметричную передачу усилия к стопе, что важно при положении задних ног, близкому к вертикальному (но еще полуулучшенному).
Архозавроморфам в той или иной степени свойственна тенденция к движению на выпрямленных в парасагиттальной плоскости конечностях. Грудина описанного для лепидозавроморф типа, облегчающая движения передних ног враскорячку, у них не встречается. У всех враскорячку, архозавроморф длинное, тонкое стремя, наводящее на мысль о наличии барабанной перепонки. Зубы расположены в ячейках (текодонтное состояние). Предчелюстная кость продолжается назад за ноздри, отделяя от них верхнечелюстную. Кроме самих архозавров, к архозавроморфам относятся еще три отряда ранних диапсид: Protorosauria, Trilophosauria и Rhynchosauria. диапсид: Protorosauria, Rhynchosauria. Проторозавры известны с верхней перми по нижнюю юру. Даже у самых ранних представителей шея длиной с туловище, что обусловлено смещением назад плечевого пояса и удлинением отдельных шейных позвонков. У Tanystropheus из среднего триаса Европы шея в четыре раза длиннее тела (не считая хвоста); ноги короткие, со слабо окостеневшим скелетом. Скорее всего, это животное жило в воде, так как на суше оно не смогло бы поддерживать равновесие ни с поднятой вверх шеей, ни с вытянутой вперед. У молодых особей зубы были трехбугорчатые, как у примитивных млекопитающих. Sharovipteryx из верхнего триаса Средней Азии имел очень длинные задние ноги, трехбугорчатые, поддерживавшие летательную перепонку. Это единственный случай преимущественного использования позвоночными задних конечностей для планирования. Трилофозавры существовали с нижнего по верхний триас. Передние концы челюстей беззубые (возможно, они были покрыты роговым клювом), а щечные зубы вытянуты в поперечном направлении и образуют острые режущие поверхности, приспособленные для перетирания растительной пищи. Нижнее височное окно закрыто ради укрепления черепа в соответствии с мощными зубами. Ринхозавры встречались также на протяжение всего триаса. У продвинутых ринхозавров короткий и очень широкий череп с непарной срединной ноздрей и безубыми концами челюстей, несшими роговой клюв. Верхнечелюстная и задняя часть зубной кости несут множество рядов мелких перетирающих зубов. Эти зубы не сменялись регулярно, а добавлялись спереди назад по мере роста челюсти. Верхние и нижние стирались друг о друга, а когда они снашивались до основания, костные поверхности смыкались. Аддукторная камера очень объемистая. Похоже, ринхозавры питались высокоабразивной растительной пищей. Размеры и пропорции тела ринхозавров свиноподобны. Задние конечности полностью улучшенного типа (двигающиеся в вертикальной плоскости), а заплюсна образует одноосный сустав для движения стопы вперед – назад. Передние ноги остались более раздвинутыми. Ринхозавры были обычными и многочисленными стадными животными. Собственно архозавры характеризуются наличием особого отверстия спереди от глазницы – предглазничного окна. Исходной группой архозавров был отряд Thecodontia. Это название дано им не совсем справедливо, так как текодонтные зубы присущи отнюдь не Thecodontia. только текодонтам. Известно несколько семейств этого отряда. Proterosuchidae – примитивнейшие из текодонтов. Известны они с верхней перми по нижний триас. Пиниальное отверствие сохранилось, мандибулярное окно отсутствует. Предчелюстные кости загибались вниз характерным крючком, способствующим удержанию жертвы. Запястье и заплюсна окостеневали слабо, но основная линия сгибания в заплюсне проходила между костями проксимального (таранной и пяточной) и дистального рядов; пальцев на ногах было пять. Ноги занимали, в отличие от всех остальных архозавров, примитивное растопыренное положение (неулучшенное). Головка бедренной кости расположена непосредственно на ее конце, а не отклонена вбок. В соответствии с горизонтальным расположением бедра лобковая и седалищная кости, на которых расположены двигающие бедро мышцы, расположены почти горизонтально. Размеры тела были крупные, до 4 м. По-видимому, протерозухиды вели полуводный образ жизни. Erythrosuchidae из нижнего триаса были похожи на протерозухид: тоже с плохо окостеневшими запястьем и заплюсной, протерозухид: мезотарсальным суставом и крючковидной верхней челюстью. Отдельные их представители были еще крупнее (череп достигал в длину 1 м). Головка бедренной кости еще не повернута внутрь, но вертлужная впадина ориентирована более вентрально, чем у протерозухид, а таз начал вентрально, протерозухид, превращаться в трехлучевой, что указывает на более вертикальное положение задних ног. трехлучевой, Euparkeriidae из нижнего триаса – мелкие (меньше 1 м), изящные животные с задними ногами в полтора раза длиннее передних, что, возможно, свидетельствует о факультативной бипедальности (не облигатной, так как суставные поверхности и конфигурация бедренной кости и заплюсны неспециализированы для двуногого передвижения). Заплюсна окостеневает не сильно (видимо, из-за мелких размеров тела), голеностопный сустав мезотарсальный. Когда линия сгибания проходит между проксимальным и дистальным рядами костей мезотарсальный. заплюсны, это обеспечивает симметричную передачу усилия к стопе, что важно при положении задних ног, близкому к вертикальному (но еще полуулучшенному).

 Phitosauria встречаются в верхнем триасе. По форме тела и образу жизни они напоминали современных крокодилов, но ноздри находились не на конце их длинной узкой морды, а сверху перед глазами. Конфигурация небных костей позади хоан указывает на наличие мягкого вторичного неба. Строение скелета конечностей и их поясов гораздо примитивнее, чем у крокодилов. Лобковая и седалищная кости пластиновидные; пятая плюсневая кость крючковидная, как у лепидозавров, а бугор пяточной кости направлен наружу, а не назад. Но, как у пластиновидные; лепидозавров, крокодилов, таранная кость прочно прикреплена к берцовым костям, а пяточная – к дистальному ряду заплюсны, и линия сгибания голеностопного сустава проходит между таранной и пяточной костями. Положение конечностей полуулучшенное, но ближе к полуулучшенное, раскоряченному. Aetosauria из верхнего триаса – единственные растительноядные текодонты. Голова у них была маленькая, зубы листовидные. Таз продвинутый, трехлучевой. Положение конечностей полуулучшенное. Линия сгибания голеностопного сустава – между таранной и трехлучевой. полуулучшенное. пяточной костью, пяточная несет крупный бугор для крепления сухожилия икроножной мышцы. Тело было покрыто мощным панцирем, размеры достигали 3 метров. Rauisuchidae – крупные (до 6 м) хищники среднего и верхнего триаса. Положение конечностей рауизухид было полностью улучшенным, но весьма оригинальным способом. Бедренная кость сохраняет примитивное строение (головка на конце), но вертлужная впадина развернута вниз. Таким образом, вес тела поддерживался непосредственно вертикальной осью конечности. Таз трехлучевой, у трехлучевой, поздних представителей появилась третья пара крестцовых ребер. Линия сгибания голеностопного сустава проходила между таранной и пяточной костью. Бугор пяточной кости направлен назад и служил для крепления икроножной мышцы. Пятая плюсневая кость (и пятый палец) редуцирована, утратив роль рычага. Симметричное (в пределах одной ноги) приложение силы икроножной мышцы важно при вертикальной постановке ноги. Передние и задние ноги рауизухид были примерно одной длины, что указывает на четвероногое передвижение. Ornithosuchidae из верхнего триаса – крупные (до 4 м) стройные животные с задними ногами на треть длиннее передних. Череп похож на череп хищных динозавров: сжатый с боков, с крупными лезвиеподобными зубами. Задние ноги имеют полуулучшенное положение благодаря частичному повороту внутрь головки бедренной кости, заплюсна сходна с таковой рауизухид, крестцовых ребер три рауизухид, пары. Lagosuchidae из среднего триаса, возможно, были предками динозавров. Это мелкие (менее ½ м), очень изящные, длинноногие животные, у которых задние ноги значительно длиннее передних, а голень гораздо длиннее бедра. В заплюсне имеется одноосный сустав между крепившимися к берцовым костям таранной и пяточной костями с проксимального конца и дистальным рядом заплюсны, крепившимся к плюсневым костям – с дистального. Пятая плюсневая кость не крючковидная. Отряд Crocodylia характеризуется массивным, укрепленным черепом в противоположность его ажурной структуре у большинства текодонтов. Предглазничное окно никогда не бывает крупным, а у современных форм закрыто. В той или иной степени развито вторичное нёбо. Его появление обычно связывают с разделением носовой и ротовой полости, что важно для дышащих воздухом водных животных, не имеющих губ. Однако вторичное нёбо начало формироваться еще у ранних наземных крокодилов, и его первоначальной функцией могло быть укрепление удлиненной и уплощенной морды. Механика питания крокодилов в корне отлична от присущей другим архозаврам, что, возможно, объясняет выживание крокодилов вместе с динозаврами на протяжение всего мезозоя, когда более динозавроподобные текодонты уже вымерли. Все мышцы, воздействующие на нижнюю челюсть, прикрепляются около ее заднего конца. Это позволяет длинным челюстям широко раскрываться и быстро захлопываться. Главная мышца, закрывающая пасть – крыловидная. Ее волокна направлены у крокодилов почти горизонтально. Она продолжается далеко вперед над нёбом, и ее длинные волокна, сокращаясь, развивают мощное усилие в широком диапазоне углов раскрытия челюстей. Результирующая сила всей челюстной мускулатуры направлена вперед и вверх, как и отклоненная в этом же направлении мощно развитая квадратная кость. Другой отличительный признак крокодилов – пневматизация почти всех костей черепа. В нем имеются две главных системы воздухоносных каналов: одна развилась из евстахиевых труб и пневматизирует затылочную, переднюю ушную, квадратную, клиновидную и сочленовную кости, а вторая связана с носовыми проходами и пневматизирует предчелюстную, верхнечелюстную, нёюную и крыловидную предчелюстную, кости. Причина сильной пневматизации черепа крокодилов точно не известна, но это может быть связано с усилением остроты слуха, по которой современные крокодилы превосходят остальных современных пресмыкающихся и не уступают птицам и млекопитающим.
Phitosauria встречаются в верхнем триасе. По форме тела и образу жизни они напоминали современных крокодилов, но ноздри находились не на конце их длинной узкой морды, а сверху перед глазами. Конфигурация небных костей позади хоан указывает на наличие мягкого вторичного неба. Строение скелета конечностей и их поясов гораздо примитивнее, чем у крокодилов. Лобковая и седалищная кости пластиновидные; пятая плюсневая кость крючковидная, как у лепидозавров, а бугор пяточной кости направлен наружу, а не назад. Но, как у пластиновидные; лепидозавров, крокодилов, таранная кость прочно прикреплена к берцовым костям, а пяточная – к дистальному ряду заплюсны, и линия сгибания голеностопного сустава проходит между таранной и пяточной костями. Положение конечностей полуулучшенное, но ближе к полуулучшенное, раскоряченному. Aetosauria из верхнего триаса – единственные растительноядные текодонты. Голова у них была маленькая, зубы листовидные. Таз продвинутый, трехлучевой. Положение конечностей полуулучшенное. Линия сгибания голеностопного сустава – между таранной и трехлучевой. полуулучшенное. пяточной костью, пяточная несет крупный бугор для крепления сухожилия икроножной мышцы. Тело было покрыто мощным панцирем, размеры достигали 3 метров. Rauisuchidae – крупные (до 6 м) хищники среднего и верхнего триаса. Положение конечностей рауизухид было полностью улучшенным, но весьма оригинальным способом. Бедренная кость сохраняет примитивное строение (головка на конце), но вертлужная впадина развернута вниз. Таким образом, вес тела поддерживался непосредственно вертикальной осью конечности. Таз трехлучевой, у трехлучевой, поздних представителей появилась третья пара крестцовых ребер. Линия сгибания голеностопного сустава проходила между таранной и пяточной костью. Бугор пяточной кости направлен назад и служил для крепления икроножной мышцы. Пятая плюсневая кость (и пятый палец) редуцирована, утратив роль рычага. Симметричное (в пределах одной ноги) приложение силы икроножной мышцы важно при вертикальной постановке ноги. Передние и задние ноги рауизухид были примерно одной длины, что указывает на четвероногое передвижение. Ornithosuchidae из верхнего триаса – крупные (до 4 м) стройные животные с задними ногами на треть длиннее передних. Череп похож на череп хищных динозавров: сжатый с боков, с крупными лезвиеподобными зубами. Задние ноги имеют полуулучшенное положение благодаря частичному повороту внутрь головки бедренной кости, заплюсна сходна с таковой рауизухид, крестцовых ребер три рауизухид, пары. Lagosuchidae из среднего триаса, возможно, были предками динозавров. Это мелкие (менее ½ м), очень изящные, длинноногие животные, у которых задние ноги значительно длиннее передних, а голень гораздо длиннее бедра. В заплюсне имеется одноосный сустав между крепившимися к берцовым костям таранной и пяточной костями с проксимального конца и дистальным рядом заплюсны, крепившимся к плюсневым костям – с дистального. Пятая плюсневая кость не крючковидная. Отряд Crocodylia характеризуется массивным, укрепленным черепом в противоположность его ажурной структуре у большинства текодонтов. Предглазничное окно никогда не бывает крупным, а у современных форм закрыто. В той или иной степени развито вторичное нёбо. Его появление обычно связывают с разделением носовой и ротовой полости, что важно для дышащих воздухом водных животных, не имеющих губ. Однако вторичное нёбо начало формироваться еще у ранних наземных крокодилов, и его первоначальной функцией могло быть укрепление удлиненной и уплощенной морды. Механика питания крокодилов в корне отлична от присущей другим архозаврам, что, возможно, объясняет выживание крокодилов вместе с динозаврами на протяжение всего мезозоя, когда более динозавроподобные текодонты уже вымерли. Все мышцы, воздействующие на нижнюю челюсть, прикрепляются около ее заднего конца. Это позволяет длинным челюстям широко раскрываться и быстро захлопываться. Главная мышца, закрывающая пасть – крыловидная. Ее волокна направлены у крокодилов почти горизонтально. Она продолжается далеко вперед над нёбом, и ее длинные волокна, сокращаясь, развивают мощное усилие в широком диапазоне углов раскрытия челюстей. Результирующая сила всей челюстной мускулатуры направлена вперед и вверх, как и отклоненная в этом же направлении мощно развитая квадратная кость. Другой отличительный признак крокодилов – пневматизация почти всех костей черепа. В нем имеются две главных системы воздухоносных каналов: одна развилась из евстахиевых труб и пневматизирует затылочную, переднюю ушную, квадратную, клиновидную и сочленовную кости, а вторая связана с носовыми проходами и пневматизирует предчелюстную, верхнечелюстную, нёюную и крыловидную предчелюстную, кости. Причина сильной пневматизации черепа крокодилов точно не известна, но это может быть связано с усилением остроты слуха, по которой современные крокодилы превосходят остальных современных пресмыкающихся и не уступают птицам и млекопитающим.

 Крокодилы, в отличие от остальных архозавров, пошли по пути глубокого усовершенствования четвероногой локомоции. Ключицы утрачены, как у бегающих млекопитающих, что позволяет гленоидной впадине смещаться вперед и назад ради увеличения длины шага. Крокодилы – единственные из современных рептилий, способные бегать галопом, а способность к этому у ранних наземных родов должна была быть выражена тем более сильно. Для таза характерно неучастие лобковых костей в формировании вертлужной впадины у всех родов с начала юры. Головка бедренной кости умеренно повернута внутрь. Берцовые кости опираются на таранную, образующую сустав с пяточной костью, несущей рычаг для крепления икроножной мышцы. Пятый палец стопы редуцирован, пятая плюсневая кость маленькая и не крючковидная. Крокодилы подразделяются на 4 подотряда. Sphenosuchia существовали со среднего по верхний триас. Они еще обладали крупным предглазничным окном, вторичное нёбо очень короткое и образовано лишь верхнечелюстными костями. Пропорции тела этих мелких, до полуметра в длину, животных очень изящные и указывают на наземный образ жизни; ноги очень длинные, приспособленные для бега. Protosuchia существовали с верхнего триаса по среднюю юру. Верхнее нёбо образовано верхнечелюстными и нёбными костями. Размеры более крупные (около 1 м), а сложение – тяжелее, чему сфенозухий, но пропорции тела все равно характерны для наземных сфенозухий, бегающих животных. Mesosuchia из поздней юры – миоцена имели вторичное нёбо, образованное верхнечелюстными, нёбными и крыловидными костями. Существовали как наземные, так и водные мезозухии. Морские позднеюрские Teleosauridae имели длинную, узкую мезозухии. гавиалоподобную морду и передние ноги в два раза короче задних. Их хвост и ноги имели ту же степень специализации к передвижению в водной среде, как и у современных крокодилов. Морские Metriorhynchidae, встречавшиеся с поздней юры до позднего мела, имели Metriorhynchidae, ластообразные конечности и загнутый вниз конец хвоста, поддерживавший гипоцеркальный хвостовой плавник. Характерный для крокодилов дермальный панцирь утрачен, шея на 1 или 2 позвонка короче, чем обычно (9). У представителей этих двух семейств верхние височные окна гораздо крупнее, чем у полуводных и наземных крокодилов. Их увеличение, возможно, связано с длинными челюстными мышцами и очень широким раскрыванием пасти. Sebecidae, жившие с палеоцена по миоцен, имели крупные (более 10 м) размеры и высокие, Sebecidae, сжатые с боков, черепа. Зубы представляли собой такие же сжатые с боков пильчатые лезвия, как у хищных динозавров (такая форма называется зифодонтной). Себециды вели наземный образ жизни. зифодонтной). Eusuchia отличаются тем, что вторичное нёбо у них образовано верхнечелюстными нёбными и крыловидными костями, а также тем, что позвонки у них процельные, а не амфицельные, как у остальных крокодилов. К эвзухиям относятся крокодилы современного процельные, амфицельные, типа, появившиеся в верхнем мелу. В раннем третичном периоде эвзухии были гораздо многочисленнее, разнообразнее и распространеннее, чем сейчас. Их упадок, по-видимому, последовал за похолоданием климата на Земле.
Крокодилы, в отличие от остальных архозавров, пошли по пути глубокого усовершенствования четвероногой локомоции. Ключицы утрачены, как у бегающих млекопитающих, что позволяет гленоидной впадине смещаться вперед и назад ради увеличения длины шага. Крокодилы – единственные из современных рептилий, способные бегать галопом, а способность к этому у ранних наземных родов должна была быть выражена тем более сильно. Для таза характерно неучастие лобковых костей в формировании вертлужной впадины у всех родов с начала юры. Головка бедренной кости умеренно повернута внутрь. Берцовые кости опираются на таранную, образующую сустав с пяточной костью, несущей рычаг для крепления икроножной мышцы. Пятый палец стопы редуцирован, пятая плюсневая кость маленькая и не крючковидная. Крокодилы подразделяются на 4 подотряда. Sphenosuchia существовали со среднего по верхний триас. Они еще обладали крупным предглазничным окном, вторичное нёбо очень короткое и образовано лишь верхнечелюстными костями. Пропорции тела этих мелких, до полуметра в длину, животных очень изящные и указывают на наземный образ жизни; ноги очень длинные, приспособленные для бега. Protosuchia существовали с верхнего триаса по среднюю юру. Верхнее нёбо образовано верхнечелюстными и нёбными костями. Размеры более крупные (около 1 м), а сложение – тяжелее, чему сфенозухий, но пропорции тела все равно характерны для наземных сфенозухий, бегающих животных. Mesosuchia из поздней юры – миоцена имели вторичное нёбо, образованное верхнечелюстными, нёбными и крыловидными костями. Существовали как наземные, так и водные мезозухии. Морские позднеюрские Teleosauridae имели длинную, узкую мезозухии. гавиалоподобную морду и передние ноги в два раза короче задних. Их хвост и ноги имели ту же степень специализации к передвижению в водной среде, как и у современных крокодилов. Морские Metriorhynchidae, встречавшиеся с поздней юры до позднего мела, имели Metriorhynchidae, ластообразные конечности и загнутый вниз конец хвоста, поддерживавший гипоцеркальный хвостовой плавник. Характерный для крокодилов дермальный панцирь утрачен, шея на 1 или 2 позвонка короче, чем обычно (9). У представителей этих двух семейств верхние височные окна гораздо крупнее, чем у полуводных и наземных крокодилов. Их увеличение, возможно, связано с длинными челюстными мышцами и очень широким раскрыванием пасти. Sebecidae, жившие с палеоцена по миоцен, имели крупные (более 10 м) размеры и высокие, Sebecidae, сжатые с боков, черепа. Зубы представляли собой такие же сжатые с боков пильчатые лезвия, как у хищных динозавров (такая форма называется зифодонтной). Себециды вели наземный образ жизни. зифодонтной). Eusuchia отличаются тем, что вторичное нёбо у них образовано верхнечелюстными нёбными и крыловидными костями, а также тем, что позвонки у них процельные, а не амфицельные, как у остальных крокодилов. К эвзухиям относятся крокодилы современного процельные, амфицельные, типа, появившиеся в верхнем мелу. В раннем третичном периоде эвзухии были гораздо многочисленнее, разнообразнее и распространеннее, чем сейчас. Их упадок, по-видимому, последовал за похолоданием климата на Земле.