

лекция 3.ppt
- Количество слайдов: 59
 Лекция № 3 Тема: Гипотезы происхождения многоклеточных животных Царство Metazoa подцарство Prometazoa
Лекция № 3 Тема: Гипотезы происхождения многоклеточных животных Царство Metazoa подцарство Prometazoa
 и вариации (Б) у одноклеточных Sarcodina Mastigophora Ciliophora А Mastigophora Ciliophora") Структурный уровень (А) и вариации (Б) у одноклеточных Sarcodina Mastigophora Ciliophora А Mastigophora Ciliophora Sarcodina Б Группа Mastigophora имеет средний структурный уровень, что при широких вариационных возможностях обеспечивает переход к более высоким уровням организации.
Структурный уровень (А) и вариации (Б) у одноклеточных Sarcodina Mastigophora Ciliophora А Mastigophora Ciliophora Sarcodina Б Группа Mastigophora имеет средний структурный уровень, что при широких вариационных возможностях обеспечивает переход к более высоким уровням организации.
 На уровень многоклеточности выходят только те одноклеточные, которые имеют оогамию и гаметическую редукцию. Поскольку: 1) Развивающуюся зиготу необходимо обеспечить питательными веществами (оогамия), а гаметическая редукция обеспечивает максимальное число генетических вариант. 2) У многоклеточных животных в жизненном цикле нет изогамии, анизогамии с зиготической или промежуточной редукцией.
На уровень многоклеточности выходят только те одноклеточные, которые имеют оогамию и гаметическую редукцию. Поскольку: 1) Развивающуюся зиготу необходимо обеспечить питательными веществами (оогамия), а гаметическая редукция обеспечивает максимальное число генетических вариант. 2) У многоклеточных животных в жизненном цикле нет изогамии, анизогамии с зиготической или промежуточной редукцией.
 Клональные колонии – многоклеточный") Возможные пути образования многоклеточного животного организма из одноклеточных эукариот: 1) Клональные колонии – многоклеточный организм образуется из потомков одной клетки, которые не расходятся после деления, а соединяются внеклеточным матриксом. . .
Возможные пути образования многоклеточного животного организма из одноклеточных эукариот: 1) Клональные колонии – многоклеточный организм образуется из потомков одной клетки, которые не расходятся после деления, а соединяются внеклеточным матриксом. . .
 Volvox
Volvox
 Агрегированные колонии: исходно отдельные клетки собираются вместе и образуют многоклеточный агрегат (шаровидная колония") 2) Агрегированные колонии: исходно отдельные клетки собираются вместе и образуют многоклеточный агрегат (шаровидная колония простейших). . .
2) Агрегированные колонии: исходно отдельные клетки собираются вместе и образуют многоклеточный агрегат (шаровидная колония простейших). . .
 Гипотеза целлюляризации: разделение исходного многоядерного одноклеточного организма на отдельные клетки. Недостаток: ни у") 3) Гипотеза целлюляризации: разделение исходного многоядерного одноклеточного организма на отдельные клетки. Недостаток: ни у кого из ныне живущих форм целлюляризация не описана; этот путь не подтверждается современными ультраструктурными данными.
3) Гипотеза целлюляризации: разделение исходного многоядерного одноклеточного организма на отдельные клетки. Недостаток: ни у кого из ныне живущих форм целлюляризация не описана; этот путь не подтверждается современными ультраструктурными данными.
: Инвагинация (впячивание) бластулообразного (шаровидного) организма (1); возникновение первичной") Гипотеза гастреи (Геккель, 1874 г. ): Инвагинация (впячивание) бластулообразного (шаровидного) организма (1); возникновение первичной кишечной полости (гастреи) с ротовой порой (2).
Гипотеза гастреи (Геккель, 1874 г. ): Инвагинация (впячивание) бластулообразного (шаровидного) организма (1); возникновение первичной кишечной полости (гастреи) с ротовой порой (2).
 инвагинационное образование двуслойности встречается только у Deuterostomia вторичноротых (иглокожие,") Недостатки гипотезы гастреи : а) инвагинационное образование двуслойности встречается только у Deuterostomia вторичноротых (иглокожие, хордовые); б) гастрея в качестве типичной личиночной стадии у ныне живущих не отмечена; гастрее соответствуют все стрекающие кишечнополостные животные; в) инвагинация возможна если бластула; заполнена первично полостной жидкостью; г) предковая форма полой бластулы; (целобластулы) не могла существовать миллионы лет поскольку у бластулы не было механизмов обеспечения питания;
Недостатки гипотезы гастреи : а) инвагинационное образование двуслойности встречается только у Deuterostomia вторичноротых (иглокожие, хордовые); б) гастрея в качестве типичной личиночной стадии у ныне живущих не отмечена; гастрее соответствуют все стрекающие кишечнополостные животные; в) инвагинация возможна если бластула; заполнена первично полостной жидкостью; г) предковая форма полой бластулы; (целобластулы) не могла существовать миллионы лет поскольку у бластулы не было механизмов обеспечения питания;
 Фагоцителла (паренхимелла) – моноаксонное (одна ось),") Гипотеза фагоцителлы (И. И. Мечников 1886 г. ) Фагоцителла (паренхимелла) – моноаксонное (одна ось), гетерополярное (различно направленные полюса), радиально симметричное (с бесконечно большим порядком радиальной симметрии) гипотетическое животное – исходная форма многоклеточных.
Гипотеза фагоцителлы (И. И. Мечников 1886 г. ) Фагоцителла (паренхимелла) – моноаксонное (одна ось), гетерополярное (различно направленные полюса), радиально симметричное (с бесконечно большим порядком радиальной симметрии) гипотетическое животное – исходная форма многоклеточных.
 из слоя поверхностных клеток –") Фагоцителла состоит (подобно личинке современных низших многоклеточных – паренхимуле) из слоя поверхностных клеток – эктодермы, или кинобласта (функции отграничения, внешнего обмена и движения), и внутренней клеточной массы – паренхимы, или фагоцитобласта (функция внутреннего обмена, внутриклеточного пищеварения (фагоцитоз)). Из кинобласта и фагоцитобласта в ходе эволюции возникло всё многообразие форм тканей многоклеточных животных организмов.
Фагоцителла состоит (подобно личинке современных низших многоклеточных – паренхимуле) из слоя поверхностных клеток – эктодермы, или кинобласта (функции отграничения, внешнего обмена и движения), и внутренней клеточной массы – паренхимы, или фагоцитобласта (функция внутреннего обмена, внутриклеточного пищеварения (фагоцитоз)). Из кинобласта и фагоцитобласта в ходе эволюции возникло всё многообразие форм тканей многоклеточных животных организмов.
 когда фагоцитирующие клетки уходят во внутрь мультиполярно") Способы перемещения фагоцитирующих клеток в фагоцителле: а) когда фагоцитирующие клетки уходят во внутрь мультиполярно образуя паренхимулу; Сейчас на этом уровне развития – паренхимула губок, планула книдарий (стрекающих). Аборальный полюс Оральный полюс
Способы перемещения фагоцитирующих клеток в фагоцителле: а) когда фагоцитирующие клетки уходят во внутрь мультиполярно образуя паренхимулу; Сейчас на этом уровне развития – паренхимула губок, планула книдарий (стрекающих). Аборальный полюс Оральный полюс
 униполярная иммиграция – фагоцитирующие клетки") Способы перемещения фагоцитирующих клеток в фагоцителле: Аборальный полюс б) униполярная иммиграция – фагоцитирующие клетки более крупные, расположены на оральном полюсе и переходят (иммигрируют) во внутрь не согласованно (по отдельности), образуя паренхимулу Оральный полюс
Способы перемещения фагоцитирующих клеток в фагоцителле: Аборальный полюс б) униполярная иммиграция – фагоцитирующие клетки более крупные, расположены на оральном полюсе и переходят (иммигрируют) во внутрь не согласованно (по отдельности), образуя паренхимулу Оральный полюс
 При униполярном типе развития идёт дифференцировка фагоцитобласта на центральный фагоцитобласт энтодермы и периферический фагоцитобласт (третий зародышевый листок - мезодерма). Телобластический тип закладки мезодермы – третьего зародышевого листка и вторичной полости тела целома
При униполярном типе развития идёт дифференцировка фагоцитобласта на центральный фагоцитобласт энтодермы и периферический фагоцитобласт (третий зародышевый листок - мезодерма). Телобластический тип закладки мезодермы – третьего зародышевого листка и вторичной полости тела целома
 возникновение инвагинации – развитие из униполярного типа – бластула крупная, полая. Возникает протоцель") в) возникновение инвагинации – развитие из униполярного типа – бластула крупная, полая. Возникает протоцель – полостная жидкость в которой накапливаются метаболиты от покровных клеток. Метаболиты являются сигнальными молекулами которые обеспечивают единовременное перемещение всех фагоцитирующих клеток орального Оральный полюса – происходит инвагинация. полюс Контроль метаболитами перемещения фагоцитов Аборальный полюс
в) возникновение инвагинации – развитие из униполярного типа – бластула крупная, полая. Возникает протоцель – полостная жидкость в которой накапливаются метаболиты от покровных клеток. Метаболиты являются сигнальными молекулами которые обеспечивают единовременное перемещение всех фагоцитирующих клеток орального Оральный полюса – происходит инвагинация. полюс Контроль метаболитами перемещения фагоцитов Аборальный полюс
 При инвагинационном типе развития происходит выпячивание карманов, затем идёт «отшнуровка» этих карманов. Эпителий карманов – это мезодерма, а полость – это целом (вторичная полость тела) Энтероцельный тип развития, при этом идёт переход к билатеральной симметрии (двусторонняя – симметрия зеркального отражения) и происходит образование целома.
При инвагинационном типе развития происходит выпячивание карманов, затем идёт «отшнуровка» этих карманов. Эпителий карманов – это мезодерма, а полость – это целом (вторичная полость тела) Энтероцельный тип развития, при этом идёт переход к билатеральной симметрии (двусторонняя – симметрия зеркального отражения) и происходит образование целома.
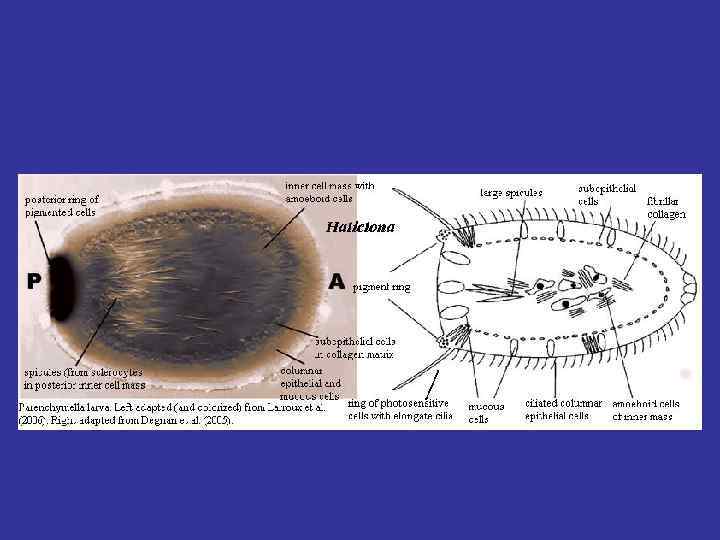
, из") Типы фагоцителл: Фагоцителла 1 – имеет один эпителий кинобласт. Соответствует паренхимуле (у губок), из ныне живущих - трихоплакс. Паренхимула Mycale lorenzi (Cornacuspongida).
Типы фагоцителл: Фагоцителла 1 – имеет один эпителий кинобласт. Соответствует паренхимуле (у губок), из ныне живущих - трихоплакс. Паренхимула Mycale lorenzi (Cornacuspongida).
 Типы фагоцителл Фагоцителла 2 – имеет два эпителия: кинобласт и фагоцитобласт. Изначально радиально симметричная, но её производные имеют тенденцию к билатеральной симметрии. Ей соответствуют: амфибластула высших губок (у которых есть тенденция к инвагинации), аскон губок, и все стрекающие кишечнополостные Амфибластула Sycon raphanus (Calcarea, Heterocoela) (по Ф. Э. Шульце) Аскон губок
Типы фагоцителл Фагоцителла 2 – имеет два эпителия: кинобласт и фагоцитобласт. Изначально радиально симметричная, но её производные имеют тенденцию к билатеральной симметрии. Ей соответствуют: амфибластула высших губок (у которых есть тенденция к инвагинации), аскон губок, и все стрекающие кишечнополостные Амфибластула Sycon raphanus (Calcarea, Heterocoela) (по Ф. Э. Шульце) Аскон губок
. Ей") Типы фагоцителл Фагоцителла 3 – три эпителия и периферический фагоцитобласт (личинка диплеврула иглокожих). Ей соответствует сикон и лейкон губок. А Б типы строения губок и их канальной системы (по Гессе) А - сикон; Б - лейкон.
Типы фагоцителл Фагоцителла 3 – три эпителия и периферический фагоцитобласт (личинка диплеврула иглокожих). Ей соответствует сикон и лейкон губок. А Б типы строения губок и их канальной системы (по Гессе) А - сикон; Б - лейкон.
: 1 - зачаток") Две стадии развития личинки морской звезды; энтероцельное образование целома (по Мечникову): 1 - зачаток каменистого канала, 2 правый целомический мешок, 3 - кишечник личинки, 4 - клетки мезенхимы
Две стадии развития личинки морской звезды; энтероцельное образование целома (по Мечникову): 1 - зачаток каменистого канала, 2 правый целомический мешок, 3 - кишечник личинки, 4 - клетки мезенхимы
, на аборальный полюс (Б), либо параллельно") Каждая фагоцителла может лечь на оральный полюс (А), на аборальный полюс (Б), либо параллельно субстрату (В). А Б В субстрат А – все кольчатые черви, членистоногие и все моллюски; Б – все кишечнополостные и иглокожие; В – плоские круглые черви, немертины
Каждая фагоцителла может лечь на оральный полюс (А), на аборальный полюс (Б), либо параллельно субстрату (В). А Б В субстрат А – все кольчатые черви, членистоногие и все моллюски; Б – все кишечнополостные и иглокожие; В – плоские круглые черви, немертины
 Типы яйцеклеток и их дробление
Типы яйцеклеток и их дробление
 и связанные с ними типы гаструляции (з – о):") Типы бластул (а – ж) и связанные с ними типы гаструляции (з – о): а — равномерная моруда; б — равномерная стерробластула; в — равномерная целобластула; г — неравномерная целобластула; д — неравномерная стерробластула; е — дискобластула; ж — плакула; з — морульная деламинация; и — клеточная деламинация; к — мультиполярная иммиграция; л — униполярная иммиграция; м — инвагинация; к — эпиболия; о — изгибание плакулы. Энтодерма отмечена пунктиром.
Типы бластул (а – ж) и связанные с ними типы гаструляции (з – о): а — равномерная моруда; б — равномерная стерробластула; в — равномерная целобластула; г — неравномерная целобластула; д — неравномерная стерробластула; е — дискобластула; ж — плакула; з — морульная деламинация; и — клеточная деламинация; к — мультиполярная иммиграция; л — униполярная иммиграция; м — инвагинация; к — эпиболия; о — изгибание плакулы. Энтодерма отмечена пунктиром.
 Сочетания вариантов: типов яйцеклеток, дробления яйцеклеток, типов гаструляции, образование третьего зародышевого листка, вторичной полости тела (целома) обеспечивают морфологическое разнообразие в эволюции многоклеточных животных
Сочетания вариантов: типов яйцеклеток, дробления яйцеклеток, типов гаструляции, образование третьего зародышевого листка, вторичной полости тела (целома) обеспечивают морфологическое разнообразие в эволюции многоклеточных животных
 Первое многоклеточное животное, оставшейся на структурном уровне паренхимулы – однослойный трихоплакс Trichoplax adhaerens
Первое многоклеточное животное, оставшейся на структурном уровне паренхимулы – однослойный трихоплакс Trichoplax adhaerens
 Строение Trichoplax adhaerens
Строение Trichoplax adhaerens
 Тема: Царство Metazoa подцарство Prometazoa
Тема: Царство Metazoa подцарство Prometazoa
 На уровень многоклеточности выходят только те одноклеточные, которые имеют оогамию и гаметическую редукцию и три этапа онтогенеза (индивидуального развития организма)
На уровень многоклеточности выходят только те одноклеточные, которые имеют оогамию и гаметическую редукцию и три этапа онтогенеза (индивидуального развития организма)
 Прогамный – образование гамет (половых клеток); 2) Сингамный этап – слияние") Этапы онтогенеза: 1) Прогамный – образование гамет (половых клеток); 2) Сингамный этап – слияние мужской и женской гаметы; 3) Метагамный этап – а) палинтомическое деление клетки (деление без увеличения массы клетки – клеток много но мелких); б) монотомическое деление клетки (деление на две дочерние клетки);
Этапы онтогенеза: 1) Прогамный – образование гамет (половых клеток); 2) Сингамный этап – слияние мужской и женской гаметы; 3) Метагамный этап – а) палинтомическое деление клетки (деление без увеличения массы клетки – клеток много но мелких); б) монотомическое деление клетки (деление на две дочерние клетки);
 У некоторых из многоклеточных животных количество клеточных делений ограничено. Конечное количество клеток является видовым признаком (например у коловраток).
У некоторых из многоклеточных животных количество клеточных делений ограничено. Конечное количество клеток является видовым признаком (например у коловраток).
 У свободноживущих животных все стадии перехода от одноклеточных к многоклеточным не прослеживаются. У многоклеточных (гетероклеточных) животных стадии перехода выявляются только на отдельных этапах онтогенеза.
У свободноживущих животных все стадии перехода от одноклеточных к многоклеточным не прослеживаются. У многоклеточных (гетероклеточных) животных стадии перехода выявляются только на отдельных этапах онтогенеза.
") Среди многоклеточных животных существуют пять типов с «отрицательными признаками» : нет дефинитивных (окончательно установленных) фаго- и цитобласта; нет рта, кишечника; никогда не было мускулатуры и нервного аппарата.
Среди многоклеточных животных существуют пять типов с «отрицательными признаками» : нет дефинитивных (окончательно установленных) фаго- и цитобласта; нет рта, кишечника; никогда не было мускулатуры и нервного аппарата.
 подцарство Prometazoa (Прометазои) 1. Надтип Plasmodiata тип Orthonectida тип Dicyemida") Царство Metazoa (Многоклеточные животные) подцарство Prometazoa (Прометазои) 1. Надтип Plasmodiata тип Orthonectida тип Dicyemida тип Myxosporae 2. Надтип Parenchimellata тип Placozoa (Пластинчатые) тип Spongia (Губки)
Царство Metazoa (Многоклеточные животные) подцарство Prometazoa (Прометазои) 1. Надтип Plasmodiata тип Orthonectida тип Dicyemida тип Myxosporae 2. Надтип Parenchimellata тип Placozoa (Пластинчатые) тип Spongia (Губки)
 тип Orthonectida Жизненный цикл ортонектиды
тип Orthonectida Жизненный цикл ортонектиды
 – слева и ее жизненный цикл. 1— 4 —") Строение Dicyemida (стадия первичного нематогена) – слева и ее жизненный цикл. 1— 4 — окружающие клетки четырех типов, 5 — осевая клетка, 6 — ядро, 7 — генеративные клетки (аксобласты), 8— 9 — бородавчатые и хвостовые клетки; а — двухклеточный зародыш, б — нематоген-основатель, в — первичный нематоген, г — ромбоген, д — инфузориген внутри осевой клетки ромбогена, е — инфузориформная личинка.
Строение Dicyemida (стадия первичного нематогена) – слева и ее жизненный цикл. 1— 4 — окружающие клетки четырех типов, 5 — осевая клетка, 6 — ядро, 7 — генеративные клетки (аксобласты), 8— 9 — бородавчатые и хвостовые клетки; а — двухклеточный зародыш, б — нематоген-основатель, в — первичный нематоген, г — ромбоген, д — инфузориген внутри осевой клетки ромбогена, е — инфузориформная личинка.
. А - Ortholinea diver gens; Б - Chloromyxum") Плазмодии разных видов миксоспоридий (из Шульмана). А - Ortholinea diver gens; Б - Chloromyxum leydigi; В - Sinuolinea dimorpha; ГMyxoproteus caudatus; ДSphaerospora irregularis; E Leptotheca agilis; ЖCeratomyxa ramosa; 3 Ceratomyxa drepanopsettae; И Ceratomyxa appendiculata; К Myxidium lieberkuhni
Плазмодии разных видов миксоспоридий (из Шульмана). А - Ortholinea diver gens; Б - Chloromyxum leydigi; В - Sinuolinea dimorpha; ГMyxoproteus caudatus; ДSphaerospora irregularis; E Leptotheca agilis; ЖCeratomyxa ramosa; 3 Ceratomyxa drepanopsettae; И Ceratomyxa appendiculata; К Myxidium lieberkuhni
 характерна отдельным стадиям жизненного цикла.") Надтип Plasmodiata - Стадия плазмодия (нет определённых контуров тела) характерна отдельным стадиям жизненного цикла. - Слабая выраженность разделения на клеточные пласты (разделение только зачаточное) - В жизненном цикле выделяются два поколения: а) трофическое (трофонт) – плазмодиальное строение; б) генеративное (гамонт) – имеет гетероклеточное строение
Надтип Plasmodiata - Стадия плазмодия (нет определённых контуров тела) характерна отдельным стадиям жизненного цикла. - Слабая выраженность разделения на клеточные пласты (разделение только зачаточное) - В жизненном цикле выделяются два поколения: а) трофическое (трофонт) – плазмодиальное строение; б) генеративное (гамонт) – имеет гетероклеточное строение
 Надтип Plasmodiata Имеются две клеточные ассоциации, которые можно ассоциировать с кинетобластом и фагоцитобластом
Надтип Plasmodiata Имеются две клеточные ассоциации, которые можно ассоциировать с кинетобластом и фагоцитобластом
 Ни у одной группы животных перешедшей к паразитизму не происходит утраты нервного аппарата и мускулатуры. Поэтому помещение Orthonectida и Dicyemida как приложения после плоских червей (Шарова И. Х. ) является необоснованным.
Ни у одной группы животных перешедшей к паразитизму не происходит утраты нервного аппарата и мускулатуры. Поэтому помещение Orthonectida и Dicyemida как приложения после плоских червей (Шарова И. Х. ) является необоснованным.
") Надтип Parenchimellata Первичная стадия всех многоклеточных животных (первый этап перехода то одноклеточности к многоклеточности) – стадия паренхимелла
Надтип Parenchimellata Первичная стадия всех многоклеточных животных (первый этап перехода то одноклеточности к многоклеточности) – стадия паренхимелла
 надтип Plasmodiata – без развития, поскольку это паразитическая") Возможные пути перехода к паренхимелле: 1) надтип Plasmodiata – без развития, поскольку это паразитическая группа (их плазмодий можно ассоциировать с двумя слоями, но к паренхимелле они не подошли). 2) Из свободноживущих ныне остались на уровне паренхимеллы – протероспонгия и трихоплакс.
Возможные пути перехода к паренхимелле: 1) надтип Plasmodiata – без развития, поскольку это паразитическая группа (их плазмодий можно ассоциировать с двумя слоями, но к паренхимелле они не подошли). 2) Из свободноживущих ныне остались на уровне паренхимеллы – протероспонгия и трихоплакс.
 Proterospongia sp. Trichoplax adhaerens
Proterospongia sp. Trichoplax adhaerens
 Губки – тип одних из наиболее примитивных многоклеточных животных.") Тип Spongia или Porifera (Губки) Губки – тип одних из наиболее примитивных многоклеточных животных.
Тип Spongia или Porifera (Губки) Губки – тип одних из наиболее примитивных многоклеточных животных.
 Тело губок строится из трёх типов клеток, которые представляют собой не ткани, а клеточные ансамбли. Два из них напоминают эпителий: 1) Пинакодерма (без жгутиковые уплощённые клетки) 2) Хоанодерма (жгутиковые, воротничковые клетки) Между пинакодермой и хоанодермой находится мезохил, который содержит органический матрикс.
Тело губок строится из трёх типов клеток, которые представляют собой не ткани, а клеточные ансамбли. Два из них напоминают эпителий: 1) Пинакодерма (без жгутиковые уплощённые клетки) 2) Хоанодерма (жгутиковые, воротничковые клетки) Между пинакодермой и хоанодермой находится мезохил, который содержит органический матрикс.
 Схематический разрез через стенку тела губки типа аскона. Вверху - наружная стенка тела, внизу - парагастральная полость (по Штемпелю): 1 - клетки, выстилающие наружную стенку тела и стенки поровых канальцев, 2 жгутиковые воротничковые клетки, 3 - яйцевая клетка в мезоглее, 4 - склеробласт с развивающейся спикулой, 5 пора, 6 - звездчатые клетки в мезоглее
Схематический разрез через стенку тела губки типа аскона. Вверху - наружная стенка тела, внизу - парагастральная полость (по Штемпелю): 1 - клетки, выстилающие наружную стенку тела и стенки поровых канальцев, 2 жгутиковые воротничковые клетки, 3 - яйцевая клетка в мезоглее, 4 - склеробласт с развивающейся спикулой, 5 пора, 6 - звездчатые клетки в мезоглее
: 1 - отводящий канал, выстланный плоскими клетками,") Жгутиковая камера пресноводной губки Ephijdatia (по Кестнеру): 1 - отводящий канал, выстланный плоскими клетками, 2 - отверстие, сообщающее жгутиковую камеру с каналом, 3 - археоцит, 4 - воротничковые жгутиковые клетки (хоаноциты)
Жгутиковая камера пресноводной губки Ephijdatia (по Кестнеру): 1 - отводящий канал, выстланный плоскими клетками, 2 - отверстие, сообщающее жгутиковую камеру с каналом, 3 - археоцит, 4 - воротничковые жгутиковые клетки (хоаноциты)
 Пинакодерма губок Экзопинакодерма Эндопинакодерма - клетки без кутикулярной выстилки (экзопинакоциты сходны с клетками погружённого эпителия плоских, круглых червей и немертин) - Без жгутиковые клетки выстилают псевдогастральную полость отдельных групп губок
Пинакодерма губок Экзопинакодерма Эндопинакодерма - клетки без кутикулярной выстилки (экзопинакоциты сходны с клетками погружённого эпителия плоских, круглых червей и немертин) - Без жгутиковые клетки выстилают псевдогастральную полость отдельных групп губок
. Под пинакоцитами очень часто располагаются миоциты") Экзо- и эндопинакоциты имеют микрофиламенты (сократительные белковые тяжи). Под пинакоцитами очень часто располагаются миоциты (сократительные клетки). Подошва губок образована базипинакоцитами. На поверхности экзопинакоцитов встречаются пороциты с микрофибриллами. Пороциты ведут в каналы, которыми пронизано тело губок.
Экзо- и эндопинакоциты имеют микрофиламенты (сократительные белковые тяжи). Под пинакоцитами очень часто располагаются миоциты (сократительные клетки). Подошва губок образована базипинакоцитами. На поверхности экзопинакоцитов встречаются пороциты с микрофибриллами. Пороциты ведут в каналы, которыми пронизано тело губок.
 Хоанодерма - выполняет гидрокинетическую функцию. Хоаноцит
Хоанодерма - выполняет гидрокинетическую функцию. Хоаноцит
 По расположению хуанодермы и пинакодерма выделяют три типа строения губок: А -аскон; Б - сикон; В - лейкон.
По расположению хуанодермы и пинакодерма выделяют три типа строения губок: А -аскон; Б - сикон; В - лейкон.
") Аскон – может быть самостоятельной губкой Сикон – как колония аскона (модульный тип строения) Лейкон – из сиконов (модульный тип строения)
Аскон – может быть самостоятельной губкой Сикон – как колония аскона (модульный тип строения) Лейкон – из сиконов (модульный тип строения)
 Строение губок
Строение губок
, спонгиобласты, миоциты, лофоциты, археоциты, гаметоциты, амебоидные") В мезохилле губок находятся колленциты, склеробласты (спикулы, иглы), спонгиобласты, миоциты, лофоциты, археоциты, гаметоциты, амебоидные клетки. У губок отсутствует дефинитивная дифференцировка клеток и клетки взаимопревращаемы. Поэтому у губок нет тканей и нет зародышевых пластов.
В мезохилле губок находятся колленциты, склеробласты (спикулы, иглы), спонгиобласты, миоциты, лофоциты, археоциты, гаметоциты, амебоидные клетки. У губок отсутствует дефинитивная дифференцировка клеток и клетки взаимопревращаемы. Поэтому у губок нет тканей и нет зародышевых пластов.
 Дефект дифференцировки губок в том, что она не окончательна и может быть этим объясняется их (губок) высокая адаптивность – губки – одна из самых древних групп.
Дефект дифференцировки губок в том, что она не окончательна и может быть этим объясняется их (губок) высокая адаптивность – губки – одна из самых древних групп.
 У трихоплакса клетки полифункциональны, а у губок имеется специализация клеток (клетки более качественно выполняют одну свою функцию)
У трихоплакса клетки полифункциональны, а у губок имеется специализация клеток (клетки более качественно выполняют одну свою функцию)
 На уровне губок проявляются интегративный механизм трёх уровней обеспечивающий реакцию губки, как единого целого организма: 1) Трансмембранная передача информации. 2) Цитоплазматические мостики между клетками мезохилла. 3) Перемещение сигнальных молекул в органическом матриксе мезохилла.
На уровне губок проявляются интегративный механизм трёх уровней обеспечивающий реакцию губки, как единого целого организма: 1) Трансмембранная передача информации. 2) Цитоплазматические мостики между клетками мезохилла. 3) Перемещение сигнальных молекул в органическом матриксе мезохилла.
 Ephidatia blembingia. А - формирование капсулы и принос") Образование геммул у бадяг (сем. Spongillidae) Ephidatia blembingia. А - формирование капсулы и принос амфидисков; Б - сформированная геммулa. cel - клеточное содержимое геммулы, chit - хитиноидная капсула, h - слой клеток, строящий капсулу и воздухоносный слой, am. c - внедряющиеся между клеток этого слоя амебоидные клетки с амфидисками, p - поровое отверстие; амфидиски (am) в сформированной геммуле заключены в воздухоносную ткань.
Образование геммул у бадяг (сем. Spongillidae) Ephidatia blembingia. А - формирование капсулы и принос амфидисков; Б - сформированная геммулa. cel - клеточное содержимое геммулы, chit - хитиноидная капсула, h - слой клеток, строящий капсулу и воздухоносный слой, am. c - внедряющиеся между клеток этого слоя амебоидные клетки с амфидисками, p - поровое отверстие; амфидиски (am) в сформированной геммуле заключены в воздухоносную ткань.
 Целобластула Clathrina reticulum Амфибластула губки Leucosolenia variabilis Паренхимула Mycale lorenzi
Целобластула Clathrina reticulum Амфибластула губки Leucosolenia variabilis Паренхимула Mycale lorenzi