3 лек2016.ppt
- Количество слайдов: 77



Лекция 3. n «Тұзды жағдайға өсімдіктердің бейімделу механизмдері» n n n 1. Тұзды стрестің сипаттамасы 2. Галофиттер, гликофиттер 3 Клетканың дегидратация механизмдері 4. Тұздың токсикалық әсері 5. Иондық насостардың маңызы

n Тұзды стреске өсімдіктердің бейімделу механизмдері» . n Жердің бетінде 1/15 бөлігі тұзды жерлер болып саналады. n n n n Тұзсыз - егер неорганикалық иондардың өлшемі 0, 2% аспаса Аз тұзды - 0, 2 - 0, 4 % Орташа тұзды жерлер - 0, 4 -0, 7%, Қатты-күшті тұзды жерлер - 0, 7 -1, 0% Солончактар - неорганикалық иондардың мөлшері 1, 0%-тен көп, Тұзды топырақта әдетте катиондардан Na көп болады, бірақ Мg 2+ мен Са 2+ өда көп жерлер болады. Аниондардың ішінде Сl және SO 4 көп болады, бірақ карбонаттар да көп болады. Галофиттер (от греч. galos — тұз, phyton — өсімдік), - тұзды жерде өсетін және тұздың жоғары концентрациясына үйренген өсімдіктер. Гликофиттер (греч. glycos — сладкий, рhyton — растение) Тұздық жерде өспейтін өсімдіктер.

n n Ауыл-шаруашылық өсімдіктер көпшілігінде гликофиттерге жатады. Олар тек тұздың аз концентрациясына шыдайды, тек кейбіреулер - орташа концентрациясына – мақта (Gossipium hirsutum), сұлы (Оvena sativa), қара бидай (Secale sereale) и бидай. Тұздың мөлшері көбейгенде егістің өнімі төмендейді. Галофиттер қатты тұзды жерлерде, солончактарда өсе алады. Әдеттегі галофиттерге маревые тұындысы (Chenopodiaceae, солерос европейский (Salicornia europaea), кохия распростертая (Косhia prostrata), петросимония трехтычинковая (Petrosimonia triandra) және т. б. Тұздың көп мөлшері олардың өсуін ынталандырады.

Гликофиттер Тұзды жерде өсетін және тұздың жоғары концентрациясына үйренген өсімдіктер n. Мақта n (Gossipium hirsutum), nсұлы (Оvena sativa), nқара бидай (Secale sereale), бидай. Галофиттер Қатты тұзды жерлерде, солончактарда өсе алады Chenopodiaceae Salicornia europaea, Косhia prostrata, Petrosimonia triandra

n Тұздардың зақымдандыратын әсері. ТҮЗДАР n Клетканың гомеостаз бұзылады Биополимерлер зақымданады Өсу процестер баяулайды

n n n n n Клетка деңгейінде болатын құбылыстар. Клетканың дегидратациялануы. Біріншіден тұзды жағдайда клеткалар дегидратацияланады. А. Ф. Шимпер ХХ ғасырдың басында Тұзды жерлерде топырақ ерітіндінің жоғары осмостық қысымынан өсімдіктер суды жоғалататынын көрсетті. Тұзды жерлерде топырақ ерітіндінің су потенциалы төмендегенде клеткалар дегидратацияланады. Осмостық әсері тұздың түрінен тәуелсіз болады. Ең маңызды көрсеткіштер: - иондардың концентрациясы; -осмостық активті заттардың концентрациясы.

n n Б. П. Строгонов - ХХ ғасырдың 50 -ші жылдарда хлоридтармен және сульфаттармен ластанған топырақтар өсімдіктерге әр түрлі әсер етеді. Осыған қарай әр түрлі иондар өзгеше зақымдану әсер етеді деген концепция ұсынған. Кәзіргі заманда клетка деңгейінде осмостық және токсикалық әсерлер – басты зақымдайтын факторлар деп саналады. Токсикалық әсерлердің табиғаты онша көп зерттелмеген. Иондардың жоғары концентрациясы Клетка мембраналарының зақымдануы Ферменттердің ырықтылығының төмендеуі Минералдық элементтердің сору процестер нашарлайды коміртекті ң ассимиляциалануы нашарлайды Клетканың Бөлінуі нашарлайды

n n n n n Хлордың токсикалық нысандары туралы аз белгілі. ХЛОР РНК және аниондық метаболиттермен бикарбонаттармен, карбоксилаттермен, қанттардың фосфаттарымен-байланысу жерлерде бәсекеленеді. Na иондар катиондар К+, Са 2+ и Mg 2+. байланысатын сайттармен қосылады. Макромолекулаларды зақымдандыратын факторларға оттегінің активті түрлері жатады: - синглеттік оттегі, - супероксид-радикал, - гидроксил-радикал, - оттегінің асқын тотығы.

n n n Олардың концентрациясы су тапшылығында, тұзды жағдайда көбейеді. Бұл жағдайда токсикалық эффектері иондардың тікелей әсерілермен байланыссыз. Олар екінші ретті процестермен байланысты - биополимерлер оттегінің активті түрлерімен байланысып, клетканы зақымдатады.

n Бүкіл өсімдік деңгейінде болатын құбылыстар. n 1. Тұзрдың үлкен концентрациялары n n өсімдіктің өсуін тежейді. Олар гормон ЦК азайғанмен және АБҚ көбейгенмен байланысты. Клеткаларда 1 -аминоциклопропан-1 -карбон қышқылы көбейеді – ол этиленнің алдынғы қосындысы. Гормондардың мөлшері өзгеруі өсімдіктердің тұрақтылығын жоғарлатады. Кейбір зерттеушілер өсімдіктердің өсуінің баялауының себебі тұздың зақымдайтын әсерінен емес, оның себебі - адаптациялық гормондық жауаптармен байланысты деп санайды. Тұрақтылықтың деңгейі өсудің жылдамдығымен теріс корреляция қатынасында. n n Баяу өсу - өсімдіктің қорғаныш реакциясы, көп ресурстарды құрылыс белоктарды, энергияны сақтап қалады.

Бүкіл өсімдік деңгейінде болатын құбылыстар Өсу тежеледі ЦК азаяды АБҚ көбейеді 1 -аминоциклопропан-1 -карбон қышқылы көбейеді – ол этиленнің алдынғы қосындысы

n n 2. Устьицаның өткізгіштігі азаяды. Тұздық жағдайда топырақ ерітінідісінің төмен су потенциалы устьицаларды қоршаған клеткалардың суды жоғалтуына әкеледі. Бұл құбылыс устьицаның қуысының арасын кішірейтеді, содан СО 2 –нің устьицадан кіру жылдамдылығын, және транспирацияны төмендетеді. Устьицаны қоршаған клеткалаарда тургорық қысым төмендегенге тұздан басқа, АБҚ түғызатын процестер әсер етеді. Бұл гормон клеткаларда ион-тасмалдаушы белоктардың активтілігін өзгертеді.

АБК АТФазаның активтігі тежеледі Клетка мембранасы деполяризацияға ұшырайды Калий-кіретін каналдар активтенеді Натрийшығатын каналдар активтенеді Анион-шығатын каналдар активтеленеді К+, Cl- - шығады клеткадан Устьица қоршайтын клеткалар тургорын жоғалдтады Устьицалар жабылады

n n n n Тұздардың осмостық әсеріне қарсы адаптациялар Бұл адаптациялардың негізініде су тапшылыққа бейімделу әдістері болады: осмолиттердің биосинтезі және неорганикалық иондардың вакуолде жиналуы. Иондық гомеостазды сақтау – тұздың токсикалық әсерінен қашу стратегиясы. Әдетте Nа мен Сl концентрациясы галофиттерде және гликофиттерде шамасында бір деңгейде болады. Галофиттер тұзды жерде өскеннен олар клеткада тұздың аз концентрациясын ұстап тұруға үйренген – бірнеше моль/кг. Бірақ гликофиттер клеткаларда тұзды жерде иондардың қалыпты концентрацияларды ұстап тұралмайды. Иондық гоместазды ұстап тұру - токсикалық әсерінен қашу стратегиясы эволюцияның ерте кезеңдерінде қалыптасқан.

n n Кәзіргі заманда тек галофильдік өсімдіктер бұл стратегияны қолданбайды. Олардың белоктары ерекше құрылымды болады. Ерекше құрылым белоктарға Na иондары көп жағдайда өз фунцияларын орындай алатын мүмкіншілік бередіHalobacterium halobium.

n n Цитозольдың суда ерігіш белоктардың және мембранамен, рибосомалар мен байланысқан белоктардың дикарбондық АҚ-дар көп болады (глутамин және аспарагин қышқылы). Нейтральдық ортада қышқыл белоктар теріс зарядталған. Олар катиондармен нейтралдану керек. Бұндай белоктарға цитоплазмада көп оң зарядталған иондары өте қажет. Ондай болмаса электростатикалық итеру нәтижесінде белоктар денатурацияланып кетуі мүмкін. Галофильдік бактериаларда Na+, К+ концентрациялары бірнеше мольдарға жетеді. Бұндай жоғары концентрациялар белоктардағы гидрофобтық байланыстарды қатайтады.

n Цитоплазмада иондардың аз концентрациялары сақталуы кіретін және шығатын иондардың ағымыдарының балансында негізделген. n n Na-дің цитоплазмаға пассивтік кіруі клеткадан активтік экспортының балансымен белгіленеді.
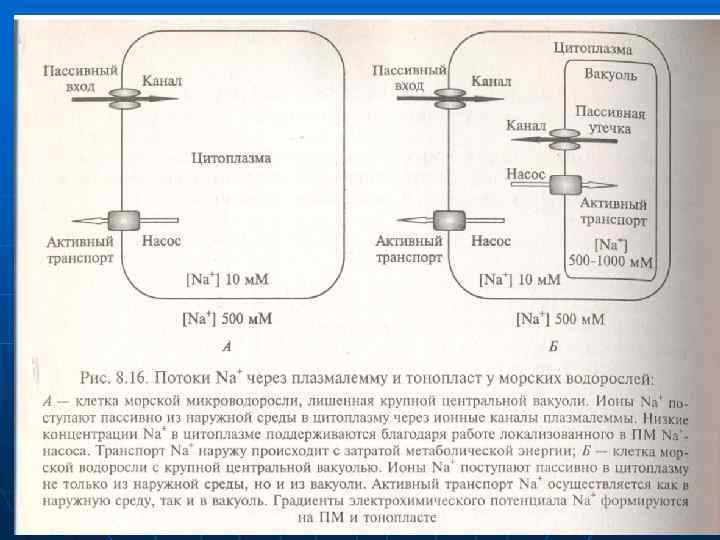

n n n Na және Сl топырақ ерітіндісінен клеткаларға кіруі. Na және Сl топырақ ерітіндісінен тамырдың эпидермис және қабық клеткаларының цитоплазмасына электрохимиялық потенциал градиенті арқылы кіреді Na және Сl сыртқы ортадан клеткаға тасмалдайтын плазмалемманың белоктары. Na және басқа иондар мембрананың қосқабатынан кіре алмайды. Олар иондық канал және тасмалдаушылар арқылы кіреді. Na- каналдар, жануарлардың Na-каналдарына ұқсас. Тамыр клеткаларда кейбір белоктар табылған. Олар арқылы иондар ЦП-ға кіреді. Ондай белоктарға ПМ-ның К+-каналдар жатады.
 – каналдар. ПМ гиперполяризацияланғанда активтеленеді, ол теріс")
n n n 1. К+-кіретін (inward rectifying) – каналдар. ПМ гиперполяризацияланғанда активтеленеді, ол теріс зарядтар көбейгенде. Олар К/Na селективтігі өте жоғары болады. Бірақ Na-дің жоғары концентрацияларда оны да өткізеді. 2. К+- шығатын (outward rectifying)—каналдар — ПМ гиперполяризацияланғанда активтеленеді, ол оң зарядтар көбейгенде. Калийді цитоплазмадан шығарады. 3. Потенциал-тәуелсіз катиондық VIC (voltageindependent channel)-каналдар —К/Na селективтілігі төмен. Олардың өткізгіштігі трансмембрандық потенциалдан тәуелсіз болады. 4. Жоғары-афинндік К-тасмалдаушы механизмдер. Naдің ЦП-ға кіруінде НКТ 1 -тасмалдаушы қатысады. — К+, Na+-симпортер. Бұл белок 2 ионды бір жаққа тасмалдайды (симпорт). К+, Н+ -симпортер
 – каналдар. К+- шығатын")
К/Na селективтігі төмен К/Na селективтігі өте жоғары К+-кіретін (inward rectifying) – каналдар. К+- шығатын (outward rectifying) —каналдар ПМ деполяризацияланғанда активтеленеді ПМ гиперполяризацияланғанда активтеленеді Теріс зарядтар көбейеді n n Оң зарядтар көбейеді Потенциал-тәуелсіз катиондық VIC (voltageindependent channel)-каналдар Олардың өткізгіштігі Трансмембрандық потенциалдан тәуелсіз болады.

n n n Жоғары-афинндік К-тасмалдаушы механизмдер n n n НКТ 1 -тасмалдаушы Na-дің ЦП-ға кіруіне қатысады К+, Na+-симпортер. n n n Бұл белок 2 ионды бір жаққа тасмалдайды (симпорт). К+, Н+ -симпортер n Тек VIC-каналдардың натрий иондарының кіруіне үлесі көп. Сондықтан гендік инжененрия әдістерімен бұл белоктармен манипуляция жасау ыңғайлы болады. Бірақ олар молекулалық гендік деңгейінде әлі белгіленген. Бұл каналдар арқылы натрийиондарымен Са-иондары бәсекеленіп өте алады. Тұзды жерлерде Са-иондарын қолданып, өсімдіктердің өнімділігін n жоғарылатуға болады.


n n n Сl-иондарының кіруі Хлор индары клеткаға анион-каналдар арқылы электрохимиялық градиент бойынша кіреді. Na. СI көбейгенде Na тезірек клеткаға кіреді де электрохимиялық потенциалдың градиенті оң зарядты болады. Соңдан соң хлор клеткаға пассивтік тасмаладанады.

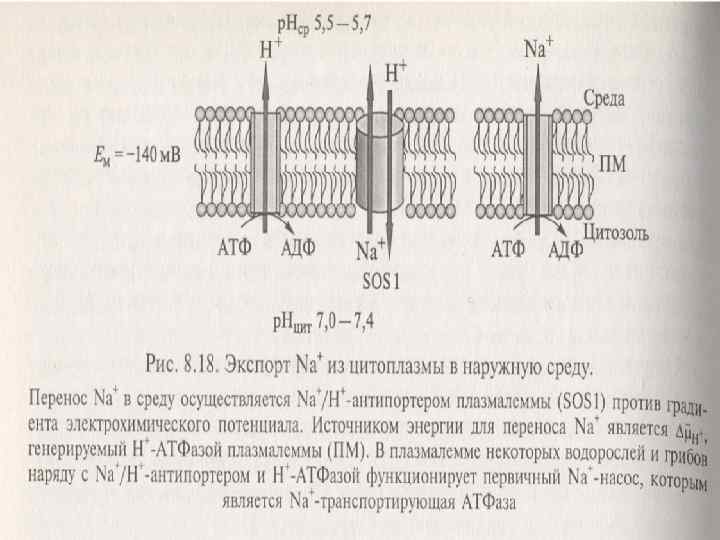
n n n n n Na+ және СI- цитоплазмадан шығуы. Экспорттың 2 түрі болады: - ПМ-дан отіп клетка арлығына өту -Вакуольға тонопласт арқылы өту. ПМ-ның Na/Н-антипортері — ионалмастыру диффузияның механизмі. Ол натрийді Н+ -протондарға ауыстырады. Ол протонның потенциалы бойынша μН+ электрохимиялық градиентке қарсы жүреді. Бұл екінші реттік активті тасмалдануға жатады. μн+ потенциалын Н+-АТфаза туғызады. Бұл жоғары сатыдығы өсімдіктердің ПМ-да, балдырларда және ашытқыштарда болады
. Кейбір ашытқыштардың және галотолерантнттық балдырлардың")
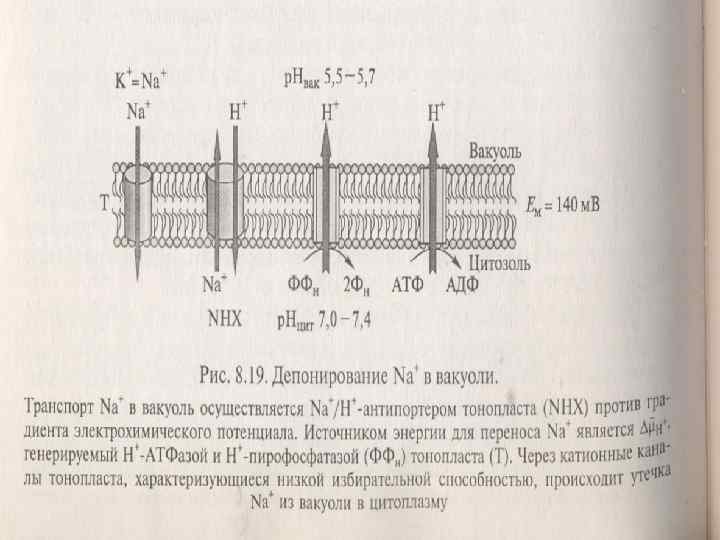
n n n n n Бірінші реттік Na-насос (Na+-АТФаза). Кейбір ашытқыштардың және галотолерантнттық балдырлардың ПМда 1–ші реттік Na –насостар табылды. Олар фосфордың энергиясын пайдаланатын Na+тасмалдайтын АТФазалар. Олар Р-тип АТФазаларға жатады. Жоғары сатылы өсімдіктерде олар болмайды. Тонопластың Na/Н-антипортері. Тонопласта Na/Н-антипортер және Н-АТФазада жұмыс істейді. Бұл АТФаза V-типке жатады, олар Нпротондарды ЦП-дан вакуольге тасмалдайды. ПМ мен тонопластағы Сl-ды экспорттайдын жүйелер. Хлордың экспорты анион каналдар арқылы жүзеге асады. Тасмалданудың бағыты ПМ және тонопластың эл/химиялық потенциалымен белгіленеді. Н-АТФаза ПМ, Н-АТФаза тонопластта и Н+пирофосфатаза тонопластта — эл/химиялық потенциалды туғызады.

n n n К+/ Na+ - селективтілігі — тұзға тұрақтылығының индикаторы. Топырақ ерітіндісінде К+ мен Na+ тамырмен соруына бәсекелеседі. Өсімдіктердің К/Na қатынасының жоғары болып ұстайтын қабілеті олардың тұзға тұрақтылығын көрсетеді. Өсімдік клеткаларда ол иондық қатынас иондық тасмалдаушы жүйелерінің таңдаушылығымен және натрийдің экспортымен байланысты

n n n Су тапшылығына және тұзды стреске қарсы клеткалық төзімділік механизмдері 1. Тұзды жағадйға ең негізгі адаптациялық стратегиясы – ол Na и СI төмен концентрациясын клеткада сақтау. 2. Осмолиттердің синтезі клеткаға керек судың деңгейін сақтайды. 3. Транспирацияның қарқындылығы жоғары болғанда заттар апопласттық жолмен клеткаға кіреді. Олар гидростатикалық градиент бойынша тамырға кіреді. Транспирацияның қарқындылығы төмен болғанда, иондар симпласттық жолмен жүреді. Онда олар иондық каналдар, тасмалдаушылар және ионтасмалдаушы АТФазалар арқылы жүреді.

n n n Ксилемаға иондар паренхималық клеткалардың ПМасынан кіреді. Гликофиттерде ксилеманы қоршаған клеткалардан калий-шығаратын канал арқылы тасмалданады. Галофиттерде тасмалданатын иондардың ішінде натрий көп болады. Хлор иондары анион каналдар арқылы өтеді. Кейбір галофиттерде тұз шығаратын бездер болады- кермек (Limonium gmelinii). Гликофиттерде Na мен СI-дың кіруі осмолиттер арқылы тежеленеді.

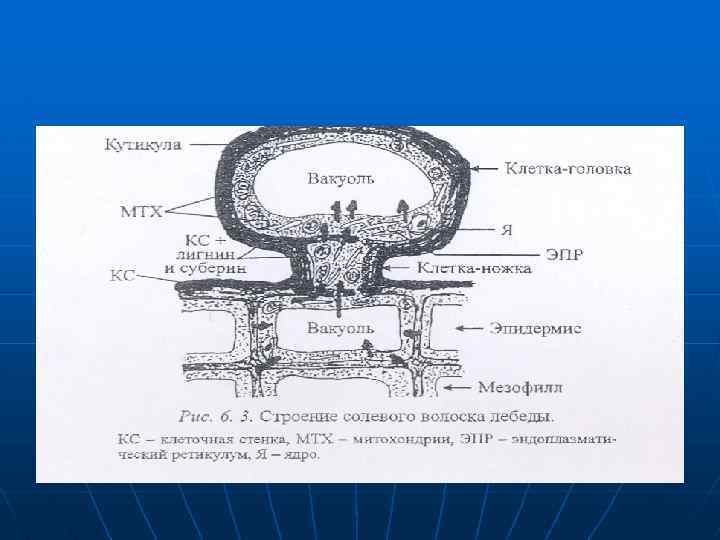
n n n Поглощаемые соли могут концентрироваться и в вакуолях клеток головок специализированных волосков (рис. 6. 3). Когда в вакуолях головки накапливаются соли головка обрывается, на ее месте несколько раз в течение роста листа образуется новая головка. Солевые волоски состоят из ножки и головки, В собирающей клетке-ножке, не имеющей центральной вакуоли много митохондрий и цистерны ЭПР.

n n n Реакции растений на засоление Основная реакция организма на засоление — нарушение водного потенциала и минерального обмена. Легкорастворимые соли повышают осмотическое давление почвенного раствора и создают физиологическую сухость, от которой растения страдают так же, как от почвенной засухи. Поэтому повреждения при засолении очень сходны с теми, что наблюдаются при засухе, т. е. прежде всего увеличивается значение соотношения связанной и свободной воды и повышается осмотический потенциал клетки. - Снижение интенсивности синтетических процессов, и усиление гидролиза. Если степень засоления среды превышает детальный порог, то синтетические функция подавляются настолько сильно, что растение гибнет. Если такой порог не достигается, то синтетические реакции

n n n - Торможение роста. - Изменяется и соотношение фитогормонов: накапливается АВК, что ведет к закрыванию устьиц и способствует ограничению потери воды. - Фотосинтез ингибируется, когда Na. Cl в высоких концентрациях накалливается в хлоропластах. Поскольку фотосинтетическая ЭТЦ относительно нечувствительна к соля. их неблагоприятный эффект сказывается в основном на углеродном метаболизме и фотосинтетическом фосфорилировании. Снижается также содержание хлорофилла (больше у неустойчивых растений). Образование хлорофилла тормозится, прекращается синтез крахмала. Торможение фотосинтеза корреллирует с замедлением транспорта ассимилятов. Солевой стресс увеличивает образование органических кислот и аминокислот.

n n n Реакция дыхания на избьтток солей неодназначна. Чаще всего интенсивность дыхания сначала возрастает. поскольку поддержание ионного гомеостаза в условиях засоления требует значительных затрат энергии. Дополнительная энергия необходима для роста клеток в условиях засоления. Клетки в стадии растяжения для обеспечения тургора должны обладать определенным осмотическом потенциалом, который обусловлен энергозависимым транспортом ионов из апопласта через плазмалемму и тонопласт в симпласт и вакуоль. Когда клетка заканчивает растяжение, дальнейшее поступление солей не нужно, оно оказывается даже вредным, поскольку способствует еще большему повышению концентрации солей. Поэтому на данном этапе осуществляется

n Увеличивается доля ПФП. n n Активность терминальных оксидаз падает, а флавиновых дегидрогеназ, наоборот, возрастает. n Происходит ингибирование переноса электронов в ЭТЦ причем Na. Cl, подобно ДНФ, выступает n n n -При сильном засолении энергетическая эффективность дыхания снижается. -Отмечается также усиление активности ПОЛ, в результате торможения функционирования антиоксидантных систем. - Изменяет ся транспорт ионов. Именно нарушения в поглощении и транспорте ионов составляют главные различия симптомов засоления и засухи. Возрастание содержания определенных солей приводят к их преимущественному поступлению в ткани растений, тогда как поглощение других тормозится. Содержание натрия — основного катиона при засолении в клетках повышается, а калия и магния снижается.

n n Отличие засоления от засухи - снижение активности ферментов и других белков в этом случае происходит не только вследствие истончения гидратных оболочек, как при водном дефиците, но и в результате прямого токсического, высаливающего действия солей. В зависимости от типа засоления определенным образом изменяется морфология растений, что обусловливает особенности физиологических процессов. При хлористом засолении растения внешне приобретают суккулентные черты: развиваются водоносные мясистые ткани. Интенсивность фотосинтеза таких растений невысока, замедлено поступление воды: интенсивность дыхания при расчете на единицу сырой массы снижена. хотя при расчете на единицу сухой массы приближается к величинам, характерным для мезофитов.

n n n При сульфатном засолении растения обнаруживают черты ксероморфности, т. е. фотосинтез и дыхание у них становятся интенсивнее, появляется мелкоклеточность и т. п. Сульфатное засоление нарушает обмен серы: в растениях накапливается много сульфоксидов. сульфонов, сульфоновых кислот. Эти соединения легко проникают через мембраны, нарушая их целостность, а на организменном уровне способствуют увяданию растений.

n n n Особенности приспособления растений n к условиям засоления Концентрация солей выше 5% летальна для большинства растений. Однако существуют виды, адаптированные к произрастанию даже в условиях сильного засоления. При этом используется различная стратегия. Растения пытаются избежать накоплекния солей (даже при их интенсивном поступлении) рис. 6. 1) или приспосабливаются к их высокой концентрации. Избежание засоления Для создания высокого осмотического давления и усиления поступления воды растения могут поглощать большое количество солей, однако для защиты цитоплазмьт от избытка солей их необходимо концентрировать в вакуолярном соке. Высокое содержание ионов в почве при засолении создает предпосылки для их пассивного поглощения по градиенту концентрации, т е, без затрат АТФ,
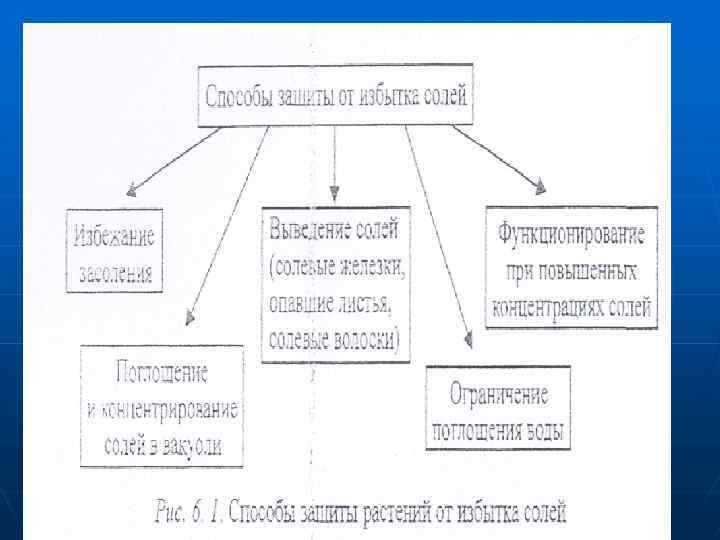

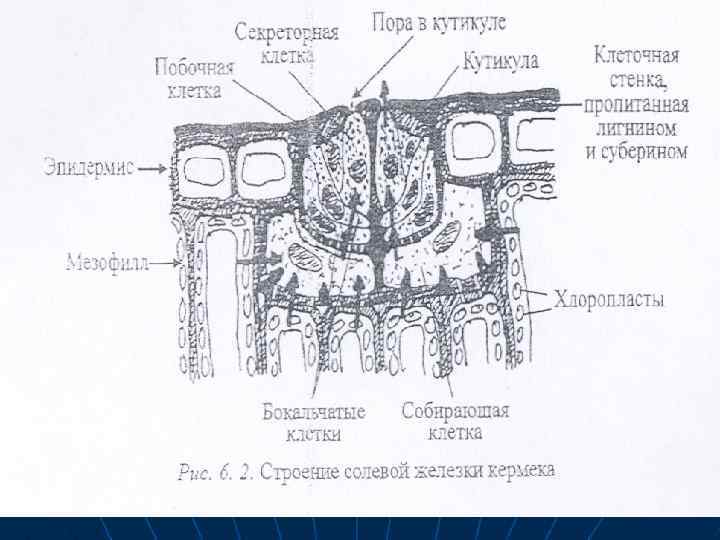
n n n Поглошенные соли выводятся также из клеток с водой с помощью специализированных соленых железок, или избыток солей удаляется с опавшими листьями, Солевые железы листьев и стеблей растений выделяют избыток ионов при их росте в высокосолевой среде. Собирательные клетки железок мало чем отличаются от элидермалных, но клеточная стенка у них значительно утолщена и лигнифицирована. Эти клетки связаны многочисленными плазмодесмами с окружающими их клетками мезофилла и эпидермиса, а также с бокальчатыми клетками. Секреторные, побочные и бокальчатые клетка также связаны друг с другом через плазмодесмы, их клеточные стенки пропитканы не только лигнином и суберином, что исключает апопластный транспорт.
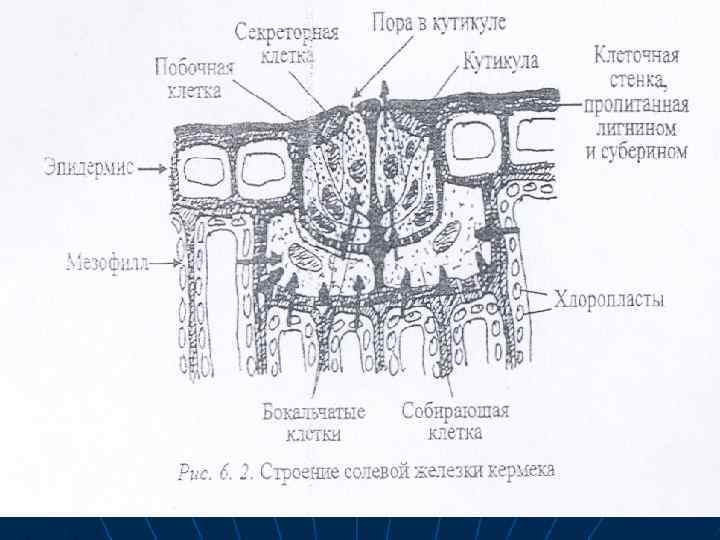

n n n Эти клетки железки находятся в своеобразной капсуле из лигнифицированной и суберинизированной клеточной стенки, которая таким образом изолируется от клеточных стенок окружаюшвх клеток. Снаружи железы покрыты мощной кутикулой, которая в зоне секреторных клеток имеет поры. Через эти поры и происходит секреция солей. В секреторных клетках имеются многочисленные рибосомы. митохондрии, много мелких вакуолей, но нет хлоропластов и запасных веществ. Соли постукают в корень, вместе с током воды двигающихся по сосудам ксилемы, а затем по апопласту тканей листа, что предохраняет детоксикацию из клеток от избытка солей. В собирающих клетках при участии ионных насосов соли переходят в симкласт и до выделения наружу

Благодаря деятельности этих и других насосов происходят активное выделение ионов хлора, натрия, кальция, магния, сульфата, . Помимо ионов через поры в кутикуле теряются значительные количества воды. В сухую погоду вода из выщелоченного раствора быстро испаряется и растение покрывается налетом солей.

n n n Поглощаемые соли могут концентрироваться и в вакуолях клеток головок специализированных волосков (рис. 6. 3). Когда в вакуолях головки накапливаются соли головка обрывается, на ее месте несколько раз в течение роста листа образуется новая головка. Солевые волоски состоят из ножки и головки, В собирающей клетке-ножке, не имеющей центральной вакуоли много митохондрий и цистерны ЭПР.

n n n Поглощаемые соли могут концентрироваться и в вакуолях клеток головок специализированных волосков (рис. 6. 3). Когда в вакуолях головки накапливаются соли головка обрывается, на ее месте несколько раз в течение роста листа образуется новая головка. Солевые волоски состоят из ножки и головки, В собирающей клетке-ножке, не имеющей центральной вакуоли много митохондрий и цистерны ЭПР.
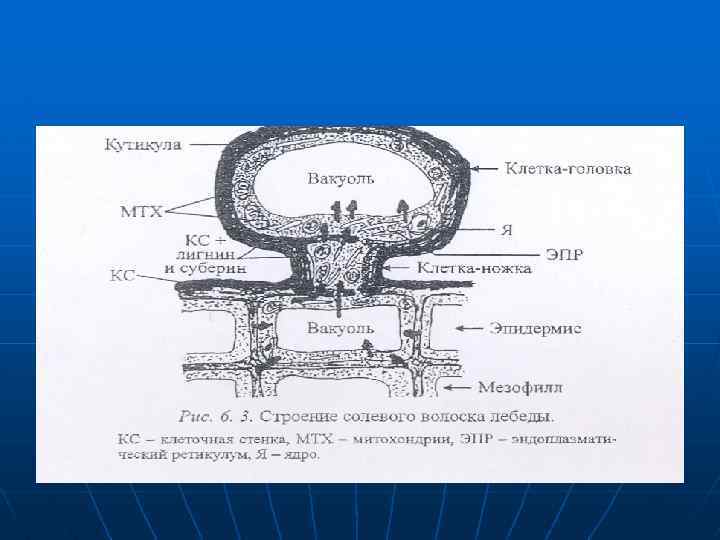

n n В верхней секреторной клетке — головке, 95% объема цитоплазмы занимает центральная вакуоль. Ножка связана плазмодесмами с головкой и эпидермальными клетками Клеточные стенки обеих клеток волоска утолщен и в значительной степени лигнифицированы, а также пропитаны суберином, но препятствует передвижению солей из апопласта в симласт. Снаружи волосок имеет толстую кутикулу. которая препятствует испарению воды

При активном участии ионных насосов соли проникают в цитоплазму эпидермальных клеток, перемещаются по симпласту в ножку и попадают в вакуоль клетки-головки. Накопление солей в клеточном соке происходит благодаря акивному ионнному транспорту на тоноаласте. Концентрация солей в вакуоли головки может в 60 раз превышать таковую в клетках эпидермиса и эквивалентна 5 -6 моль/л натрия и калия 9 -9. 5 моль/л хлора. в шггоалазме iекьшается

Вакуоль содержит также большое количество оксалата. Кроме того, волоски выделяют катионы: Са. и анионы - НСОЗ, SO 4 2 -. Солевые волосы теряют очень мало воды, но зффектавность выделения солей у них ниже, чем у соленых железок. Уровень солей

n n n Уровень солей в цитоплазме понижается при сбрасывания старых листьев. через гидатоды вместе с секрецией воды (особенно если нет значiпе. тьного водного дефтаiата). Локальное снижение уровня солей происходит и при выделении их в сосуды ксилемы и/или флоэмы и дальнейшем передвижении по растению. Ограничено поглошение солей клетками корней вследствие соленепроницаемости мембран. Повышается специфичность ионных каналов в основном калиевых. Уменьшается общее число. Происходят структурные перестройки в липидах мембан. появляются пропитанные суберином эндодермальные клетки, которые тормозят поступление солей по апопласту. Однако мембраны не проницаемы для солей лишь при незначительном засолении. В случае сильного засоления соли проникают внутрь клеток. В этих хсловкях начинают действовать другие

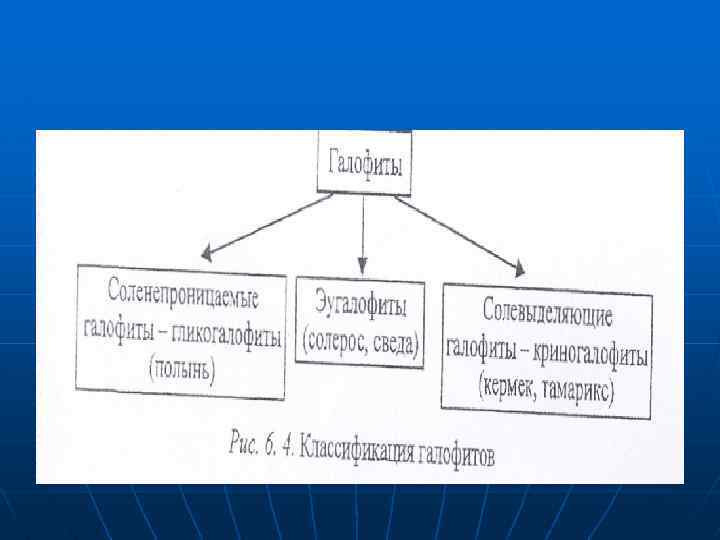
n n n Функционирование при повышенных n концентрациях солей Растения могут также а приспосабливаться к высоким конценрациям солей. Такие растения. толерантные к засолению, называются галофитами (галос — соль. фитос — растение). Галофиты классифющруются следующим образом (рис. 6. 4).
 — наиболее солеустойчивые растения, нак апливают в вакуолях")
n n Эугалофиты (настоящие. или соленакапливающие) — наиболее солеустойчивые растения, нак апливают в вакуолях соли в значительных концентрациях. Растут на влажных засоленных почвах, как солокчаки по морским побережьям и по берегам временных и постоянных соленых озер. Проницаемость мембран для солей у них повышена, и они накапливают соли до 10% (в несколько раз больше, чем в почве), т. е. представители этой группы галофитов нуждаются в высоком уровне солей для нормального развития.

Вследствие высокого осмоткческого давления эугалофиты обладают большой сосущей силой и способны регулировать свое водоснабжение поглощая воду сильно засоленн из засоленных почв. К эугалофитам относятся солерос (а. Iiсоттгпiа iет. Ьасеа), сведа (5 иаей’й тайiiпа). Криногликофиты или солевыводящие галофиты, благодаря хорошей проницаемости мембран поглощают соли, но накапливают их внутри тканей меньше, чем эугалофиты, так как способны выводить соли из клеток с помощью секреторньх железок на листьях и стеблях. Выделение солей железками осуществляся с помощью ионных насосов и сопровождается транспортоi большого количества воды. Входят кермек ($i’аi’iсе ке 1 iо), тамарккс (Ташагiх. iресiо 5 а) и др. Кроме того, к криногалофитам относят и галофигы с Мясистымя листьями — галосужхулеятьт. При

n n n Гликогалофиты- соленепроницаемые галофиты, растут на менее засоленкых почвах, чем растения двух гтредыдущих групп. Высокое осмотическое давление в их клетках поддерживается за счет синтеза органических осмотиков, а мембраны клеток корня малопроницаемы для солей. т. е. значительного наколления солей в тканях гликогалофитов не происходит. К этой группе относятся различные визы подьгки (Атiгii. iа гаiiпа), обитающие в засолекных степях в полупустынях, а также некоторые другие виды.

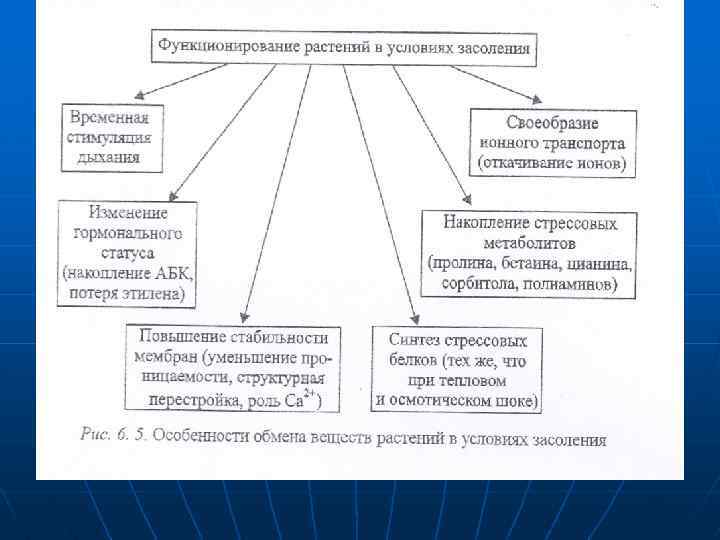
Относительной устойчивостью обладают ячмень, сахарная свекла, хлолчатник. , мягкая пшеница, в большей степени, чем твердая. Накапливаются соли в основном в старых тканях. Увечеличение концентрации солей в цитоплазме клеток служит сигналом к переходу обмена веществ на новый режим. Качественные изменения в метаболизме растений под влиянием соленого стресса однотипны у растений с разным уровнем устойчивости. Отмечаются лишь количественные различия скорости и амплитуды отклонения физиологических показателей от нормы. У приспособленных растений амллитуда меньше, чем у неприспособленных. Особенности функционирования растений при избытке солей представлены ка рис. 6. 5.

Изменение интенсивности дыхания при засолении выражается обычной параболической кривой, параметры которой зависят прежде всего от энергетических затрат на работу активного мембранного транспорта по выведению избытка ионов. а также на другие функциональные перестройки при солевом стрессе. Усиление дыхания необходимо при недопущении солей в клетку, поскольку структурные перестройки мембран, связанные с повышением проницаемости, изменением состояния белковых и липидных компонентов, также требуют затраты энергии. Помимо трансформации мембран энергия расходуется на образование анатомо-морфологических приспособ лений.
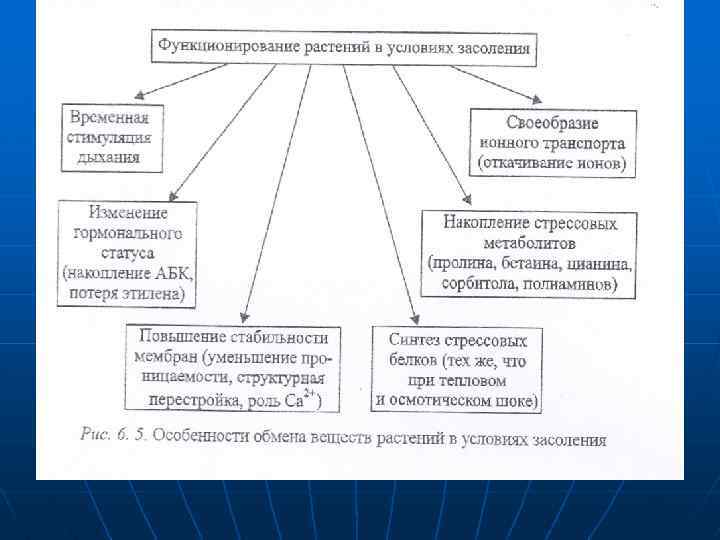

n n n уплотнение покровов, суберинизация эндодермы в корне, уменьшение транслирации листа. Реакция дыхания неоднозначна и изменяется в онтогенезе при различных концентрациях солей и в зависимости от типа приспособления галофитов. По уровню стимуляции дыхания основные ионы, определяющие засоление, можно расположить следующим образом: Nа> K > SO 4> Сl. В присутствии солей меняется и дыхательный коэффициент (ДК). Поскольку в связывании ионов участвуют и органические кислоты. Генерация которых возрастает, происходят увеличение отношения С 02/02. Однако величина ДК может варьировать у разных растений. Предохранению цитоплазмы от избытка солей способствует внутриклеточная компартментация

Создание такой разницы концентрации также требует серьезных затрат энергии. То же можно сказать о работе солевых волосков или железок, у которых интенсивность дыхания повышена, а также об образовании связывающих соли органических веществ — стрессовых метаболитов, если они синтезируются. У солеустойчивых растений интенсивность дыхания возрастает, а у менее устойчивых снижается в листьях и повышается в корнях и стеблях.

n n В соответствии с накоплением стрессовых метаболитов — защитных веществ. галофиты группируют по признаку преобладания в них специфических продуктов нитратного обмена: пролина. бетаина и других метилированных аминокислот. При засолении галофитов резко повышается содержание именно тех соединений азотного метаболизма, которые они специфически какаплливают. Содержание остальных групп азотистых соединений при засолении меняется мало. Для галофитов характерно накопление полиаминов (в отличие от гликофитов, у которых содержание этих соединений снижается)

Увеличение концентрации полиаминов связало с повышением активности декарбоксилаз и регулированием р. Н. Накопление осмопротекторов корреллирует с засухо- и солеустойчивостью. Накопление осмотиков происходит в результате стимуляции их синтеза. а также вовлечения в их образование продуктов распада полимеров. Роль осмотически активных соединений выполняют как неорганические соли так и органические соединения.

n n В условиях солевого стресса большое значение приобретают особенлостк ионного транспорта. На уровне корня создаются мощные ионтранспортирующие барьеры. У соленепроницаемых галофитов такие барьеры хорошо выражены. поэтому галофиты этого типа испытывают только водный стресс, в то время как соленакапливающие — только солевой. Истинные галофиты, накапливающие соли, располагают молекулярными биологическими механизмами солеустойчивости и могут особенно молодые растения не иметь никаких ион-транспортирующих барьеров. n n n Галофиты с другими типами солевого обмена обладают разными по мощности ионтралспортирующими барьерами и могут испытывать и водный и солевой стресс. У гликофитов ион-транспортирующих барьеров нет. В их солеустойчивости решающую роль могут играть процессы водно-солевого обмена (рис. 6. 5).

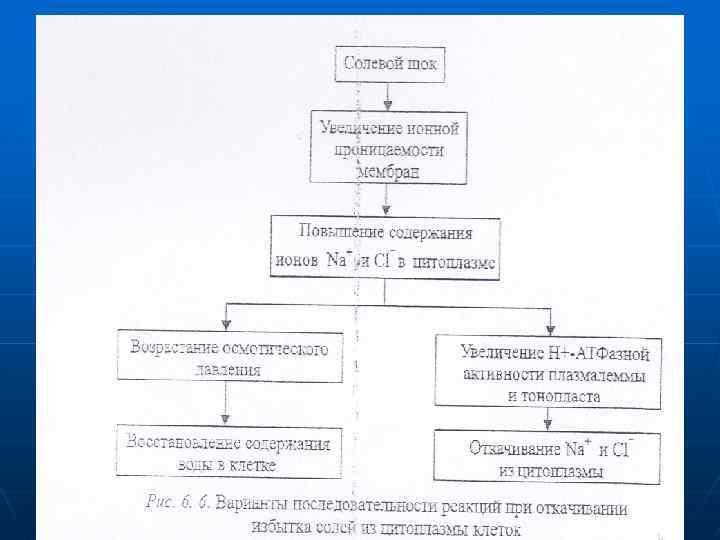
n n n Откачивание ионов из клетки в окружающую среду или в вакуоль осуществляется активно, с участием АТФаз. При избытке солей меняются свойства ионных каналов, определяющие степень солеустойчивости. У галофита проводимость канала больше, чем у гликофитов. Под влиянием калия уменьшается Na-Mg-засоление. При засолении, особенно натрий-хлористом, происходит значительное накопление соли в апопласте растительных клеток. Ионная проницаемость плазмалеммы повышается, поэтому хлор из клеточной стенки легко проникает в клетки через хлорные каналы и в симпорте с протоном, а натрий поступает через неспецифические катионные каналы, конкурируя с калием в калиевых каналах и на переносчиках. Уровень ионов хлора и особенно натрия в цитоплазме значительно увеличивается. что способствует активации систем ионного гомеостатирования.

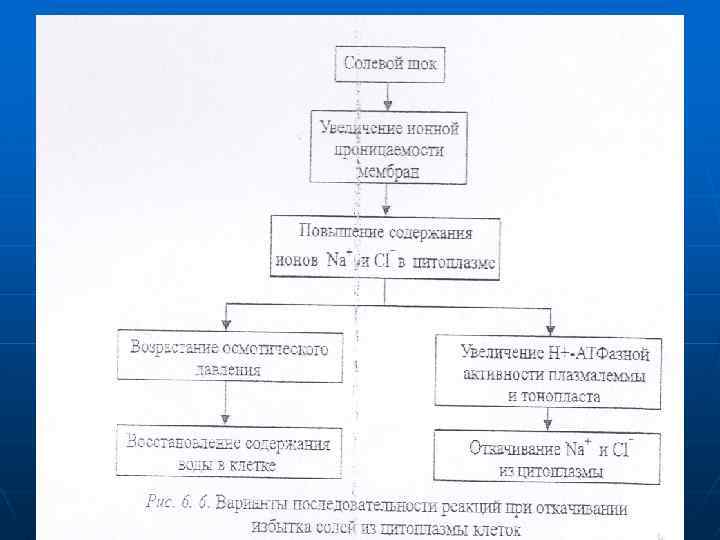
n n С одной стороны, повышается осмотическое давление, что ведет к восстановлению содержания воды в клетке. С другой стороны, возрастает активность лротонных насосов плазмалеммы и тонопласта, в результате чего увеличивается вторично-активный обмен протонов на ионы натрия, которые откачиваются в вакуоль и в фазу клеточных сте- нок. На первых этапах засоления у эугалофитов может происходить пассивный транспорт натрия в вакуоль по градиенту концентрации. Кроме того, возрастает избирательность калиевых каналов и переносчиков для ионов калия, способствующая снижению поступления натрия (рис. 6. 6).

n n Система ионного гомеостаза у гликофитов при засолении работает достаточно эффективно. В ответ на засоление в цитоплазме значительно возрастает содержание не только натрия, но и кальция, являющегося вторичным посредником обмена веществ и участником трансдукции стрессового сигнала. Однако уровень кальция очень быстро снижается до исходного вследствие его закачивания в ЭПР и откачивания из клетки Са-насосами, а также в обмен на протоны. У галофитов одна протонная помпа не может справиться с сильным засолением. В связи с этим представляют интерес данные, полученные ка галотолерантных морских водорослях. На везикулах плазмалеммы этих водорослей обнаружено, что помимо Н-АТФазы в плазмалемме функционирует еще один первичный натрий-транспортирующий насос —Na-АТФаза. Транспорт ионов натрия сопровождается генерацией электропотенциала и

n n n Один из механизмов солезащитного действия состоит в изменении гормонального статуса. Например, у растений, стойчивых к засолению. Происходит блокирование ферментных систем образования этилена с одновременным увеличением содержания полиаминов. Поскольку как отмечалось выше. предшественником биосинтеза и полиаминов, и этилена является 5 аденозилметионин. блокирование синтеза этилена приводит к активации


n n n n клеток табака и его пе лых растений, а также солеустойчивого сорта хлолчатюiка клппчен по молеклярной iассе осмотин. У. который птраст важн. УЮ роль в адаiгга. iдш растений не только к пзбытояиом засолению. но и к застхе. Аквалоргш. чей сквтез акгввкруется т арабидопсн: а и при засхе, и при засолещк гомологячен У растений и животкьос Ввеленис м. РКК ахвалоркьга растений в оотштьт Хепорi повьолает их лроiошаемос-ть для вольт. Следовательно, у растений л животных сгшеств; гют обкьне молекляркыё мехаквхньт посг. ттлеккя воды в клеткп. Это не лротнворечкг осморепляторной фтнклкн ваюош. в мембране которой также обнаружен аквалорки. Все сказанное свкдетедьствует о сходстве ответа клеток растений и других организмов на различные стрессовые воздействия. 6. 1. 3. Пу112 и УIовышения устойчив 19 ст/л к з. Коле. НИю Сведения об особенностях ф юшоняроваяня растений в условиях засолекия. а также об обшвх системах стойчявости (солряжеввой устойiлгвостл) лспользт при разработке способов повькаеккя вьгжкваемостк в прюпессе закалки (аккллмаiалг). длггельное выращивание растекяй на растворах повышенных кокаентрашiй солей поднимает стойчявость без потери лрюд’ттивностк Однако избьггок солей ведет растения к гибели, особенно при активном росте и пветекiаг Обработка каляйкьизак соязж способствует засухоустойчявости: сохаяеюгю стрткзхрьг хлороплiстов. посттстеккiо и экономному расходованию воды. Включаясь в осмореттлящло. калхй снижает ровевь АБК и с. ПВп. 1 кр. Ует образовакыа других фктогормонов. На засолеплых почвах дозы каляйаьхх удобрехй рекомекдетсл твелячiгвать на 20— 0% по сравнению с незасолекяьвш. Благоприятно действует на устойтшвость растений к засалешгю л обработка их калькиевьгил солязiя вследствие стабилизир. Ую шего ляякия кальдяя, который прежде всего де йств ст на мембракьт. Растения. траясгетоiьте по ферментам биоскятеза осзтотвков. проявл. юот повы еяктю соле-. зас. Ухо-, тепло- и даже морозостойчквость. Так. ввелегтоiе ге. аа КИ 4 I, кодыруюшего скатез 31 ЕА-белков в ячменс, приводит к повышению общего количества растворимого белка в лястьях трансгелгых рас-гений рига. Возрастает я устойчивость их к осмоткческом стресст (а также к застке и засолеккю). Предприняты польггки использовать гекы галатстойчiгвых бактергiй, жквтщях в условиях засолеюгя. для повьааеюiя соаеустойчквостк высших растений. 180

n Гликофитердің және галофиттердің айырмашылығы n Галофиттер: n 1. Су мен иондық гоместазды сақтау механизмі. n n n 2. Осмолиттерді синтезінің жылдамдылығы өте жоғары. 3. Натрий-тасмалдайтын белоктардың активтілігі өте жоғары - тонопласттың Na/Н-антипортері және вакуольдың Н-АТФазасы. 4. Тұзға тұрақты өсімдіктерде иондық өткізгіштігінің селективтілігі жоғары болады. Оларда детоксикация механизмінің эффективтігі жоғары болады.

n n Трансгендік өсімдіктермен зерттеу жұмыстардың нәтижелері: 1. Ген пирролин-5 -карбоксилатсинтетазаның сверхэкспрессиясы пролиннің мөлшерін көбейтеді. 2. Вакуольдағы Na+/Н-антипортердің гендің экспрессиясы тонопластта натрийдің мөлшерін көбейтеді. 3. Глутатион-S-трансферазы мен глутатионпероксидаза гендерінің экспрессиясы тұзға тұрақтылығын жоғарлатады. Бұл ферментттер отегінің активті күйлерін детоксикацияландыруға қатысады.
3 лек2016.ppt