2 Лек.2016.ppt
- Количество слайдов: 107



Лекция 2 Су тапшылықтағы өсімдіктердің бейімделу механизмдері n n n n 1. Сутапшылық стресіне сипаттама 2. 3. 4. 5. Сусыздықтан қашу стратегиялар. Су потенциалы Осмолиттер Полиаминдер Шаперондар, убиквитенедер, протеазалар.

n n Егер транспирация жылдамдығы тамыр жүйесінің суды сіңіру жылдамдығынан жоғары болған жағдайда өсімдік су тапшылығына ұшырайды. Бұндай жағдай құрғақшылық кезінде ғана емес, топырақтың тұздануы кезінде де және өте төмен температура жағдайында да байқалады.

n Соңғы екі жағдайда дегидратация табиғаты “сусыз” стресс компоненті болып табылады және ол өсімдікте көбінесе топырақтағы судың мөлшері шектелмеген жағдайда дамиды. Бірақ өсімдік қабылдай алмайды. Мысалы, топырақтың тұздануы жағдайында топырақтың осмостық қысымының жоғары болуына байланысты тамыр жүйесі суды сіңірмейді (су потенциалы төмен).
 - бөлiктерiнiң бiр орынынан баск, а орындарға үздiксiз")
2. Судың осмостық ciңyi. ІРІГУ (ДИФФУЗИЯ) - бөлiктерiнiң бiр орынынан баск, а орындарға үздiксiз қозғалып араласуы. ОСМОС - ерiткiштің өзiнен жартылай өткiзгiш мембрана арқылы бөлiнген epiтiндiгe өтуі. Осы құбылысты бiрiншi рет француз физиологы Г. Дютроше 1826 ж. жасанды осмометр арқылы зерттедi. В. Пфеффер - осмометрдiң жетiлiңкiреген түрін ұсынды. Жартылай өткiзгiш мембрана арқылы судың ерсiлi-қарсылы жылжуы да теңеседi. Бул жағайдағы гuдростатuқ. алық қысым Р потенцuалдық осмос қысымына тең болады.

n n Кез келген организмнің тiршiлiк әрекеттерi қалыптагыдай жузеге асуы ушiн сыртқы ортадан жеткiлiктi мелшерде судың eнyi қажет. Өciмдiк клеткасына судың eнyiнe биоколлоидтардың бөрту, гидраттану куштерi әсер етедi. Ол қасиет тұқымдарды суға малғанда байқалады. Бiрақ тipi клеткаға судың eнyi негiзiнен осмостық кушке байланысты болады. Жартылай еткiзгiш мембрана арқылы судың ерсiлiқарсылы жылжуы да теңеседi. Бул жағайдағы гuдростатuкалық қысым Р потенцuалдық осмос қысымына тең болады. n Р=π* Ерітінділерде осмостық қысым тұрақты температурада еріген заттың концентрациясымен /көлемінің бірлігіндегі санымен/ белгіленеді.

S= π Р S cу сорғыш күші π осмостық қысымы Р тургор қысымы S=0 , π =Р π – осмос қысымы Р – тургор қысымы

n n Ең жоғары судың потенциалы – химиялы таза судың потенциалы. Ол 0 –ге тең Сондықтан әр бір ерітіндінің және биологиялық сұйықтықтардың су потенциалы теріс мәнінде. Су потенциалына осмостық потенциал ψπ және гидростатикалық қысымымен байланысты потенциал ψр кіреді

Өсімдік клетка – осмостық жүйе Клеткаға кіретін судың күшін сорғыш күш S. деп атайды n Ол клетканың су потенциалына тең (ψН 2 О). n Су сорғыш күші клетка сөлінің осмостық қысымымен (π*) және клетканың тургорлық (гидростатикалық) қысымымен (Р) белгіленедіі. n Ол клетка қабығының электростатикалық созылғандағы қарсы күшіне тең: n S = π*- Р

n n Осы белгілерді термодинамикалық ауқымдарына /мөлшерлеріне/ ауыстырғанда бұл теңдіктің түрі мынадай болады: - (ψН 2 О)= - ψπ - ψр n Клетка суға қанығып тұрғанда (тургесцентті) оның сорғыш күші 0 -ге тең, ал тургорлық қысым осмостық потенциалдық қысымына тең n n S=0; Р= π

n n Клеткада толық тургор топырақта және ауада су жеткілікті болғанад байқалады. Клеткада су азайғанда (жел тұрғанада, топырақта су азайғанда), бірінші клетка қабығы су тапшылыққа ұшырайды, оның су потенциалы вакуольдың потенциалынан төмен болады, су клетка қабығына жылжиды n n Вакуольдан су шыққыанда, клетканың тургор қысымы төмендейді, сорғыш күші ұлғаяды. Ұзақ су тапшылықта өсімдіктер солып қалады. Осындай жағдайда Р=0, S = π*
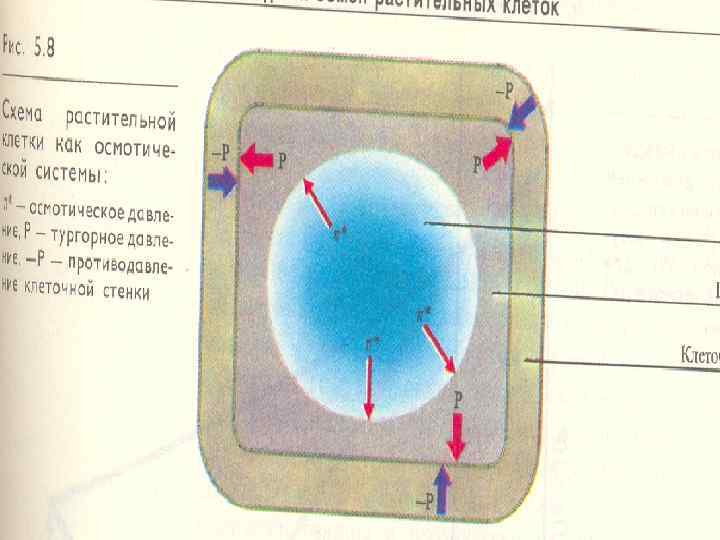

n Тургорды жоғалтқан клетканы экспериментте көруге болады. Клетканы нипертоникалық ерітіндіге салғанда плазмолиз деген құбылыс байқалады. n n n Жас ұлпаларда су жоғалтқанда тургор қысымы теріс мәнінде болып, протопластардың көлемы азайып, клетка қабығынан айырылмай бүріседі. Ол құбылыс циторриз деп аталады. Тамыр клеткалардың вакуоль сөлінің концентрациясы - 0. 31, 2 МПа. Жер үсті мүшелердің клеткаларында вакуоль сөлінің концентрациясы - 1, 0 -2, 6 МПа. Ол осмостық концентрациясының және сорғыш күштің вертикальды градиентін қамтамасыз етеді. . Галофит клеткаларында ең жоғары осмостық қысым: 15 МПа.

n n n Өсімдіктегі су жылжудың механизмдері. Су алмасу 3 сатыдан тұрады: Тамырмен су сору. Сосудтар арқылы судың жылжуы Транспирация, жапырақтар арқылы су булануы. Осы процестер бірімен байланысты.

n n n Өсімдік клеткасындағы су потенциалының азаюы сусыздан қашу стратегиясының көрінісі. Су потенциалы градиенті бойынша су пассивті түрде су потенциалы жоғары аймақтан төмен аймаққа қарай қозғалады ψ. Егер тамыр клеткасының су потенциалы ψкл топырақтың су потенциалынан жоғары болса, онда өсімдік топырақтан суды сіңіре алмайды. Су топырақтан тамыр клеткаларына қарай қозғалу үшін ψорт қарағанда ψкл өте төмен деңгейге дейін жету керек. Кейбір өсімдіктер су тапшылығына өте сезімтал болады және ψкл өте төмен болғанда, тез солып қалады(тургордан айырылу).

n Р — тургорлық қысым; πкл — осмостық қысым ; ψ — су потенциалы n ψ кл > ψ орта Су клеткаға кірмейді ψ кл < ψ орта Су клеткаға кіреді

n n Клетка цитоплазмасында су және тұз стресстерінде осмостық қысымның реттелуі төмен молекулалы органикалық заттардың биосинтезі есебінен жүзеге асады және оларды осмолиттер деп атайды. Ал вакуольде осмостық қысымның реттелуінде бейорганикалық иондардың жинақталуы негізгі рольді атқарады.

n n n Осмолиттер. Функциясы—осмореттеу процестеріне қатысу. Көптеген бейорганикалық иондар, мысалы, Na+ және Сl иондарының жоғары концентрациясы улы, сондықтан өсімдік клеткасында цитоплазманың осмостық қысымын реттеуге пайдалану мүмкін емес. Биополимерлермен үйлескен осмолиттер цитоплазмада грамға шаққанда бірнеше жүз микромольге дейін жинақтала алады және ол ешқандай улы әсер етпейді. Осыған байланысты өсімдіктер цитоплазманың осмостық қысымын реттеу үшін клеткадағы бейорганикалық иондарды емес осмолиттерді пайдаланады.

n n Осмолиттер - химиялық табиғаты бойынша алуан түрлі төмен молекулалы органикалық қосылыстар. Олар суда жақсы ериді, улы емес және бейорганикалық иондармен салыстырғанда метоболизмде өзгерістер туғызбайды. Сондықтан да олардың екінші аты үйлесетін заттар деп аталады (compatible solutes). Үйлесетін заттардың р. Н-тың физиологиялық мәнінде бейтарап болу тенденциясы бар.

n Олар цитоплазмада диссоцияланбаған формада немесе цвиттерион түрінде, яғни теріс және оң зарядтарды алып жүретін кеңістік бойынша бөлінген молекулалар түрінде болады. n n n Кейбір осмолиттер амфифильді қосылыстар болып табылады. Амфифильді заттар молекулаларында полярлы емес (гидрофобты), сонымен бірге полярлы (гидрофильді) топтары болады


n n Осмолиттердің қос функционалдық ролін ескере оларды көбінесе осмопротекторлар деп атайды. Мысалы, өсімдік жапырағы препаратында жүргізілген тәжірибеде глицин-бетаин рибулозобисфосфаткарбоксилазалардың инактивтелуін және фотожүйе II комплексінде бөлінетін О 2 дестабилизациялануын жояды. Осмолиттер биополимердің гидратты қабықтарын бұзбайды деп есептеледі. .

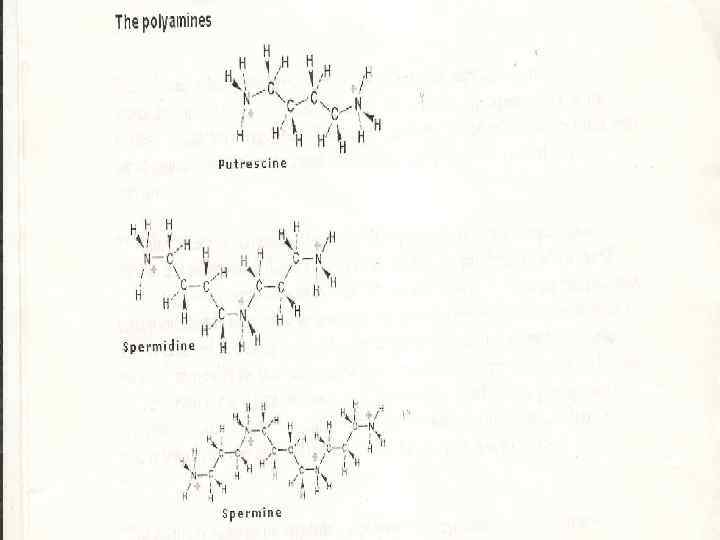
n n n Полиаминдер Стрессор әсерінен синтезделетін кейбір төмен молекулалы органикалық қосылыстар мардымсыз мөлшерде синтезделеді және ол осмореттеу қызметін атқара алмайды. Алайда олар стресс жағдайында биополимерлер протекторы ретінде маңызды роль атқарады. Ондай қосылыстарға диамин путресцин және одан түзілетін полиамин спермидин және спермин жатады.

ПА-дер өсімдіктерде көп таралған. n Олар ДНК, РНК және белоктардың синтезін қарқындандырады. n Поликатион ретінде, полиаминдердің теріс зарядты биомолекулалармен әрекеттесетін қабілеті жоғары n әсіресе, ДНК, РНК, фосфолипидтер және қышқыл белоктармен, мембрана компоненттерінің және клетка қабығының анион топтарымен әрекеттесетін қабілеті жоғары n

n n n Дегидратация нәтижесінде түзелген белоктар. Әр түрлі қызметтер атқаратын белоктар жауап реакция ретінде de novo синтезделенеді. Lea–белоктар. Кейбір су тапшылықта пайда болатын белоктар цитоплазмалық биополимерлерді және клеткалық құрылымдарды дегидратациядан зақымданудан қорғап тұрады. Оларға Lеа (1 аte еmbryogenesis abundant) белоктар жатады. Олар пісіп және қурап жатырған дәндердің ішінде экспрессияланатын LЕА гендердің әсерінен пайда болғанда ашылған. Соңынан кейбір Lеа белоктар су тапшылық жағдайда, тұзды ортада және төмен температураларда вегетативтік мүшелерде табылған.
, -")
n n n Шаперондар мен протеазалардың ингибиторлары. Шаперондар - ширатылып–бұрылып бүктеліп, оралып (folding), - жатқан полипептидтермен байланысатын белоктар, Полипептидтермен байланысып, шаперондар белоктардың қателіксіз агрегацияланбай ширатылуын қамтамасыз етеді. Кейбір шаперондар «жөндеу станциялар» рөлін атқарады, олар белоктардың қате ширатылғанын жөндейді. Шаперондардың ең басты функциясы – мембраналардан өткенде белоктарды ширатылдандыру және өз ұзындап орналастыру (unfolding), және белоктарды жиналту мен таралту. Полипептидтік тізбек мембранадан өткенде ұзын, ашық күйінде өтелады.

n n n Протеазалар мен убиквитиндер Су тапшылық жағдайда протекторлық әрекеттерері бар қосындылар және шаперондар барына қарамастан кейбір клеткалық белоктар денатурацияға ұшырайды. Денатурацияланған белоктар гидролиздену керек. Ол функцияны протеазалар және убиквитиндер атқарады, олардың гендеры стресс жағдайларында экспрессияланады. n n Убиквитиндер – молекулалық салмағы төмен (8, 5 к. Да) белоктар. Олар денатурацияланған белоктың N ұшына –байланысып, протеазалардың жұмысын қамтамасыз етеді.

n n n Аквапориндер Су мембранадан каналдар арқылы өтеді. Ол каналдарды аквапорин белоктары құрады. n n n n Каналдардың өткізгіші және саны өзгергеніне қарай судың трансмембрандық жылжуы реттеліп тұрады. Осмолиттер көбейгеннен клетканың суды соруы су каналдар арқылы жүзеге асырылады. Су каналдарының активтілігі мембраналардың су өткізгіштік қабілетін реттеп түрады. Осындау реттелу механизмнің бірі аквапориндердің фосфорлану және дефосфорлану. Фосфорланғанда су каналдары активтеленеді, дефосфорланғанада – активтігі төмендейді.

n n n Стресс жағдайлар өсімдіктредің судың статусын өзгертеді Сумен өсімдікке минералдық заттар тасмалданады. Леп тесіктерді ашық күйінде ұстап тұру үшін су жеткілікті деңгейінде болуы керек

n n Температура 10 - 15°С жоғарылағанда 35 -40°С су тапшылыққа ұшырайды.
 - более")
n n ЗАСУХОУСТОЙЧИВОСТЬ Территории засушливых регионов (годовая смма осадков ниже 250 мм) - более трети земной поверхности. Половина этой площади (около 12% суши) относятся к крайне аридным территориям. Многие сельскохозяйственные районы расположены в аридных зонах. Ежегодные потери урожая от засухи огромны, так как даже в умеренной зоне за вегетативный период растения испаряют воды больше, чем выпадает атмосферных осадков.

n n n Засуха, т. е. явления, при которых расход воды превышает ее поступление, бывают почвенные и атмосферные. Чаще всего засуха начинается с атмосферной, характеризующейся низкой относительной влажностью воздуха, чему сопутствуют повышенная температура и высокая инсоляция. При длительном отсутствии дождей к атмосферной засухе добавляется почвенная, возникающая в результате
")
n n n При сильном ветре происходит высыхание тканей без пожелтения ( «захват» ) или появляются ожоги в виде бурых пятен ( «запал» ) Глинистые минералы и гумусовые вещества почвы – коллоиды и могут удерживать значительное количество влаги. Вода входит в состав многих химических комплексов - «связанная» вода. Она недоступна для растений. Растениям доступна вода почвенного раствора. , содержащаяся в капиллярах почвы, «капиллярная» вода.

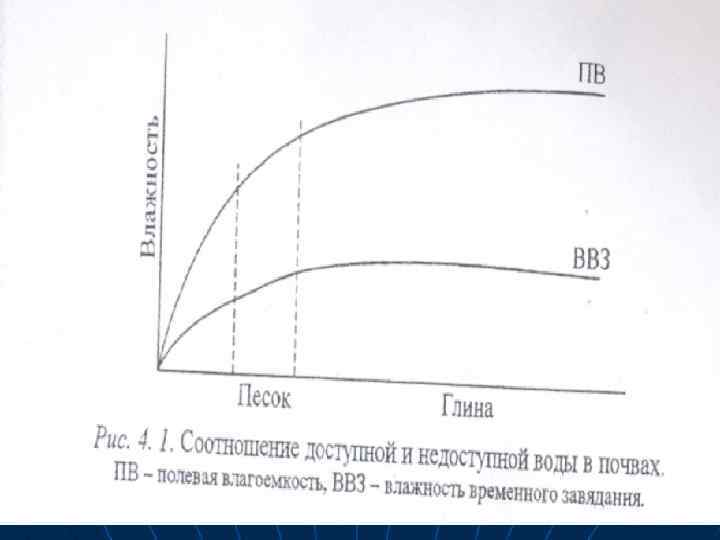
n n n n Количество воды, которое почва способна удержать вследствие капиллярных взаимодействий и оводнения коллоидных частиц, называется «полевой влагоемкостью» . Она характеризует максимальное количество почвенной влаги. После атмосфервых осадков в почве бывает гравитационная вода. Она тоже потребляется растениями, но довольно быстро ухолит в нижележащие почвенные горизонты. Почва среднего механического состава (супесчаная и суглинистая) имеет максимальную полевую влагоемкость. В глинистых почвах больше связанной воды, а в

n n n n Дефицит влаги наблюдается в жаркое подуденное время. При этом увеличивается сосущая сила листьев, что активизирует поступление воды из почвы и раскрывание устьиц. Тургор листьев, потерянный днем, восстанавливается вечером и ночью. Такое состояние растений называется временным завяданием, а влажность почвы, приводящая к этому — влажностью временного завядания (ВВЗ). Когда запасы доступной воды в почве исчерпываются, развивается глубокое завядание, или стойкое завядание растений. в этом случае тургесцентность тканей утром не восстанавливается. Судың топырақтан тамыр арқылы ciңyi Өciмдiк суды өте көп мелшерде пайдаланатын дықтан, оньщ уздiксiз eнуін қажет етедi. Өсiмдiктiң бұл қажетін қамтамасыз eтетін негiзгi мушесi тамыр жуйесi болып есептеледi. Астык. тук. ымдастардьщ тамыр жуйесi 1, 5 2 м тереңдеп және жан жагына тарамдалып өceдi. Өсімдіктерде су алмасу процестер і кезеңнен тұрады; 1. Тамыр ақылы сіңуі. 2. Түтіктер арқылы жылжуы. 3. Танспирация арқылы судың жапырақ арқылы булануы. Су құрғақ топыраққа тигенде тез сіңіліп кетеді. Соңынан судің сіңуінің жылдамдығы азаяды. Судің сіңуінің жылдамдығы ең аз болғанада оны топырақтың далалық ылғал сиымдылығы деп атайды. Ол топырақтағы жалпы ылғал қорының ең жоғарғы деңгейін сипаттайды. Тұрақты солуының ылғалы деп өсімдіктің солу көрсеткіші ылғалдылығын айтады. Ол кезде топырақа су қуйғанша өсімдіктер солып тұрады. Бұл топырақтың ылғал қорының ең төменгі деңгейін сипаттайды. В песке содержание доступной вольт н влагоемкость ниже, чем в глине (рис. 4. 1).

n n Влажность почвы в этих условиях — это влажность стойкого завядания (4 О% от полной влагоемкости). Разность между полевой влагоемкостью и влажностью временного завядания отражает запас доступной воды в почве.

n n n Большинство активно вегетирующих растений завядают при водном дефиците — 1. 5 МПа, а гибнет при — 15 МПа (ВВЗ — 2, 5 — 3. 5 МПа). Пыльца растений выносит водный дефицит до — 100, семена до — 150 МПа. При небольшом дефиците воды (5— 10%) интенсивность метаболизма у большинства растений достаточно велика. В этих условиях транспирация высока при максимально открытых устьицах. Эффективно осуществляют фотосинтез и

n Снижение содержания воды на 20— 30% приводят к развитию водного дефицита в тканях, который возникает задолго до того, как содержание влаги в почве достигнет уровня устойчивого завядания. n n n Реакции растений на засуху При недостатке влаги в растениях происходят значительные изменения обмена веществ. Содержание свободной воды в клетках уменьшается. Возрастает концентрация клеточного сока и цитозоля. Гидратные оболочки белков и других полимеров утончаются и утрачиваются.

n n Потеря гидратных оболочек у белков приводят к нарушению их третичной и четвертичной структурыи, в конечном счете, к денатурации. Активность ферментов снижается, а затем они инактивируются

n n Проницаемость мембран возрастает. Вследствие дегидратации мембраны утрачивают бислойные структуры и в них обнаруживаются значительные конформационные изменения (образоваше гелевой фазы). Меняется и качественный состав мембран, поскольку инактивируются десатуразы, что приводят к уменьшению количества ненасыщенных жирных кислот (линолевой, линоленовой) в составе липидных компонентов мембран. В результате индекс ненасыщенности

n n n Текучесть мембран уменьшается, и ингибируется их функциональная активность. При увеличении проницаемости плазмалеммы она теряет кальций, при этом улучшается ее проводимость для воды. Одновременно с транспортом воды изменяется активности ц-АМФ и Са 2 зависимой протеинкиназы, которые в качестве мессенджерных систем участвуют систем в регуляции фосфорилирования белков мембран, которое возможно в присутствии определенного количества Са.

n n Гидролиз различных органических вешеств начинает преобладать над синтезом. Протеолиз - накопление низкомолекулярных белков усиливается гидролиз полисахаридов, увеличивается концентрация растворимых углеводов. - Осмотически активные вещества, и увеличение их содержания способствует повышение концентрации вакуолярного клеточного сока и росту осмотического давления, а значит, и большему поступлению воды. - С другой стороны, состав клеток меняется, значительная часть ионов выходит из клеток, что ведет к подавлению синтеза и

n n Гидролитические процессы активируются в первую очередь в старых листьях нижних ярусов. Интенсивный отток продуктов гидролиза в верхние, более молодые листья и включение их в обмен веществ позволяют растению более экономно расходовать питательные субстраты, а опадение листьев уменьшает транспиририрующую поверхность.

n n Интенсивный распад полирибосом, снижается интенсивность синтеза РНК, и уменьшается ее количество. Активируются рибонуклеазы. Изменение же ДНК наблюдается только при сильной и длительной засухе. Происходит задержка роста вследствие торможения и клеточного деления и растяжения, для которого особенно необходима вода. В результате клетки становятся более мелкими, чем обычно. Наряду с ингибированием роста листьев и стеблей рост корней, особенно засухоустойчивых растений, может даже активироваться для

n n Изменения интенсивности дыхания в зависимости от содержания волы в тканях графически в большинстве случаев могут бьпь представлены колообразной кривой, отражающей их двухфазный характер. При небольшой потере воды наблюдается возрастание интенсивности дыхания, а при глубоком дефиците — понижение. Первоначальное повышение интенсивности дыхания связано с увеличением энергетических затрат на синтез осмотиков, защищающих белки при обезвоживании. При более продолжительном воздействии происходит спад интенснвности дыхания и снижается его энергетическая эффективность в результате повреждения дыхательных систем

n n n Интенсивность фотосинтеза снижается раньше, чем интенсивность дыхания. Ингибируются как фотохимические реакции, так и реакция восстановления СО 2. Ограничение работы фотосинтетического аппарата обусловлено закрыванием устьиц и уменьшением поступления углекислого газа, а также нарушением структуры хлоропластов н синтеза хлорофилла, разобщением транспорта электронов и фосфорилирования, что ведет к дефициту АТФ в клетке. В совокупности с ингибированием ЭТЦ дыхания это способствует торможению синтетических процессов у растения в целом. К тому же при засухе накапливаются токсические продукты обмена, в том числе интермедиааты ПОЛ, особенно в связи с инактивацией фотосинтетического и окислительного

n n Неблагоприятное действие засуха состоит в первую очередь в обезвоживании и нарушения метаболических процессов. Такие реакции растений развиваются в ответ на недостаточное снабжение водой или комплексное действие водного дефицита, интенсивного света и перегрева. Кроме того, надо иметь в виду, что в онтогенезе существуют наиболее чувствительные периоды к высокой температуре и дефициту влаги. Эти периоды совпадают с наиболее интенсивным ростом растений и

n n . Группы растений, способных переносить засуху По способности переносить условия засухк различают растения гомойогидридрические, т е. способные активно регулировать свой водный обмен, и пойкилогидрические, водный обмен которых определяется содержанием воды в окружающей среде. При засухе они пассивно теряют воду. Гомойогидрическими являются большинство высших растений. Они обладают тонкими механизмами регуляции устьичной и кутикулярной транспирации, а также мощной корневой системой, обеспечивающей

n n n Поэтому даже при значительных изменениях влажности не наблюдается резких колебаний содержания воды в клетках. Различают ксерофиты — растения сухих местообитаний, мезофиты — обитатели мест с умеренным содержанием воды, и гигрофиты — наземные растения, находящиеся обычно в условиях избыгочной влажности. Гигрофитам близки гелофиты — растения болот, берегов водоемов, и гидрофиты, обитающие в водной среде. Ксерофиты, в свою очередь, по механизму приспособления к засухе подразделяются на следующие группы (рис 4. 2). Первая — суккуленты, или псевдоксерофиты (кактус, алоэ, молочай), запасающие влагу в листьях,


n n Транспирация, фотосинтез, рост осуществляются медленно. Разветвленная корневая система ходнт в почву неглубоко. Траты воды сведены к минимуму, и обезвоживание клетки переносят плохо. Эти растения способны использовать даже скудную влагу атмосферных осадков. К ним относятся растения с САМ-типом фотосинтеза, которые эффективно используют воду. У таких растений мало устьиц, днем они обычно закрыты, что предотвращает потерю воды, а открываются ночью. Тогда СО 2 поступает в листъя и в результате карбоксилирования фосфоенолпирувата сохраняется в виде оксалоацетата, который, восстанавливаясь до малата, накапливается в вакуолях клеток листа.

n n n Использование внутренних резервов СО 2 происходит днем при закрытых устьицах. Малат из вакуоли поступает в цитоплазму, декарбокилируется НАДФзависимым малик-энзимом, в результате чего образуются пируват и СО 2. Углекислота поступает в хлоропласты и включается в цикл Кальвина

n n n Вторая — несуккуулентные ксерофиты, которые, в свою очередь, по уровню транспирации делятся на четыре группы: 1) эуксерофиты, или настоящие ксерофиты (полынь), жароустойчивы, с небольшими, отдельными листьями, транспирация невысока, могут выносить сильное обезвоживание, осмотическое давление велико; корневая система разветвлена, находится на небольшой глубине; хотя для них характерно экономное расходование воды благодаря регуляции работы устьиц они переносят как обезвоживание. так и перегрев: тем не менее при обилии воды в почве рост этих растений интенсифицируются 2) гемиксерофиты, или полуксерофиты (шалфей), отличаются очень интенсивной транспирацией благодаря деятельности глубокой корневой системы доходящей до грунтовых вод; транспирация способствует охлаждению; число устьиц велико, хорошо развита система проводящих элементов, вязкость цитоплазмы низкая, полное обезвоживание
 стипаксерофиты (степные злаки — ковыль) быстро используют влагу кратковременных дождей, хорошо")
n n 3) стипаксерофиты (степные злаки — ковыль) быстро используют влагу кратковременных дождей, хорошо выносят перегрев, но лишь недолгое обезвоживание; 4) пойкилоксерофиты, в отличие от гомойогидрофитов, не способны регулировать водный режим, поэтому теряют воду, вплоть до перехода в воздушно-сухое состояние, и при засухе впадают в анабиоз; к этой группе относятся лпаа’шкки, мхи, ряд цветковых растений; водный дефицит вызывает у них постепенное снижение интенсивности обмена веществ; клетки пойкилоксерофитов, отдавая воду, постепенно уменьшаются в объеме.

n n n Третья — эфемеры, растения, которые “уходят” от засухи: их короткий вегетационный период совпадает с периодом покоя. Мезофиты также могут приспосабливаться к временной засухе, при этом у них выявляются признаки ксерофитов. Например, верхние листья, которые

n n n -У них большое количество мелких устьиц на поверхности, которые дольше остаются открытыми; -сеть проводящих пучков гуще, палисадная паренхима развита лучше. Явление ксероморфизации верхних листьев известно как закон В. Р. Заденского (1904)

Благодаря открытым устьицам фотосинтез поддерживается на высоком уровне, образуется значительное количество ассимилятов, что способствует увеличению концентрации клеточного сока и оттягиванию воды от ниже расположенных листьев. Возникновение ксероморфной структуры наряду с редукцией листьев, опушенностью, толстой кутиклой и т. п. служит анатомическим приспособлением к недостатку воды.

n n n Механизмы приспособления растений к засухе Существование растений при водном дефиците зависит от доступности воды и от эффективности ее использования. Лучше переносят эасуху растения, обеспечивающие себя водой или использующие ее наиболее продуктивно. Механизмы устойчивости к водному дефициту обычно делят на два способы избежания стресса – сохранения нормальной оводненности растительных тканей, механизмы толерантности, позволяющие существовать при дегидратации - рис. 4. 3).
 глубокое проникновение корней вплоть до грунтовых")
n n n Для этих растений характерно: 1) глубокое проникновение корней вплоть до грунтовых вод и больщая скорость их роста (лжексерофиты); 2) хорошая устьичная регуляция (суккуленты); 3) меньшее количество устьиц, погруженные устьица (суккуленты, ксерофиты): 4) определенная ориентация листьев и их скручивание (гемиксерофиты, стипаксерофиты);
 уменьшение размеров листьев, их опушенность, многослойный эпителий и палисадная паренхима")
n n n 5) уменьшение размеров листьев, их опушенность, многослойный эпителий и палисадная паренхима (мезофиты, ксерофиты); 6) толстая кутикула для снижения кутикулярной транспирации, которая может составлять от 5% до 25% и даже 50% от полной транпирации у приспособленных видов (ксерофиты) 7) большое число крупных сосудов ксилемы (гемиксерофиты): 8) высокое содержание осмотически активных веществ (ксерофиты); 9) опадение листъев, поскольку листопадность деревьев возниклакак приспособление к

Стрестен қашу әдістері Судың қалыпты мөлшерінде сақталуы Суды экономды жұмсаған және тапқан жағдайы арқылы судың потенциалының жоғарылауы Тұрақтылықтың механизмдері Метаболизмнің өзгеруі Эфемерлер Эуксерофиттер Снижение ин. Зат алмасу процестердің баялауы Пойкилоксерофитте р
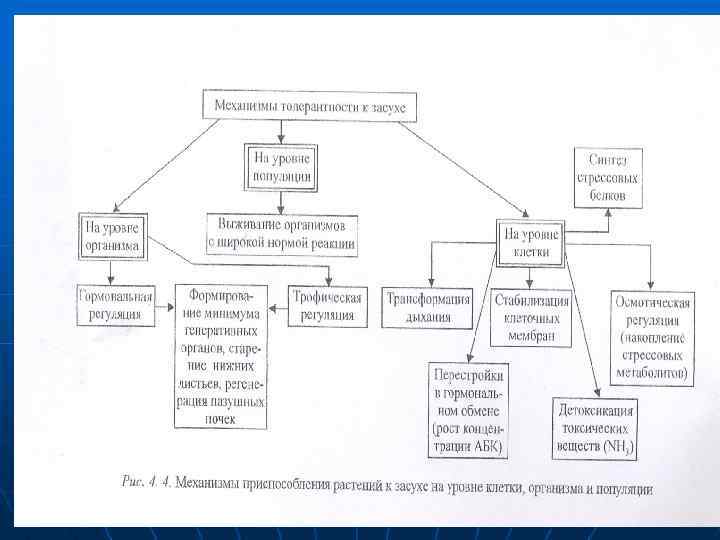

n n Нередки также случаи «ухода» от периода засухи. Например, прохождение вегетации до наступления сильной засухи (эфемеры). Механизмы толерантности (рис. 4. 4) обеспечивают растениям прирост при низком водном потенциале в результате снижения интенсивности обмена веществ (пойкилоксерофиты) или метаболических перестроек (эуксерофиты). Во многом эти механизмы те же, что и для избежания обезвоживания. Особое значение они имеют в определенных зонах при экстремальных условиях, когда достигают запасов влаги в почве нет, а водоудерживающая способность оказывается нелостаточной, чтобы защитить растения от обезвоживания и низкого водного потенциала (настоящие ксерофиты — эуксерофиты, пойкилоксерофиты, впадаюшщие в анабиоз).

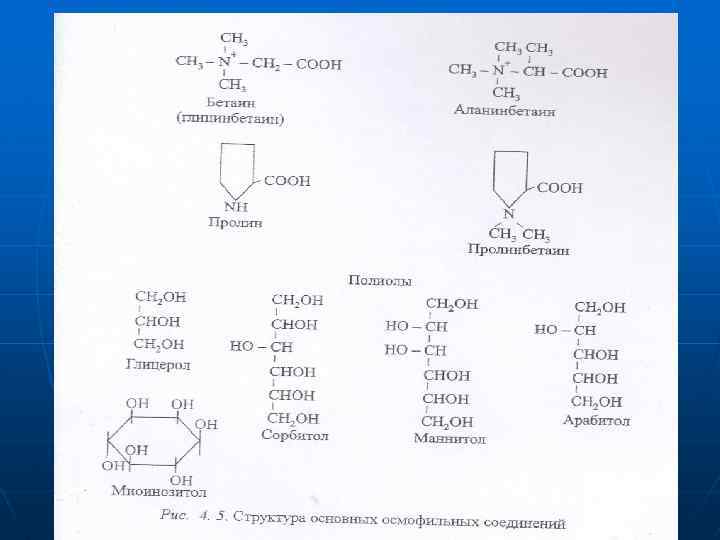
При этом у мезофитов и ксерофитов высокий водный потенциал может сочетаться как с большой эффективностью использования воды (низкий коэффициент транспирации у эуксерофитов), так и со значительной потребностью в воде (высокий транспирационный коэффициент у геми- и стипаксерофитов). Механизмы толерантности определяют на уровне клетки, организма и популяции. На уровне клетки - осмотическая регуляция (рис. 4. 4). Осмотикн должны быть нетоксичными, обладать очень высокой растворимостью, по возможности быть биполярными, чтобы заменять диполи воды и не участвовать в основных метаболических процессах (именно поэтому сахара — плохие осмотики).

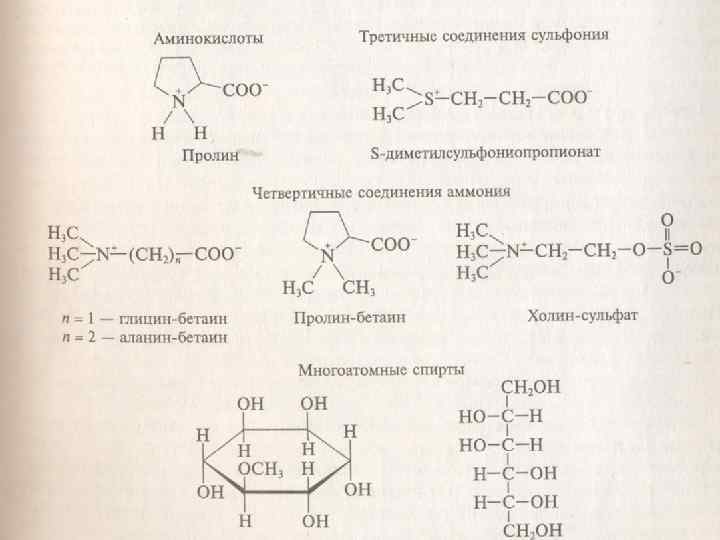
n n Роль осморегулирующих веществ играют личные моно- и олигосахариды, аминокислоты, в первую очередь пролин, производные амиокислот — бетаин и многоатомные спирты (рис. 4. 5). Особое значение в осморегуляции имеет свободный пролин — стрессовый метаболит, содержание которого сильно возрастает при засухе, что связывают с распадом хлорофилла, белков. Установлена связь пролина с АБК, поскольку последняя увеличивает его содержание. .

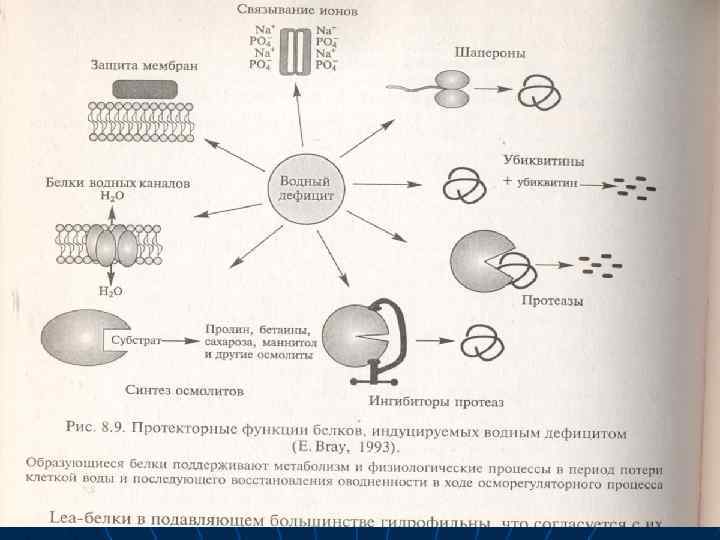
Пролин образуется из органических кислот — интермедиатов дыхания, и азота нитратов, которые особенно интенсивно накапливаются в сосудистых клетках и эпидермисе. Он синтезируется из глицина, а бетаин — из аминоспирта холина. входящего в состав фосфолипидов. Содержание этих предшественников увеличивается в стрессовых условиях в связи с активацией гидролитических процессов и распадом белков и липидов В условяях водного дефицита активируется синтез не только низкомолекулярных осмотиков, но и различных стрессовых белков, в том числе и белков, удерживающих воду.

n n n n При засухе накапливается осмотин — катионный белок. Его содержание может составлять 12% от общего содержания белка. Он локализован в вакуолях и везикулах тонопласта. Стимулируют синтез осмотина и его м. РНК и АБК. При обезвоживании синтезируются белки — дегидрины семейства LEА. Эти белки при нормальном водоснабжении растений обычно синтезируются в зародышах на стадиях позднего эмбриогенеза, когда происходит естественное обезвоживание, созревающих семян.

n n n Различные представителя этого семейства белков взаимодействуют с клеточными структурами предохраняют их от деградации при обезвоживании, связывают воду. В состав стрессовых белков входят также аквапорины — каналы для поступающей воды, которые облегчают водный транспорт через мембраны в осмотически завпсимых ситуациях. У растений найдены два типа аквапоринов — в плазматической и вакуолярной мембранах.

n n n n Функциональные белки, способствующим повышению устойчивости к засухе - многочисленные гидролазы, - ферменты протеазы, - ингибиторы протеаз, *- ферменты биосинтеза осмотиков. При водном стрессе синтезируются регуляторные белки, которые участвуют в экспрессии генов в сигнальной трансдукции. У многих растений. промежуточных между С 3 - С 4 или Сз- и САМ-семействами, под действием засухи происходит подавление С 4 - или САМ-фотосинтеза.

n n Это обусловлено экспрессией главных ферментов указанных альтернативных путей фиксации углекнслоты: ФЕПкарбоксилазы и НАДФ-малик-энзима. Например, при недостатке воды обнаружена пятикратная активная изоформы ФЕП-карбоксилазы, которая обычно в молодых растениях проявляет низкую экспрессию. В результате в условиях засухи в 100 раз увеличивается содержание м. РНК и активность САМ-специфичной ФЕПкарбоксилазы. т. е. происходит индукция САМ пути, способствующая увеличению эффективности использования воды. При засухе увеличивается уровень окисленных радикалов кислорода — супероксидов, и вместе с тем экспрессируются некоторые изоформы СОД, что ведет к удалению радикалов и увеличению устойчивости.

n n n Накопление указанных осмотически активных веществ обеспечивает высокую водоудерживающую способность цитоплазмы. Благодаря осмотической регуляции растений при засухе сохраняются тургор и открытость устьиц, а следовательно, возможность нормального роста и поддержание двух физиологических процессов. Степень осморегуляции специфична для видов

n n n Помимо осмотической регуляции в обеспечении толерантности клетки важную роль играет стабилизация клеточных мембран (рис. 4. 4). У растений, устойчивых к засухе, - предотвращение разрушения бислойной структуры липидов и соответствующие структурные и конформационные изменения белков мембран. Этому способствует накопление при обезвоживании ди- и олигосахаридов. Они взаимодействуют с полярными головками фосфолипидов, не допуская слипания мембран.
. В")
n n Эти соединения противостоят также повышению температуры фазового перехода гель—жидкий кристалл (гель—золь). В результате целостность мембран у организмов, устойчивых к засухе, сохраняется лучше. Так, плазмалемма у приспособленных к засухе растений теряет барьерные свойства в ходе дегидратации при значительно более низком содержании воды, чем у чувствительных. Сохранение стабильности мембран важно для

n n Стабильность мембран необходима для поддержания энергетических процессов — аэробного дыхания и фотосинтеза У мезофитов, различающихся по устойчивости к засухе, кривые интенсивности дыхания в зависимости от степени обезвоживания отличаются по положению максимума.
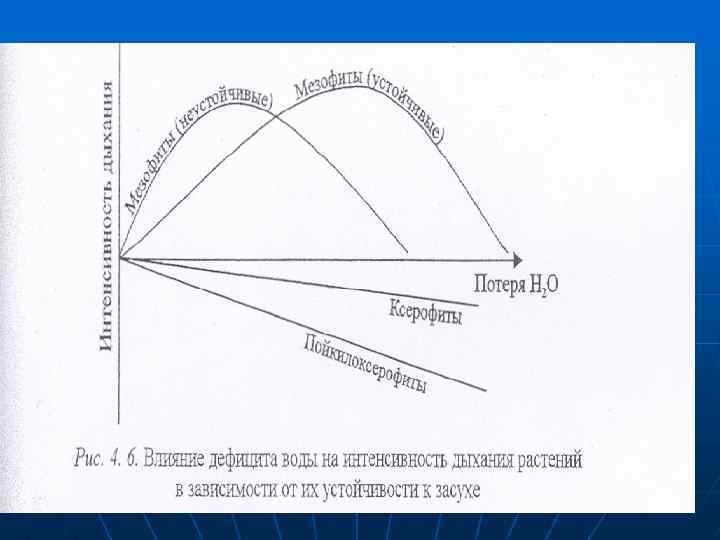

n n n У ксерофитов с потерей воды дыхание долгое время не меняется. т. е. их дыхательные системы более устойчивые к обезвоживанию. У пойкилоксерофитов, например, фиалки сербской (Ramondica serbica), наоборот, интенсивность дыхания определяется насыщенностью клеток водой (рис. 4. 7). Дыхание при засухе. - При засухе возрастает доля ПФП по сравнению с дихотомическим. Это явление связывают с потребностью в увеличении оттока восстановленных
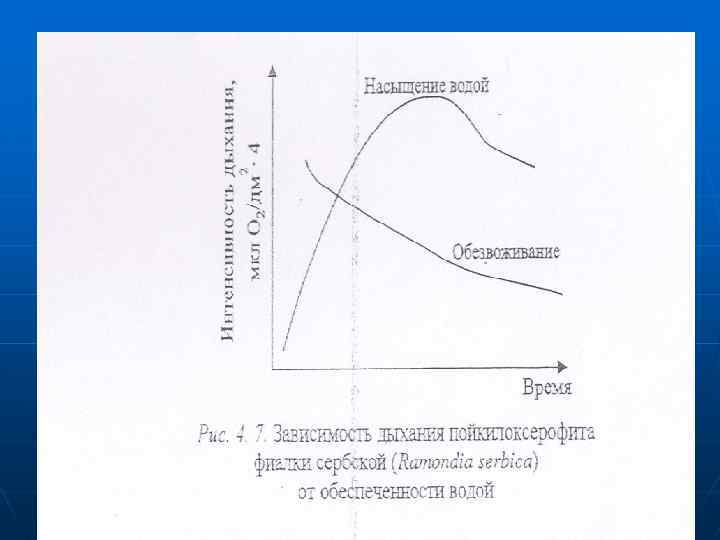

n n - Усиливается активность внемитохондриальных систем окисления в результате ослабления переноса электронов в ЭТЦ митохондрий при засухе, что связано с обеспечением детоксикации продуктов распада. Обезвреживание свободных радикальных соединений, которые образуются при засухе. как и при других стрессовых воздействиях, за счет

n n Детоксикация образующегося при протеолизе аммиака осуществляется с участием органических кислот, количество которых возрастает при засухе н высоких температурах за счет активации дыхания. С необходимостью обезвреживания накапливающихся при засухе

n n При засухе –-новые изозимы пероксидазы. Поскольку пероксидаза частично выполняет функции ИУК-оксидазы, увеличение числа ее изозимов объясняется необходимостью разрушения с их помощью ИУК и торможением роста. Повышение активности этого фермента н расширение ее изоэнзимного спектра при засухе может быть связано с разрушением пероксида водорода и ограничением свобнорадикальных процессов. Пероксидаза активируется также под влиянием этилена, уровень которого, а также АБК возрастает при недостатке влаги. Количество же гормонов-активаторов роста: гиббереллинов (ГБ), цитокинина (ЦК). наоборот. падает.
. Работа устьичного аппарата основывается")
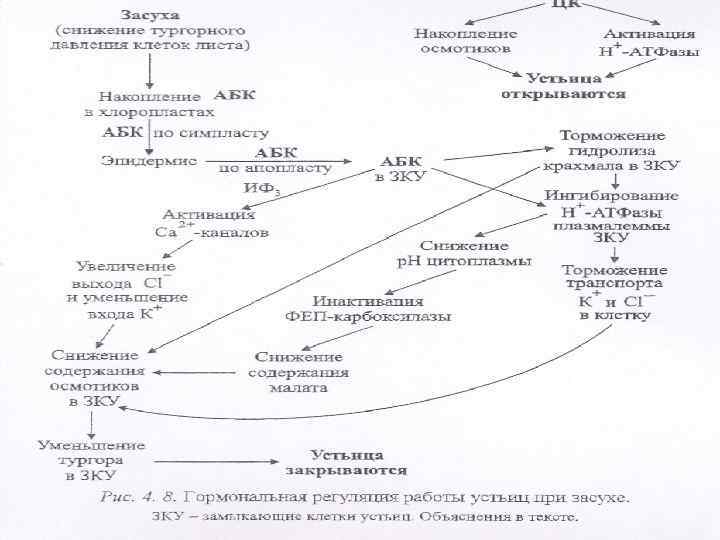
n n Накопление АБК вызывает закрывание устьиц (рис. 4. 8). Работа устьичного аппарата основывается на осморегуляции замыкающих клеток устьиц, которые благодаря ассимметрическому утолщению клеточной стенки открывают устьичную щель в тургесцентном состоянии и закрывают ее при потере тургора.

n n n Снижение тургорного давления в клетках при обезвоживании резко активирует синтез АБК, которая накапливается главным образом в хлоропластах клеток листа. АБК выходит из хлоропластов и по симпласту транспортируется в эпидермис. Оттуда по апопласту гормон попадает в замыкающие клетки устьиц, где инактивирует работу Н-АТФазы

n n n Инактивация протонной помпы снижает поступление ионов калия и хлора (вторичный активный транспорт тормозится при ингибировании первичного). Это приводит к закислению цитоплазмы. Низкое значение р. Н инактивирует ФЕП-карбоксилазу оптимум активности которой находится в щелочной среде, и ведет к снижению уровня малата. Через фосфоинозитольный путь АБК активирует кальциевые каналы. Нарастание содержания калия в цитоплазме ведет к деполяризации мембраны, активные каналы плазмалеммы открываются, а ионы хлора покидают клетку. Одновременно закрываются калиевые каналы, что приводит к понижению поступлению в клетку кальция.

n n В результате этих процессов содержание осмотиков в замыкащих клетках падает, они теряют тургор и устьица закрываются. В отличие от АБК накопление ЦК способствует открыванию устьиц вследствие активации протонной помпы и накопления осмотиков в замыкающих клетках, что весьма неблагоприятно при

n АБК участвует в осморегуляции и других клеток. Активирует синтез других целого ряда стрессовых белков: аквапоринов, дегидринов, осмотиков, ферментов синтеза пролина и т. п. n Это способствует запасанию гидратной воды, тормозит синтез РНК и обычных белков. n n n Действие АБК затрагивает метаболизм других фитогормонов: - усиливает синтез этилена, - оказывает ингибиторное воздействие на транспорт ауксина, - задерживает синтез и ускоряет распад цитокинина.

n n n Изменение гормонального статуса приводит к торможению роста, при этом потребности организма в воде уменьшаются. Обмен веществ переводится в режим покоя, энергетические процессы переключаются ва поддержание целостности растения и репарацию повреждений. У пойкилоксерофитов, переходящих при

n n n Изменения в содержании фитогормонов, но более кратковременные наблюдаются и у мезофитов при засухе. Для синтеза стрессовых белков и успешного восстановления обмена после засухи важно сохранение ненарушенным генетического аппарата клетки. Защита нуклеиновых кислот от засухи осуществляется различными стрессовыми белками — шаперонами. Они в состоянии долгое время поддерживать целостность ДНК ( даже при абсолютном обезвоживании (у пойкилоксерофитов).

n n n Сигнал засухи воспринимается осмосенсорами, которыми могут быть механические изменения плазматической мембраны или состояния цитоскелета, окислительный взрыв, т. е. накопление активных форм кислорода (рис. 4. 9). В цитоплазме клетки имеются белковые факторы транскрипции (трансфакторы), связывающиеся с элементами дегидратации — регуляторными элементами (промоторами) генов в ядре

n n Трансдукция сигнала из цитоплазмы в ядро связана с участием ионов кальция, инозитольного цикла, фосфорилированием, которое осуществляется Са- или МАРзависимыми протеинкиназами. В результате передачи сигнала в ядро происходят изменения экспрессии генов, которые влекут за собой биохимические и фзиологкческие изменения — синтез стрессовых белков и повышение устойчивости.

n n n Ингибитором синтеза белков в условиях засухи или засоления, холода, высокой температуры и даже кислородной недостаточности, т. е. причиной нарушения осмотической регуляции, может быть и АБК. Последовательность компонентов реакций при действии АБК подробно описаны на рис. 4. 9. Их дополняет только иной регуляторкый белок, относящийся к белковым факторам трансдукции , и регуляторный элемент генов — АВRE, т. е. элементы ответа на АБК.
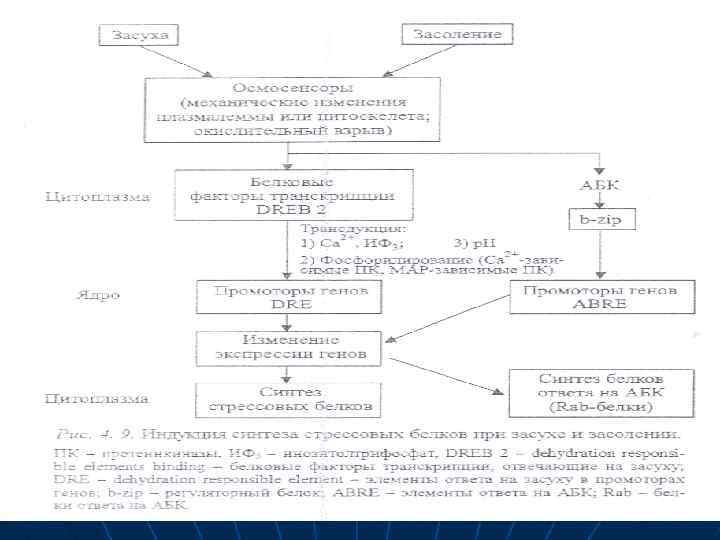

n n n n В результате синтезируются АБК-чувствительные или Rab-белки. При обычном водоснабжении их нет в листьях, но недостаток воды вызывает накопление в листьях АБК, которое индуцирует синтез этих гидрофильных белков, устойчивых к деградации Итак, к клеточным механизмам толерантности относятся: 1) осмотическая регуляция; 2) стабилизация клеточных мембран; З) перестройки в процессе дыхания 4) детоксикация токсических продуктов

n n n Помимо механизмов устойчивости к действию обезвоживания на клеточном уровне включаются защитные реакции и на уровне организма (рис. 4. 4). в которых участвует трофическая система регуляции. Нарушение работы фотосинтетического аппарата должно приводить к ингибированию оттока ассимилятов и нарушению донорно-акцепторных взаимодействий в организме. Передвижение ассимилятов существенно ингибируется только при очень сильном водном дефиците. Эта относительная нечувствительность передвижения ассимилятов позволяет растениям мобилизовать и использовать их резервы, например для роста семян, даже при довольно сильном воздействии. Способность поддерживать транспорт углеводов -
 генеративных")
n n К механизмам устойчивости относится и формирование минимума (вследствие опадания ила недоразвития) генеративных органов, которые растение в состоянии обеспечить питательными веществами. Это обусловлено конкретными отношениями между органами за воду, питательные и другие

n Механизмы защиты на популяционном уровне также участвуют в формировании устойчивости растений, поскольку выживают именно те растения, у которых норма реакции находится в широком диапазоне экстремальных факторов, т. е. существует внутрипопуляционная вариабельность уровня устойчивости.

n n n n Повышение устойчивости к засухе Для повышения устойчивости к водному дефициту в практических целях необходимо знание механизмов устойчивости к засухе. Например, при учете гормональных перестроек, которые происходят в обмене веществ в таких условиях, можно использовать обработку растений экзогенно вводимыми гормонами. Действие АБК: - уменьшение клеток количества устьиц на листе, - увеличение опушения листьев - снижение реагирования организма на водный дефицит.

n Внесение цитокинина. гиббереллина, ауксина во время засухи усугубляет ее отрицательный эффект. Однако после воздействия в период репарации состояние растений значительно улучшается. n n Обработка семян до засухи цитокининовыми препаратами. Защитное действие цитокинина связывают с его влиянием на структуру и функции макромолекулярных компонентов клетки. Обработка гормонами важна и в целях повышения деятельности белок-синтезирующих систем. При предпосевной закалке к засухе- функциональная активность рибосом стимулируется, что также определяется большой устойчивостью белок-синтезируюшей системы.

n n n У закаленных растений сохраняется способность к синтезу белка при частичном обезвоживании. Предпосевное замачивание и высушивание семян. Полученные из таких семян растения отличаются ксероморфной структурой и повышенной засухоустойчивостью. Стойкость к засухе возрастает при достаточном обеспечении калием, который улучшает поглощение и использование воды, снижается транспирация в результате более эффективной регуляции работы устьичного аппарата, поступления связывания воды.

n n n Благодаря таким обработкам в клетке развиваются и глубже проникают в почву корневые системы, как работа фотосинтетического аппарата, так и отток ассимилятов в репродуктивные органы, поскольку калий участвует в транспорте сахарозы. Будучи кофактором многих реакций фотосинтеза и дыхания, калий оказывает стимулирующее действие на поддержание основных физиологических процессов. Итак, закалке, или акклимации к осмотическому стрессу, способствует обработка защитными веществами (сахарами. АТФ, пролином), гормонами (цитокинином), а также препаратом цитокининоподобного действия — картолином). калием.

n n n В связи с выявлением четкой зависимости устойчивости к засхе от содержания осмотиков в последние годы используется введение в геном растений генов, кодирующих энзимы, которые катализируют образование осмотически активных продуктов. Эти методы генной инженерии позволяют получить устойчивые к засухе трансгенные растения. Так например, ген который кодирует бифункциональные энзимы, обладают каталитическими активностями γ-глутамилкиназы и глутамил- γ -семиальдегид-дегидрогеназы, использовался для трансформации табака. у

n n Этот энзим катализирует превращение глутамата ∆пирролин-5 -карбоксилата, который затем восстанавливается до пролина. Трансгенные растения табака продуцировался в 1018 раз больше пролина и общей биомассы по сравнению с контрольными объектами. Повышение концентрации осмотика способствовало увеличению осмотического потенциала и облегчению выживания в условиях водного дефицита. В листья табака вводили также ген , кодируемый

n n Трансгенные растения табака приобретали устойчивость к засухе при возрастании уровня нередуцирующего полисахарида трегалозы, также обладающей осмозащитными функциями. Близкие данные получены при стимулировании синтеза фруктана и ряда других веществ с осмотическими свойствами. Трансгенные растения табака, у которых удалось экспрессировать СОД, также оказались более толерантными к дефициту воды. Таким образом, повышение осмотического потенциала трасгенных растений за счет активирования синтеза защитных осмофильных веществ или ферментов системы детоксикации токсических веществ является новым и перспективным способом повышения устойчивости растений к засухе.
2 Лек.2016.ppt