

Lection 2.ppt
- Количество слайдов: 26
") ЛЕКЦИЯ 2. Структура нуклеиновых кислот (ДНК)
ЛЕКЦИЯ 2. Структура нуклеиновых кислот (ДНК)
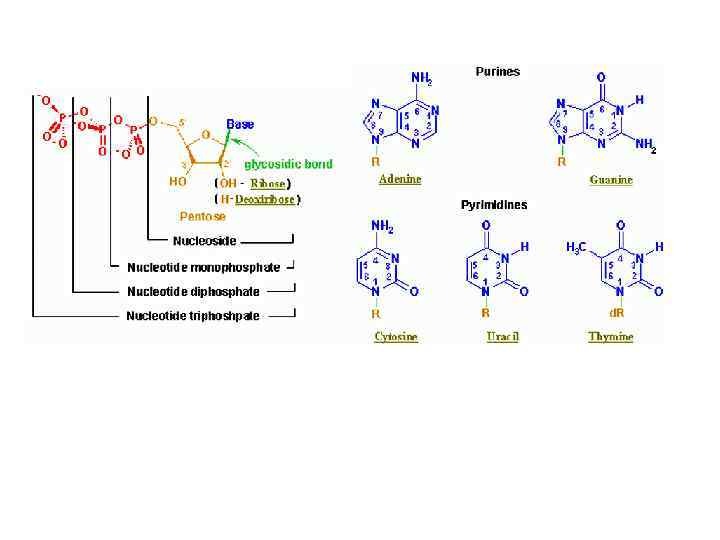
 Первичная структура нуклеиновых кислот Нуклеотид
Первичная структура нуклеиновых кислот Нуклеотид
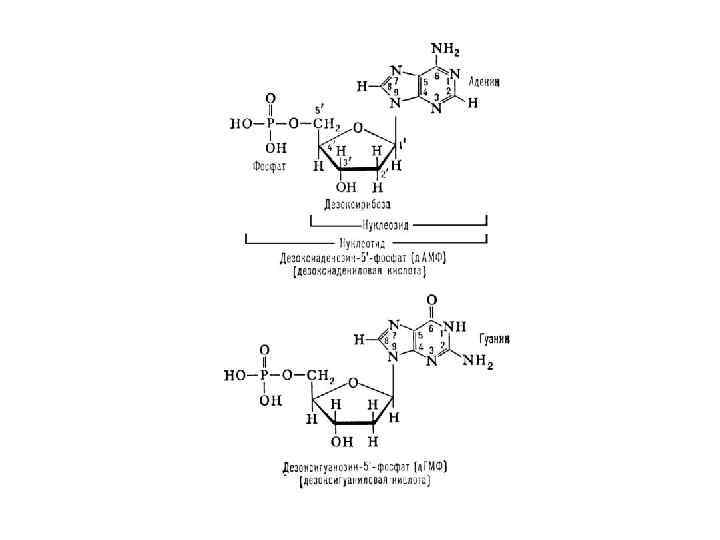
 Нуклеотид
Нуклеотид
") Гетероциклические основания Пентоза (5 углеродов)
Гетероциклические основания Пентоза (5 углеродов)
 Структурные формулы минорных пуриновых и пиримидиновых оснований ДНК РНК
Структурные формулы минорных пуриновых и пиримидиновых оснований ДНК РНК
 – плоская конформация энергетически невыгодна") Гетероциклические основания – плоская конформация Пентоза (5 углеродов) – плоская конформация энергетически невыгодна
Гетероциклические основания – плоская конформация Пентоза (5 углеродов) – плоская конформация энергетически невыгодна
 Две конформации углеводных остатков S N
Две конформации углеводных остатков S N
 Анти- и син-конформации в нуклеотидах anti syn
Анти- и син-конформации в нуклеотидах anti syn
 Структура молекулы ДНК Предпосылки: - - ДНК – полимер, состоящий из нуклеотидов, соединенных 3’-5’- фосфодиэфирной связью. Состав нуклеотидов подчиняется правилам Чаргаффа: A+G=T+C, A=T, C=G. Имеются водородные связи, наблюдается денатурацияренатурация. Рентгенограмма (М. Уилкинсон, Р. Франклин) – спиральная структура, более одной нити.
Структура молекулы ДНК Предпосылки: - - ДНК – полимер, состоящий из нуклеотидов, соединенных 3’-5’- фосфодиэфирной связью. Состав нуклеотидов подчиняется правилам Чаргаффа: A+G=T+C, A=T, C=G. Имеются водородные связи, наблюдается денатурацияренатурация. Рентгенограмма (М. Уилкинсон, Р. Франклин) – спиральная структура, более одной нити.
 – двойная спираль ДНК - правовинтовая") Модель Уотсона и Крика ( 1953 г. ) – двойная спираль ДНК - правовинтовая спираль; - двунитевая; - нити имеют антипараллельную ориентацию; - комплементарность оснований; - основания связаны водородными связями (Pr-Pu); - гидрофильные пентозофосфатные остовы цепей расположены на внешней стороне двойной спирали; Дж. Уотсон род. в 1928 г. Ф. Крик 1916 -2004
Модель Уотсона и Крика ( 1953 г. ) – двойная спираль ДНК - правовинтовая спираль; - двунитевая; - нити имеют антипараллельную ориентацию; - комплементарность оснований; - основания связаны водородными связями (Pr-Pu); - гидрофильные пентозофосфатные остовы цепей расположены на внешней стороне двойной спирали; Дж. Уотсон род. в 1928 г. Ф. Крик 1916 -2004
 1953 г. – James Watson, Francis Crick предложили модель структуры ДНК. 1962 г. – получили Нобелевскую премию за трехмерную структуру ДНК. Модель трехмерной структуры ДНК: 1. Молекула состоит из 2 нитей. 2. Нити образуют спираль. 3. Нити антипараллельны (5’-3’ и 3’-5’). 4. Сахаро-фосфатный остов, остатки оснований располагаются между нитями, фосфаты придают молекуле ДНК отрицательный заряд. 5. Пиримидин связывается только с пурином. Молекула имеет постоянную ширину 2 нм. 6. Спираль имеет желобки, позволяющие специфически связываться с белками. 7. Нити комплементарны – А-Т, G-C.
1953 г. – James Watson, Francis Crick предложили модель структуры ДНК. 1962 г. – получили Нобелевскую премию за трехмерную структуру ДНК. Модель трехмерной структуры ДНК: 1. Молекула состоит из 2 нитей. 2. Нити образуют спираль. 3. Нити антипараллельны (5’-3’ и 3’-5’). 4. Сахаро-фосфатный остов, остатки оснований располагаются между нитями, фосфаты придают молекуле ДНК отрицательный заряд. 5. Пиримидин связывается только с пурином. Молекула имеет постоянную ширину 2 нм. 6. Спираль имеет желобки, позволяющие специфически связываться с белками. 7. Нити комплементарны – А-Т, G-C.
 1 нм=10Å
1 нм=10Å
 Взаимодействие между основаниями - Комплементарные пары: А-Т – две водородные связи С-Г – три водородные связи - Стэкинг (внутри одной нити) (staking - укладываться в стопку) ван-дер-ваальсовы силы водородные связи N-гликозидная связь фосфодиэфирная связь
Взаимодействие между основаниями - Комплементарные пары: А-Т – две водородные связи С-Г – три водородные связи - Стэкинг (внутри одной нити) (staking - укладываться в стопку) ван-дер-ваальсовы силы водородные связи N-гликозидная связь фосфодиэфирная связь
 и В (C 2’-эндо-) формы, правая спираль Z") Конформационные состояния ДНК А (C 3’-эндо-) и В (C 2’-эндо-) формы, правая спираль Z форма
Конформационные состояния ДНК А (C 3’-эндо-) и В (C 2’-эндо-) формы, правая спираль Z форма
 Характеристика некоторых конформационных состояний ДНК
Характеристика некоторых конформационных состояний ДНК
 B- и Z-формы Переход В в Z разрушает нуклеосомную структуру.
B- и Z-формы Переход В в Z разрушает нуклеосомную структуру.
 Функциональная роль различных форм ДНК А – шпильки он. РНК, ДНК в спорах бактерий (споровые белки превращают В в А, повышается в 10 раз УФ-резистентность), гибриды ДНК-РНК (ДНК и РНК в C 3’-эндо, переход ДНК из В, т. е. в передаче информации (транскрипция - затравки, репликация -праймеры). Одноцепочечный и двуцепочечный стэкинг. В – обычная нуклеосомная форма (репликация), т. е. в умножении информации. Одноцепочечный стэкинг. С – надмолекулярная структура хроматина, т. е. при хранении информации. Д – фаговая ДНК. Z – poly(d. G-d. C), высокая концентрация солей. Стэкинг только между С, С 3’ C 2’ син анти отсутствует большой желобок, регуляторная роль, например в транскрипции, обеспечивает сверхспирализацию в междисковых областях политенных хромосом. Левая спираль. Стэкинг между остатками цитозина противоположных цепей.
Функциональная роль различных форм ДНК А – шпильки он. РНК, ДНК в спорах бактерий (споровые белки превращают В в А, повышается в 10 раз УФ-резистентность), гибриды ДНК-РНК (ДНК и РНК в C 3’-эндо, переход ДНК из В, т. е. в передаче информации (транскрипция - затравки, репликация -праймеры). Одноцепочечный и двуцепочечный стэкинг. В – обычная нуклеосомная форма (репликация), т. е. в умножении информации. Одноцепочечный стэкинг. С – надмолекулярная структура хроматина, т. е. при хранении информации. Д – фаговая ДНК. Z – poly(d. G-d. C), высокая концентрация солей. Стэкинг только между С, С 3’ C 2’ син анти отсутствует большой желобок, регуляторная роль, например в транскрипции, обеспечивает сверхспирализацию в междисковых областях политенных хромосом. Левая спираль. Стэкинг между остатками цитозина противоположных цепей.
 Формы ДНК • Линейная • Кольцевая: - релаксированная - сверхспирализованная 1 - линейная одноцепочечная ДНК - бактериофаг φХ 174 и другие вирусы; 2 - кольцевая одноцепочечная ДНК вирусов и митохондрий; 3 - кольцевая двойная спираль ДНК.
Формы ДНК • Линейная • Кольцевая: - релаксированная - сверхспирализованная 1 - линейная одноцепочечная ДНК - бактериофаг φХ 174 и другие вирусы; 2 - кольцевая одноцепочечная ДНК вирусов и митохондрий; 3 - кольцевая двойная спираль ДНК.
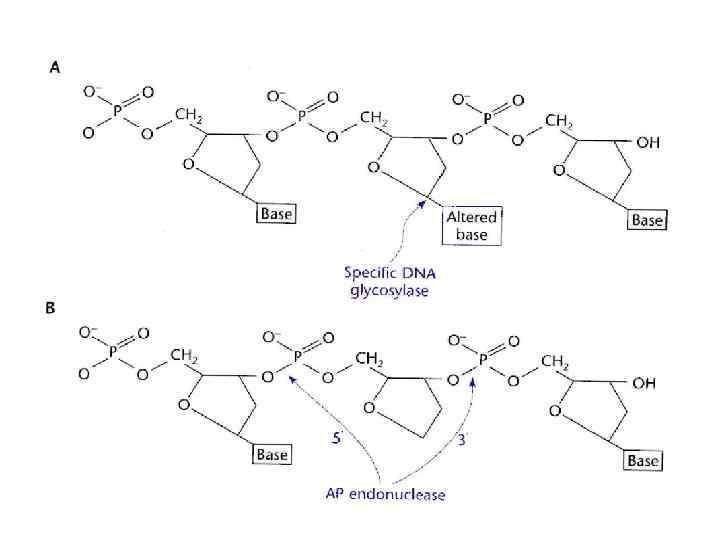
 Основные повреждения ДНК • • повреждения одиночных нуклеотидов; повреждения пары нуклеотидов; разрыв одной или двух цепей ДНК; образование поперечных сшивок между основаниями одной цепи или разных цепей ДНК.
Основные повреждения ДНК • • повреждения одиночных нуклеотидов; повреждения пары нуклеотидов; разрыв одной или двух цепей ДНК; образование поперечных сшивок между основаниями одной цепи или разных цепей ДНК.
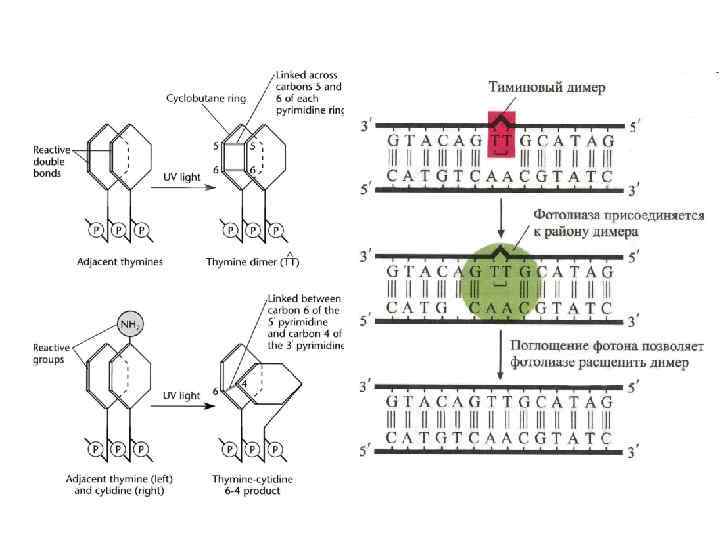
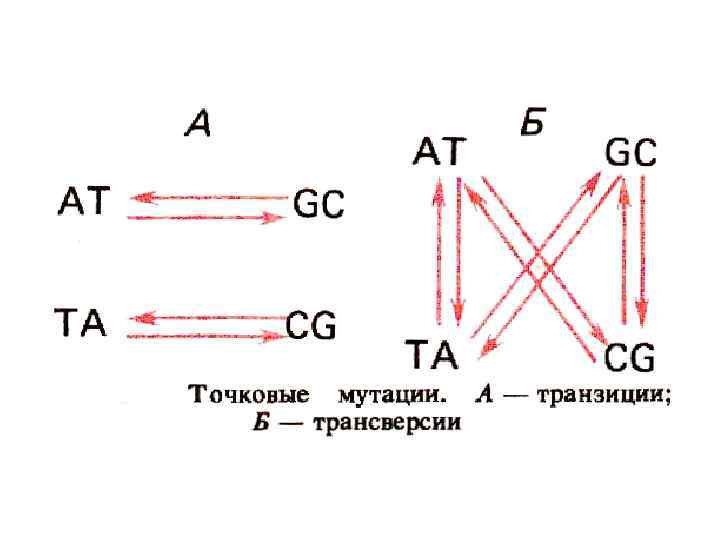
 IV -") Повреждения нуклеотидов I - дезаминирование II - апуринизация III – метилирование (алкилирование) IV - размыкание кольца V – окисление (8 -oxo-G - A» GC→AT C→ 5 -OH-U-C; 5 -OH-C-A C→G C→T) VI - образование тиминовых димеров
Повреждения нуклеотидов I - дезаминирование II - апуринизация III – метилирование (алкилирование) IV - размыкание кольца V – окисление (8 -oxo-G - A» GC→AT C→ 5 -OH-U-C; 5 -OH-C-A C→G C→T) VI - образование тиминовых димеров
 впервые выделил") Основополагающие открытия • • • • 1869 – Ф. Мишер (F. Misher) впервые выделил ДНК из лейкоцитов и молок лосося. 1935 – А. Н. Белозерский выделил ДНК из растений. 1940 – У. Эстбюри (W. Astbury) получил первую рентгенограмму ДНК. 1944 – О. Т. Эвери (O. T. Avery) установил, что ДНК (а не белок, как полагали ранее) является носителем генетической информации. 1953 – Дж. Уотсон и Ф. Крик (J. Watson, F. Crick) создали модель двойной спирали ДНК на основе рентгенограмм, полученных Р. Франклин и М. Уилкинсом (R. Franklin, M. Wilkins). 1966 - М. Ниренберг, С. Очоа и Х. -Г. Корана (M. Nirenberg, S. Ochoa, H. -G. Khorana) расшифровали генетический код. 1975 -1977 – Ф. Сангер (F. Sanger), а также А. Максам и У. Гилберт (A. Maxam, W. Gilbert) разработали методы быстрого определения первичной структуры ДНК. 1976 – Ф. Сангер (F. Sanger) расшифровал нуклеотидную последовательность ДНК фага φХ 174. 1976 – У. Гилберт (W. Gilbert) открыл мозаичное строение генов эукариот. 1982 – созданы банки нуклеотидных последовательностей: Gen Bank (USA) и EMBL (Европа). 1996 – был секвенирован первый бактериальный геном. 1997 – секвенирован геном дрожжей Saccharomyces cerevisiae. 2000 – секвенирован геном человека.
Основополагающие открытия • • • • 1869 – Ф. Мишер (F. Misher) впервые выделил ДНК из лейкоцитов и молок лосося. 1935 – А. Н. Белозерский выделил ДНК из растений. 1940 – У. Эстбюри (W. Astbury) получил первую рентгенограмму ДНК. 1944 – О. Т. Эвери (O. T. Avery) установил, что ДНК (а не белок, как полагали ранее) является носителем генетической информации. 1953 – Дж. Уотсон и Ф. Крик (J. Watson, F. Crick) создали модель двойной спирали ДНК на основе рентгенограмм, полученных Р. Франклин и М. Уилкинсом (R. Franklin, M. Wilkins). 1966 - М. Ниренберг, С. Очоа и Х. -Г. Корана (M. Nirenberg, S. Ochoa, H. -G. Khorana) расшифровали генетический код. 1975 -1977 – Ф. Сангер (F. Sanger), а также А. Максам и У. Гилберт (A. Maxam, W. Gilbert) разработали методы быстрого определения первичной структуры ДНК. 1976 – Ф. Сангер (F. Sanger) расшифровал нуклеотидную последовательность ДНК фага φХ 174. 1976 – У. Гилберт (W. Gilbert) открыл мозаичное строение генов эукариот. 1982 – созданы банки нуклеотидных последовательностей: Gen Bank (USA) и EMBL (Европа). 1996 – был секвенирован первый бактериальный геном. 1997 – секвенирован геном дрожжей Saccharomyces cerevisiae. 2000 – секвенирован геном человека.