Лекция 2 (Ископаемые беспозвоночные ).ppt
- Количество слайдов: 10



Лекция 2 Ископаемые беспозвоночные

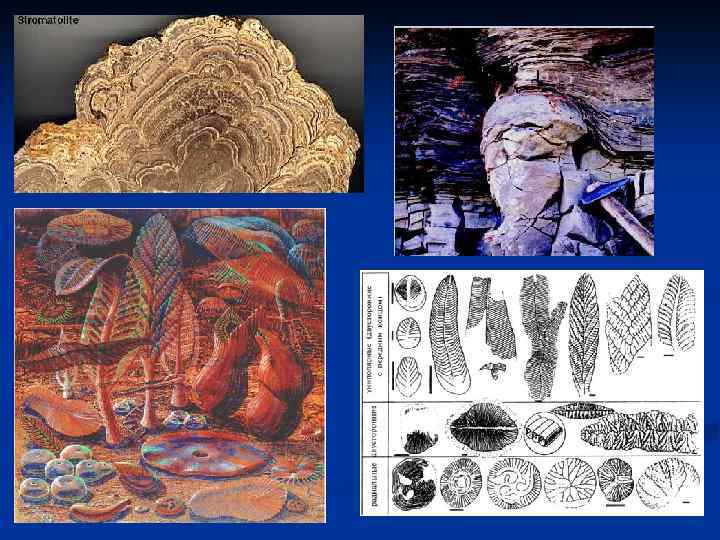
Остатки первых живых организмов датируются возрастом 3, 5 млрд. лет. Это строматолиты- слоистые камни из углекислых кальция и магния. Колонии бактерий и цианобактерий использовали в своем обмене веществ карбонаты и откладывали их поверхностном слое колонии. Потом выше поселялись новые колонии, и камень рос вверх. Такие строматолиты образуются и теперь. Как и теперь, тогда этот процесс шел на мелководье, но морское мелководье в протерозое простиралось на огромные расстояния от берега. Моря и суши в привычном для нас понимании тогда не было, так как из-за отсутствия наземных растений эрозия горных пород происходила очень быстро. Первые экосистемы состояли лишь из продуцентов (цианобактерии) и редуцентов (бактерии). Вопрос о том, кто из них появился первым, (цианобактерии) неправомочен: если бы кто-то из них возник раньше, он бы очень быстро подорвал собственные пищевые ресурсы или загрязнил среду отходами жизнедеятельности. И те, и другие должны были возникнуть одновременно. Консументы появились позже и существенно ускорили эволюцию, так как началась гонка вооружений между хищниками и жертвами. Теоретически консументы не обязательны, но чем больше звеньев в цепи питания, тем стабильнее экосистема. Широкое распространение в протерозое строматолитов можно объяснить именно отсутствием потребителей тех, кто их создавал. Кроме того, в кембрии произошло массовое появление раковинных организмов, конкурировавших с создателями строматолитов за кальций в морской воде. Можно утверждать, что многоклеточные прокариоты в истории жизни на Земле не появлялись никогда. Видимо, этому препятствует характер строения их клеточной стенки, не позволяющий осуществлять сколько-нибудь существенные межклеточные взаимодействия. Достоверных ископаемых одноклеточных эукариот неизвестно, но многоклеточные появились, по меньшей мере, 700 млн. лет назад. Широкую известность имеет комплекс вендских животных, найденный в Эдиакаре (Австралия). Такая же фауна известна и из России, с Белого моря. Некоторые из вендских животных похожи на представителей современных типов: кораллы, медуз, червей и членистоногих, а некоторые современных аналогов не имеют. Теперь считается, что никакие из них современным животным не родственны. Все они мягкотелые. Среди них очень велико число форм, которым приписывали специализацию на перехвате медленно падающих на дно осадков – животные в форме торчащих со дна пышных перьев и распластанных по дну пирогов. Однако изучение остатков с очень высокой степенью сохранности обнаружило, что ни одного из этих созданий не было рта. По-видимому, вендские животные были нашпигованы автотрофными бактериями, которые и снабжали их пищей, как это имеет место у современных погонофор. Все они обладали ещё двумя общими особенностями строения: “скользящей симметрией”, как у половинок застежки “молния”, тело же их состояло из чередования вздутий и перехватов, как стеганое одеяло; и изометрическим ростом. Возможно, что среди вендобионтов также никто никого не ел. В кембрии появляются представители почти всех современных типов. Археоциаты, которых теперь относят к губкам, несмотря на отсутствие у них спикул, считались вымершими в конце кембрия, но недавно был найден живой представитель. Многие кораллы, жившие с кембрия по спикул, пермь, были непохожи на современных: например, одиночные, как актинии, но с мощным известковым скелетом; свободно лежащие на дне; пермь, замыкающиеся крышечкой. С ордовика известны мшанки, их родственники брахиоподы – с позднего докембрия. И те, и другие, видимо, произошли от форонид, которые не имеют двустворчатой раковины, как брахиоподы, и не образуют колоний, как мшанки. Все три типа форонид, брахиоподы, имеют лофофор – раскручивающиеся две руки с гребенкой для улавливания пищевых частиц. На гребенке – реснички, двигающие пищу ко рту. Лофофор не может хорошо работать в мутной воде, поэтому как раньше, так и теперь брахиоподы более избирательны в местообитаниях, чем двустворчатые моллюски. Кишечник у брахиопод слепозамкнутый. Есть ножка, короткая у форм, крепящихся к слепозамкнутый. твердому субстрату и длинная у зарывающихся в грунт; лежачие ее вообще утратили. В отличие от двустворчатых моллюсков, створки у брахиопод не правая и левая, а верхняя и нижняя (педальная и брахиальная). Даже питаясь, они раздвигают их нешироко. Существует два брахиальная). класса брахиопод – замковые и беззамковые. Брахиоподы живут и сейчас, но современные по сравнению с палеозойскими малочисленны. беззамковые. От червей в большинстве случаев остаются лишь следы. Тем не менее паразитические плоские черви обнаружены у насекомых карбона, перми и третичного периода; паразитические круглые черви – у насекомых карбона, мела, третичного и четвертичного периодов. Животные, похожие на кольчатых червей, встречаются с венда; несомненные приапулиды – с кембрия, но разнообразие последних указывает на более раннее происхождение. Немертины и скребни в ископаемом состоянии пока не обнаружены.

Среди кембрийских животных есть такие, которых нельзя отнести к современным типам. Из местонахождения Бёджесс (Канада) известны причудливые червеобразные и похожие на членистоногих существа. Ротовой диск ракоподобного Anomalocaris (это самый крупный представитель Бёджесской фауны, до 70 см длиной) был в начале описан как медуза и по своему строению свидетельствует о том, что ракам это животное совсем не родственно. Marella напоминает нечто среднее между трилобитом и раком, а Hallucigenia не похожа вообще ни на кого, хотя может быть родней тихоходкам и онихофорам. В кембрии происходил мощный процесс, названный артроподизацией. Мода на членистые конечности, как и на раковины, характерная для того времени, возможно, объясняется тем, что как только у одной филогенетической линии появлялся такой признак, другие похожие линии оказывались в проигрышном положении, если его не приобретали. Почти наверняка современные ракообразные, хелицеровые и одноветвистые родственны другу не больше, чем Бёджесским диковинам. Бёджесские полихеты характеризуются мощно развитыми жабрами вдобавок к параподиям. В колониях палеозойских кораллов были широко распространены полихеты – комменсалы. Еще более, чем полихеты, в Бёджесской фауне многочисленны приапулиды, редкие теперь. Из этого же местонахождения известна онихофора Aysheaia. В отличие от современных, она жила в море и ползала по губкам, о чем говорит нахождение вместе с их спикулами. Самой характерной и многочисленной группой морских палеозойских членистоногих были трилобиты, названные так за тело, поделенное вдоль на три доли. Известно около 2 тыс. родов и более 10 тыс. видов трилобитов. Как и в отношении современных членистоногих, трилобитов принято выводить под аннелид, но эта позиция не бесспорна. Эдиакарская Spriggina похожа на трилобитов, но не обязательно родственна им. Трилобиты создавали огромное разнообразие форм, сохраняя общий план строения: осевая часть и плевральные по бокам. Спереди находился цефалон с глабелью посередине; позади торакс и пигидий, сегменты которого могли сливаться. В глабели располагался желудок, сзади от нее снизу был рот, а сверху сердце. Цефалон нес также три-четыре пары конечностей, членистые антенны и фасеточные глаза. Глаза трилобитов с фасетками, покрытыми общей оболочкой, называются голохроическими; с разделенными фасетками – шизохроическими. Размеры глаз и число фасеток сильно колебались, у некоторых их не было вовсе. Конечности, как у ракообразных, были двуветвистыми, и притом однотипными. Базиподит у каждой нес жевательную ветвь – гнатобазис, эндоподит служил для хождения, а экзоподит – для дыхания и плавания. Следы трилобитов говорят о том, что они хорошо двигались не только вперед, но и вбок, и могли зарываться. Сверху конечности были прикрыты плеврами. Примитивные виды с маленьким пигидием называются микропигиевыми, с пигидием большего размера – гетеропигиевыми. У изопигиевых он такой же, как цефалон; а у макропигиевых – еще больше. На пигидии открывалось анальное отверстие. Трилобиты откладывали яйца в вырытые ямки, которые потом заносило осадком. Развитие происходило с метаморфозом. У протаспидной стадии цефалон и пигидий слиты в единый щит, есть четыре пары конечностей с шипами для плавания в воде. У мераспидной стадии появляется сегмент торакса между цефалоном и пигидием; сегменты нарастали далее от передней части пигидия. Голаспидная (зрелая) стадия достигалась приобретением полного набора сегментов. Лицевые швы цефалона служили для линьки, а их форма и расположение используются для диагностики отряда трилобитов. Большинство трилобитов были детритофагами, но первые формы, по-видимому, являлись хищниками. Трилобиты едва ли были предками современных членистоногих. От одноветвистых их отделяют двуветвистые конечности, от хелицеровых – наличие антенн, от ракообразных – наличие одной пары антенн.


Появление в конце протерозоя-начале палеозоя мелких планктонных ракообразных имело далеко идущие последствия. Дело в том, что такие животные пропускают через себя в процессе питания огромные (благодаря своей численности) количества воды, вместе с пищевыми частицами захватывают минеральную взвесь и упаковывают её в быстро оседающие на дно комочки помёта. Их деятельность привела к увеличению прозрачности воды, бывшей до того очень мутной из-за быстрой эрозии горных пород на лишённой растительности суше. В результате в океане расширилась зона фотосинтеза и увеличилось количество производимого биосферой кислорода. Количество организмов, потребляющих кислород, возросло не сразу, и накопление его в атмосфере вызвало “обратный парниковый эффект” - похолодание. А поскольку планктонные ракообразные очень эффективно удаляют из воды не только несъедобные взвеси, но и съедобные, они составили мощную конкуренцию другим фильтраторам (возможно, в этом причина вымирания археоциат). Из ракообразных в ископаемом состоянии известны жаброногие, ракушковые, усоногие и высшие раки. Интересно, что некоторые ордовикские и девонские высшие раки (филлокариды) выглядят, как щитни из числа жаброногих. Первые хелицеровые известны с кембрия (это мечехвосты, живущие и ныне). Другой их класс – Eurypterida – просуществовал с ордовика по пермь. Эвриптериды имели 6 пар конечностей – хелицеры, 4 пары ходильных ног и 1 пару плавательных. У первых эвриптерид хелицеры были малы, так что пищу им приходилось удерживать передними ногами. Но у силурийского Pterygotus хелицеры превратились в огромные клешни. Тельсон эвриптерид представлял собой шип (по-видимому, ядовитый), или нёс на себе шипы, или был уплощён. Многие эвриптериды достигали в длину 2 м. Древнейшие паукообразные – скорпионы- известны с силура. Они уже были наземными; в отличие от эвриптерид, их клешни образованы из педипальп, а не из хелицер. И скорпионы, и пауки (известные с девона), в прошлом были более разнообразны, чем сейчас. Паук Megarachne из позднего карбона Аргентины обладал полуметровым размахом ног. Морские пауки, чья принадлежность к хелицеровым небесспорна, известны уже из раннего девона Германии. В настоящее время они весьма разнообразны в антарктических водах. Многоножки известны с карбона. Наиболее представительные из них- двухметровые артроплевриды, относящиеся к двупарноногим. Насекомые, как первичнобескрылые, так и крылатые, тоже известны с карбона. Они произошли, видимо, путём педоморфоза от многоножек, о чем говорят стадии эмбриогенеза последних. Зачатки крыльев на гипотетической переходной стадии, видимо, возникли на всех грудных сегментах (известны карбоновые крылатые с крыловыми зачатками на переднегруди) и могли удлинять и стабилизировать прыжок насекомого среди растительности. Понятно, что извлечь выгоду из такого приобретения могли только крупные насекомые, мелких же обладателей неподвижных крыловых зачатков сносило бы ветром. Первые обладатели подвижных крыльев относились к древнекрылым: они были неспособны складывать крылья, имели крыловые мышцы прямого действия и, видимо, сохраняли внутри крыловой пластинки живую ткань подобно современным подёнкам. Таковы были одни из самых многочисленных карбоновых насекомых – диктионевриды. Они вели открытый образ жизни и питались спорангиями и семязачатками папоротникообразных и голосемянных растений (палинофагия). Диктионевриды пришли в упадок после появления видимо, слишком эффективно охотившихся на них стрекоз, утративших живую ткань в крыловой пластинке и потому лучше летающих.


Ответом на воздушное хищничество стрекоз стало появление новокрылых насекомых, способных осваивать закрытые местообитания. Новокрылые известны с позднего карбона. Характерной особенностью насекомых и других наземных членистоногих карбона являются их крупные размеры – известна стрекоза с метровым размахом крыльев. Это может объясняться высокой концентрацией кислорода в атмосфере карбона, что важно для животных с трахейным дыханием. Карбоновые стрекозы, в отличие от современных, имели водных личинок. Прямокрылые также известны с карбона, уховёртки с перми, термиты с третичного периода (в действительности они гораздо древнее), пухоеды, вши и блохи с третичного периода (связаны с захоронениями своих хозяев, а свободноживущие могут быть гораздо древнее), трипсы с юры, полужесткокрылые и равнокрылые с перми, перепончатокрылые с триаса, жуки с перми, скорпионницы с карбона, двукрылые с триаса, ручейники с перми, бабочки (видимо, самый молодой отряд)- с мела. Переломный этап в эволюции насекомых связан с появлением в нижнем мелу цветковых растений, что спровоцировало бурную радиацию листогрызущих, а также питающихся нектаром форм. С кембрия известно большинство классов моллюсков: моноплакофоры, хитоны, скафоподы, двустворчатые, брюхоногие и головоногие. Все они появились в летописи уже в легкоузнаваемом состоянии, кроме головоногих- их ранние представители отличаются от моноплакофор только наличием сифона. Брюхоногие, видимо, тоже произошли от моноплакофор. Для тех, кто считает моллюсков потомками кольчатых червей, моноплакофоры с их сериальными жабрами и мышцами ноги выглядят самым примитивным классом. До 1957 года моноплакофоры были известны только в ископаемом состоянии. Напротив, присутствие ресничек у класса Neomeniomorpha заставляет видеть предков моллюсков в плоских червях. Они, как и Chaetodermomorpha, в ископаемом состоянии неизвестны. Головоногие, как уже говорилось, изменились наиболее сильно. Среди них наиболее примитивны Nautiloidea. Их ранние представители имели прямую или слабоизогнутую раковину (уже с сифоном и септами). Многие из них только ползали по дну, другие и ползали и плавали. Они прекрасно подходят на роль поедателей трилобитов, поскольку при плавании та часть раковины, из которой торчало само животное, была обращена ко дну. Ордовикские, силурийские и девонские наутилоидеи демонстрируют неполное сворачивание раковины в спираль. Большинство наутилоидей, кроме отряда Nautilida, вымерло в конце палеозоя. В девоне появился подкласс Ammonoidea, у представителей которого раковина имела складчатые септы. Ранние формы были спирально- плоскостными, но появившиеся в карбоне ползающие представители стали шаровидными. Некоторые аммониты имели парные крышечки на устье- как качающиеся двери. Среди аммонитов были животные с раковиной очень причудливых форм. Их образ жизни совершенно непонятен. Некоторые из них явно не смогли бы и ползать, а разве что лишь пассивно дрейфовать. Об образе жизни других аммонитов известно довольно много. Все они были морские. Не жили возле берега в мелкой, содержащей песок и хорошо прогретой воде, избегали обширных мелководий (куда заходили динозавры), избегали рифов в беспокойных водах, наибольшего разнообразия достигали на глубине 30 – 180 м, служили пищей рыбам, гигантским крабам и мозазаврам; жили стаями. Челюсти их не были приспособлены для мощных укусов. Одни из них питались планктоном, другие бентосом, падалью, растениями и морскими лилиями, и лишь некоторые- другими аммонитами. Вымерли аммониты в конце мела.


В то время как Ammonoidea совершенствовали свою раковину, чтобы сэкономить силы на поддержание плавучести, представители подкласса Coleoidea редуцировали раковину, чтобы плавать быстрее. Среди них белемниты ещё сохраняли в раковине гидростатические камеры и сифон. Позади раковины был длинный ростр, твёрдый балласт, который стабилизировал тело и не давал ему сгибаться от встречного тока воды. Это основное, что остаётся от белемнитов. На многих рострах видны смещения слоёв- результаты столкновений при стремительном плавании. Белемниты появились в карбоне, изобиловали с триаса по мел и очень редко встречаются в эоцене. После исчезновения белемнитов их место заняли кальмары, появившиеся в ранней юре. Вымирание же аммонитов, возможно, было связано с завоеванием моря костистыми рыбами. Безраковинные головоногие, нечувствительные к давлению воды, избежали конкуренции с обладателями плавательного пузыря в глубинах моря. Иглокожие, как и многие другие типы, значительно эволюционировали в течение кембрия и ордовика. Разнообразие их возрастало с кембрия до карбона, сильно снизилось в триасе и опять увеличилось в третичном периоде. На протяжении палеозоя среди иглокожих доминировали стебельчатые. Большая часть их классов давно вымерла. Eocrynoidea были мелкими (6 -7 см), имели конусообразный стебель и неветвящиеся короткие ручки; вымерли в силуре. Cystoidea имели шаровидное или яйцевидное тело, стебель был не у всех. На табличках чашечки, защищавшей тело, были поры для водообмена. Большинство прикреплённые, но были и подвижные, с гибким стеблем без якоря. Blastoidea тоже имели дыхательные поры, а их чашечка была в виде маленького бутона. У Crynoidea поры сконцентрированы на мадрепоровой пластинке, руки ветвящиеся, обычно длинные. Из стебельчатых лишь они живы и поныне. Asteroidea- это перевёрнутые ртом вниз Crynoidea, утратившие стебель. Как ни странно, в ископаемом состоянии они редки, хотя известны с ордовика, как и родственные им Ophyuroidea. Нижнепалеозойские Echinoidea были сигарообразными; в ходе эволюции округлялись и уплощались и упрочняли панцирь. Holothuroidea вторично билатеральны, так как лежат вдоль своей кишки. Они утратили скелет, кроме крошечных спикул. Среди иглокожих особое место занимают Homalozoa. Они не обладали не только радиальной, но часто и какой- либо другой симметрией, хотя некоторым присуща билатеральная. Среди них представители отряда Homostelea состояли из теки (тела) с плоской нижней половиной и выпуклой верхней и стелы (хвоста), и отдалённо напоминали закованного в броню головастика. Были ли они подвижны, неизвестно. Stylophora имели стебель и тело в виде ботинка с подобием жаберных щелей возле носка. Homoiostelea похожи на Homostelea, но имели одну руку, в отличие от двух у последних. Гомалозои, существовавшие с кембрия по ранний девон, некоторыми считаются за предков хордовых, но они слишком плохо изучены, чтобы точно что- то утверждать. С кембрия по карбон известны, а в ордовике наиболее разнообразны граптолиты - тонкие, стержневидные, колониальные животные, родственники современных полухордовых. Очень немногие из них были ползающими, но большинство строило кустистые колонии с веточками, растущими вверх. Были и плавучие - крепившиеся к плавающим предметам или с собственными воздухоносными пузырями; их ветви росли вниз.
Лекция 2 (Ископаемые беспозвоночные ).ppt