

Ядро Геном2013 НВ.ppt
- Количество слайдов: 67
 Лекция 2. Геном. Реализация генетической информации Лекция проф. Немцевой Н. В. , кафедра биологии Ор. ГМА
Лекция 2. Геном. Реализация генетической информации Лекция проф. Немцевой Н. В. , кафедра биологии Ор. ГМА
 Геном — это совокупность всей генетической информации организма. Информация заложена в его полный хромосомный набор и в различные внехромосомные элементы.
Геном — это совокупность всей генетической информации организма. Информация заложена в его полный хромосомный набор и в различные внехромосомные элементы.
 Термин «геном» впервые был введен в двадцатые годы 20 столетия немецким ботаником Гансом Винклером (Hans Karl Albert Winkler) для обозначения генетического материала, заключенного в гаплоидном наборе хромосом организмов одного биологического вида.
Термин «геном» впервые был введен в двадцатые годы 20 столетия немецким ботаником Гансом Винклером (Hans Karl Albert Winkler) для обозначения генетического материала, заключенного в гаплоидном наборе хромосом организмов одного биологического вида.
 Отличия структуры геномов про- и эукариот 1 Наличие ядра 2 Количество хромосом и их организация 3 Форма молекул ДНК 4 Наличие плазмид 5 Компактность генетического материала 6 Наличие белков, связанных с ДНК в составе хромосом 7 Функциональная организация
Отличия структуры геномов про- и эукариот 1 Наличие ядра 2 Количество хромосом и их организация 3 Форма молекул ДНК 4 Наличие плазмид 5 Компактность генетического материала 6 Наличие белков, связанных с ДНК в составе хромосом 7 Функциональная организация
 Генетическая информация") Отличия структуры геномов про- и эукариот 1 Наличие ядра Генетический материал (ДНК) Генетическая информация содержится в сосредоточен в нуклеоиде ядре, т. е. отделена от цитоплазмы клетки 2 Количество хромосом и их организация У прокариот обычно 1 - 2 разные хромосомы. Относительно малые размеры генома, гаплоидность, проявление в фенотипе практически каждой мутации в сочетании с коротким временем генерации, обусловливающие высокую приспособленность в клетках разных организмов - от 4 до нескольких десятков разных хромосом; значительный объем генома; соматические клетки –диплоидный набор, половые клетки – гаплоидный набор, встречаются полиплоиды (растения)
Отличия структуры геномов про- и эукариот 1 Наличие ядра Генетический материал (ДНК) Генетическая информация содержится в сосредоточен в нуклеоиде ядре, т. е. отделена от цитоплазмы клетки 2 Количество хромосом и их организация У прокариот обычно 1 - 2 разные хромосомы. Относительно малые размеры генома, гаплоидность, проявление в фенотипе практически каждой мутации в сочетании с коротким временем генерации, обусловливающие высокую приспособленность в клетках разных организмов - от 4 до нескольких десятков разных хромосом; значительный объем генома; соматические клетки –диплоидный набор, половые клетки – гаплоидный набор, встречаются полиплоиды (растения)
 Отличия структуры геномов про- и эукариот 3 Форма молекул ДНК имеют линейную и кольцевую форма линейная форму 4 Наличие плазмид наряду с хромосомами в геном входят ДНК плазмид и некоторых умеренных вирусов Наряду с хромосомами в геном входят ДНК митохондрий (хлоропластов), а также в ДНК других органоидов 5 Компактность генетического материала Геном организован очень компактно, непрерывен Значительный объем генома, избыточность, наличие в составе некодирующих участков — интронов (экзон -интронная структура); наряду с уникальными генами встречаются многократно повторяющиеся гены
Отличия структуры геномов про- и эукариот 3 Форма молекул ДНК имеют линейную и кольцевую форма линейная форму 4 Наличие плазмид наряду с хромосомами в геном входят ДНК плазмид и некоторых умеренных вирусов Наряду с хромосомами в геном входят ДНК митохондрий (хлоропластов), а также в ДНК других органоидов 5 Компактность генетического материала Геном организован очень компактно, непрерывен Значительный объем генома, избыточность, наличие в составе некодирующих участков — интронов (экзон -интронная структура); наряду с уникальными генами встречаются многократно повторяющиеся гены
 Отличия структуры геномов про- и эукариот 6 Наличие белков, связанных с ДНК в составе хромосом Отсутствие белков гистонов У эукариот ДНК нековалентно связана с белками - гистонами и негистоновыми 7 Функциональная организация Оперон Домены
Отличия структуры геномов про- и эукариот 6 Наличие белков, связанных с ДНК в составе хромосом Отсутствие белков гистонов У эукариот ДНК нековалентно связана с белками - гистонами и негистоновыми 7 Функциональная организация Оперон Домены
 Таким образом: Под геномом организма понимают суммарную ДНК гаплоидного набора хромосом и каждого из внехромосомных генетических элементов, содержащуюся в отдельной клетке зародышевой линии многоклеточного организма.
Таким образом: Под геномом организма понимают суммарную ДНК гаплоидного набора хромосом и каждого из внехромосомных генетических элементов, содержащуюся в отдельной клетке зародышевой линии многоклеточного организма.
 Основная функция генома - обеспечить жизнедеятельность клеток, тканей и органов и передать информацию о наследственных свойствах организма следующему поколению.
Основная функция генома - обеспечить жизнедеятельность клеток, тканей и органов и передать информацию о наследственных свойствах организма следующему поколению.
элементов") ХАРАКТЕРИСТИКА ГЕНОМА • Видоспецифичность; • Дискретность; • Избыточность; • Наличие мобильных (мигрирующих)элементов
ХАРАКТЕРИСТИКА ГЕНОМА • Видоспецифичность; • Дискретность; • Избыточность; • Наличие мобильных (мигрирующих)элементов
, т. е алгоритм с различной степенью детализации Мозаичное строение (У. Гилберт,") Дискретность (мозаичное строение), т. е алгоритм с различной степенью детализации Мозаичное строение (У. Гилберт, 1976 ), чередование кодирующих (экзонов) и некодирующих (нитронов) последовательностей в пределах единицы транскрипции. Схема нуклеотидной последовательности пре-м PHK гена CDK 4 человека. Большую часть последовательности занимают интроны (показаны серым цветом) -
Дискретность (мозаичное строение), т. е алгоритм с различной степенью детализации Мозаичное строение (У. Гилберт, 1976 ), чередование кодирующих (экзонов) и некодирующих (нитронов) последовательностей в пределах единицы транскрипции. Схема нуклеотидной последовательности пре-м PHK гена CDK 4 человека. Большую часть последовательности занимают интроны (показаны серым цветом) -
 - отсутствие корреляции между физическими размерами генома и сложностью организмов Геном эукариот") Избыточность (С-парадокс) - отсутствие корреляции между физическими размерами генома и сложностью организмов Геном эукариот значительно больше по своим размерам чем геном прокариот
Избыточность (С-парадокс) - отсутствие корреляции между физическими размерами генома и сложностью организмов Геном эукариот значительно больше по своим размерам чем геном прокариот
 внутри генома.") Мигрирующие генетические элементы — отдельные участки ДНК, способные осуществлять собственный перенос (транспозицию) внутри генома. Транспозиция связана со способностью мигрирующих элементов кодировать специфический фермент рекомбинации — транспозазу. Транспозоны состоят из 2000 -25 000 пар нуклеотидов, содержат фрагмент ДНК, несущий специфические гены, и два концевых IS-элемента (ДНК бактерий, умеренных и дефектных вирусов). Вертикальный из поколения в поколение перенос информации (дупликация, инсерция, трансдукция) внутри одного генома. Важная роль в адаптации. Ретранспозоны последовательность ДНК, способная реплецироваться и внедрять одну из копий в новое место генома. Перенос информации по горизонтали. Это онкогены, ретровирусы, фаги, эписомы – активно передают информацию между разными видами организмов. ДНК способна к самовоспроизведению с помощью обратной транскрипции. Важная роль в эволюции.
Мигрирующие генетические элементы — отдельные участки ДНК, способные осуществлять собственный перенос (транспозицию) внутри генома. Транспозиция связана со способностью мигрирующих элементов кодировать специфический фермент рекомбинации — транспозазу. Транспозоны состоят из 2000 -25 000 пар нуклеотидов, содержат фрагмент ДНК, несущий специфические гены, и два концевых IS-элемента (ДНК бактерий, умеренных и дефектных вирусов). Вертикальный из поколения в поколение перенос информации (дупликация, инсерция, трансдукция) внутри одного генома. Важная роль в адаптации. Ретранспозоны последовательность ДНК, способная реплецироваться и внедрять одну из копий в новое место генома. Перенос информации по горизонтали. Это онкогены, ретровирусы, фаги, эписомы – активно передают информацию между разными видами организмов. ДНК способна к самовоспроизведению с помощью обратной транскрипции. Важная роль в эволюции.
 Транскрипция не связана с другими генами Активность") ОРГАНИЗАЦИЯ ГЕНОМА Структурные гены Независимые (уникальные последовательности) Транскрипция не связана с другими генами Активность этих генов регулируется гормонами Тандемные гены(умеренно повторяющиеся гены. Высоко повторяющиеся гены) Кластерные гены Регуляторные гены Группы генов, объединенные в домены общей функции Неспецифические ТАТА-блок; СААТ-блок (область промотора) Специфические Энхансеры-усилители; Инсуляторы-ингибируют транскрипцию; Сайленсеры-помошники Регуляция транскрипции через белки, кодируемые этими генами
ОРГАНИЗАЦИЯ ГЕНОМА Структурные гены Независимые (уникальные последовательности) Транскрипция не связана с другими генами Активность этих генов регулируется гормонами Тандемные гены(умеренно повторяющиеся гены. Высоко повторяющиеся гены) Кластерные гены Регуляторные гены Группы генов, объединенные в домены общей функции Неспецифические ТАТА-блок; СААТ-блок (область промотора) Специфические Энхансеры-усилители; Инсуляторы-ингибируют транскрипцию; Сайленсеры-помошники Регуляция транскрипции через белки, кодируемые этими генами
 Высоко повторяющиеся последовательности Выше 105 Это функционально и структурно обособленная ДНК, сателлитная ДНК, состоящая из коротких тандемных повторов 1 -20 п. о. , составляющих джлинные блоки. Структурная функция. (Центромеры, теломеры хромосом) Умеренно повторяющиеся последовательности 10 - 10 4 копий на геном Гены домашнего хозяйства, отвечающие за выполенине жизненно важных функций. Рассеянные и кластерные гены Уникальные последовательности 15 – 98% генома ДНК интронов, спейсерная ДНК и т. п.
Высоко повторяющиеся последовательности Выше 105 Это функционально и структурно обособленная ДНК, сателлитная ДНК, состоящая из коротких тандемных повторов 1 -20 п. о. , составляющих джлинные блоки. Структурная функция. (Центромеры, теломеры хромосом) Умеренно повторяющиеся последовательности 10 - 10 4 копий на геном Гены домашнего хозяйства, отвечающие за выполенине жизненно важных функций. Рассеянные и кластерные гены Уникальные последовательности 15 – 98% генома ДНК интронов, спейсерная ДНК и т. п.
 Все повторы можно подразделить на тандемно повторяющиеся, т. е. расположенные друг за другом «голова к хвосту» , и диспергированные по геному. Тандемные повторы встречаются между генами и в нитронах, но больше всего их в центромерных и теломерных районах хромосом.
Все повторы можно подразделить на тандемно повторяющиеся, т. е. расположенные друг за другом «голова к хвосту» , и диспергированные по геному. Тандемные повторы встречаются между генами и в нитронах, но больше всего их в центромерных и теломерных районах хромосом.
 У человека высокоповторяющаяся ДНК составляет примерно 15% всей ДНК, среднеповторяюшаяся — 8% генома, слабоповторяюшиеся фрагменты (десятки и сотни копий) - около 4, 4% генетического материала человека (эти цифры не совпадают у разных авторов и по мере дальнейших исследований могут быть изменены). Роль повторяющихся элементов генома может быть различной: • структурное значение (например, теломерные и альфоидные повторы); • функциональное значение (повторяющиеся гены р. РНК обеспечивают более высокий уровень синтеза продукта)
У человека высокоповторяющаяся ДНК составляет примерно 15% всей ДНК, среднеповторяюшаяся — 8% генома, слабоповторяюшиеся фрагменты (десятки и сотни копий) - около 4, 4% генетического материала человека (эти цифры не совпадают у разных авторов и по мере дальнейших исследований могут быть изменены). Роль повторяющихся элементов генома может быть различной: • структурное значение (например, теломерные и альфоидные повторы); • функциональное значение (повторяющиеся гены р. РНК обеспечивают более высокий уровень синтеза продукта)
 — международный научноисследовательский") Проект по расшифровке генома человека (англ. The Human Genome Project, HGP) — международный научноисследовательский проект, главной целью которого было определить последовательность нуклеотидов, которые составляют ДНК и идентифицировать 20, 000 -25, 000 генов в человеческом геноме. Проект начался в 1990 году, под руководством Джеймса Уотсона под эгидой Национальной организации здравоохранения США (англ. ). В 2000 году был выпущен рабочий черновик структуры генома, полный геном - в 2003, однако и сегодня дополнительный анализ некоторых участков ещё не закончен.
Проект по расшифровке генома человека (англ. The Human Genome Project, HGP) — международный научноисследовательский проект, главной целью которого было определить последовательность нуклеотидов, которые составляют ДНК и идентифицировать 20, 000 -25, 000 генов в человеческом геноме. Проект начался в 1990 году, под руководством Джеймса Уотсона под эгидой Национальной организации здравоохранения США (англ. ). В 2000 году был выпущен рабочий черновик структуры генома, полный геном - в 2003, однако и сегодня дополнительный анализ некоторых участков ещё не закончен.
 Генетическое картирование - это картирование, основанное на методах классической генетики - определении групп сцепления, частоты рекомбинации и построении генетических карт, где единицей измерения служат проценты рекомбинации, или сантиморганы (с. М). Цитогенетическое картирование осуществляется с применением методов цитогенетики, когда для локализации каких-либо нуклеотидных последовательностей и определения их взаимного расположения используются цитологические препараты. Физическое картирование - это обширная группа методов, позволяющая строить карты генома (обычно их называют физическими) высокого уровня разрешения и определять расстояния между локализуемыми нуклеотидными последовательностями с точностью от нескольких десятков тысяч п. н. до одной нуклеотидной пары.
Генетическое картирование - это картирование, основанное на методах классической генетики - определении групп сцепления, частоты рекомбинации и построении генетических карт, где единицей измерения служат проценты рекомбинации, или сантиморганы (с. М). Цитогенетическое картирование осуществляется с применением методов цитогенетики, когда для локализации каких-либо нуклеотидных последовательностей и определения их взаимного расположения используются цитологические препараты. Физическое картирование - это обширная группа методов, позволяющая строить карты генома (обычно их называют физическими) высокого уровня разрешения и определять расстояния между локализуемыми нуклеотидными последовательностями с точностью от нескольких десятков тысяч п. н. до одной нуклеотидной пары.
 Ген— структурная и функциональная единица наследственности живых организмов. Ген представляет собой участок ДНК, задающий последовательность определённого полипептида либо функциональной РНК.
Ген— структурная и функциональная единица наследственности живых организмов. Ген представляет собой участок ДНК, задающий последовательность определённого полипептида либо функциональной РНК.
 Центральная догма биологии
Центральная догма биологии
 Репликация ДНК
Репликация ДНК
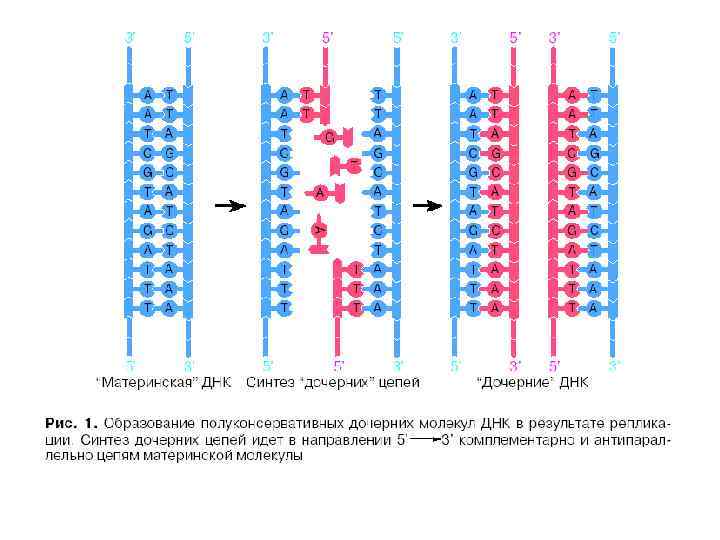
 В основе репликации лежит несколько принципов: • Преемственность генетического материала • Прерывистость • Комплементарность • Полуконсервативность • Униполярность
В основе репликации лежит несколько принципов: • Преемственность генетического материала • Прерывистость • Комплементарность • Полуконсервативность • Униполярность
 Репликация. Показано образование репликонов у эукариот в сравнении с прокариотами
Репликация. Показано образование репликонов у эукариот в сравнении с прокариотами
 Условия, необходимые для репликации ДНК: 1. Набор дезоксирибонуклеозидтрифосфатов (д. АТФ, д. ГТФ, д. ЦТФ, д. ТТФ). 2. 2. Образование РНК – «затравки» (праймера) 3. Расплетенные участки двойной спирали ДНК 4. Необходимые ферменты (геликаза; ДНК топоизомеразы I, III (соответсвуют Е-гиразе); праймаза; ДНК полимеразы I, III; ДНК лигазы
Условия, необходимые для репликации ДНК: 1. Набор дезоксирибонуклеозидтрифосфатов (д. АТФ, д. ГТФ, д. ЦТФ, д. ТТФ). 2. 2. Образование РНК – «затравки» (праймера) 3. Расплетенные участки двойной спирали ДНК 4. Необходимые ферменты (геликаза; ДНК топоизомеразы I, III (соответсвуют Е-гиразе); праймаза; ДНК полимеразы I, III; ДНК лигазы
 Процесс репликации ДНК включает в себя 3 основных этапа: 1. Инициация 2. Элонгация 3. Терминация
Процесс репликации ДНК включает в себя 3 основных этапа: 1. Инициация 2. Элонгация 3. Терминация
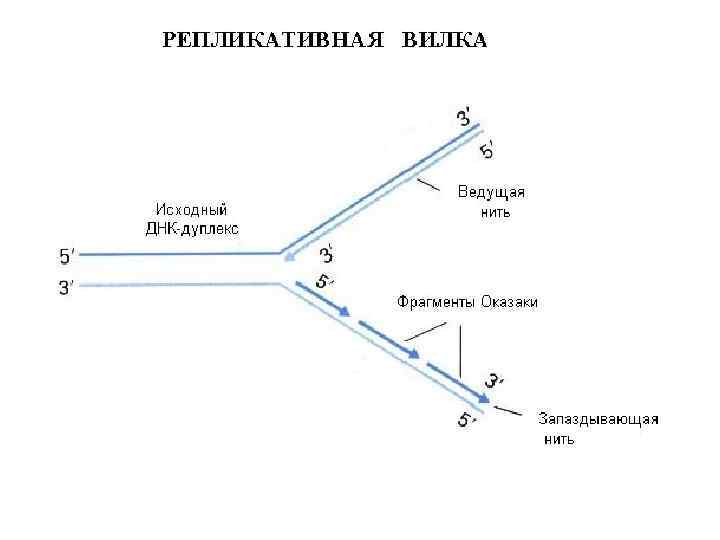
 Схема синтеза ДНК в репликативной вилке
Схема синтеза ДНК в репликативной вилке


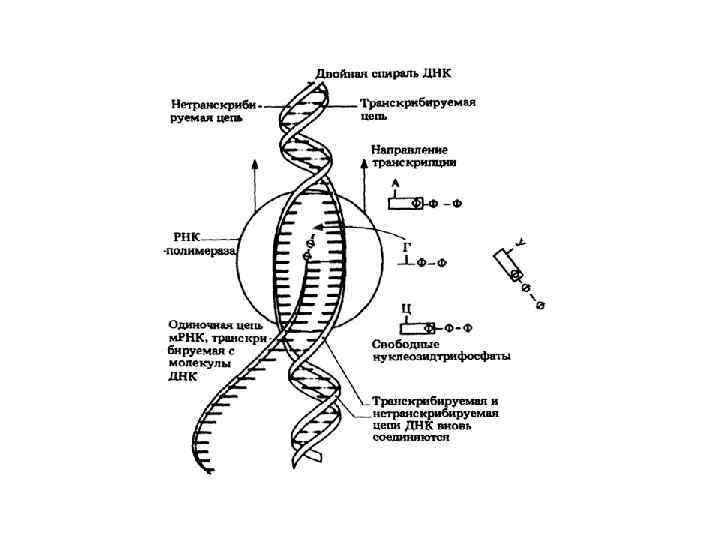
 Транскрипция
Транскрипция
 Условия, необходимые для транскрипции: 1. Наличие транскриптона 2. Рибонуклеозидтрифосфаты: n. АТФ; n. УТФ; n. ЦТФ; n. ГТФ 3. ТАТА-фактор – белковый фактор, который активирует промотор, присоединяясь к специфической последовательности нуклеотидов промотора ТАТА. Это облегчает взаимодействие промотора с РНК-полимеразой 4. Факторы инициации А и В, которые инициируют раскручивание примерно одного витка двойной спирали ДНК 5. Факторы элонгации (E, H, F) повышают активность РНКполимеразы и облегчают локальное расхождение нуклеотидных цепей 6. Белковые факторы транскрипции – это (сигма-факторы: белки shl, CTF, B-белок) для распознавания промоторных участков РНК-полимеразой.
Условия, необходимые для транскрипции: 1. Наличие транскриптона 2. Рибонуклеозидтрифосфаты: n. АТФ; n. УТФ; n. ЦТФ; n. ГТФ 3. ТАТА-фактор – белковый фактор, который активирует промотор, присоединяясь к специфической последовательности нуклеотидов промотора ТАТА. Это облегчает взаимодействие промотора с РНК-полимеразой 4. Факторы инициации А и В, которые инициируют раскручивание примерно одного витка двойной спирали ДНК 5. Факторы элонгации (E, H, F) повышают активность РНКполимеразы и облегчают локальное расхождение нуклеотидных цепей 6. Белковые факторы транскрипции – это (сигма-факторы: белки shl, CTF, B-белок) для распознавания промоторных участков РНК-полимеразой.
 7. Фермент – ДНК-зависимая РНК-полимераза I, II. III 8. Фермент поли А-полимераза 9. Сплайсосомы (рибозимы) 10. Mn++ и Mg ++
7. Фермент – ДНК-зависимая РНК-полимераза I, II. III 8. Фермент поли А-полимераза 9. Сплайсосомы (рибозимы) 10. Mn++ и Mg ++
 Процесс транскрипции включает в себя 3 основных этапа: 1. Инициация 2. Элонгация 3. Терминация
Процесс транскрипции включает в себя 3 основных этапа: 1. Инициация 2. Элонгация 3. Терминация
 -полидромный участок ДНК, разделяющий транскриптоны, образуя так называемые «шпильки»") Транскриптон Спенсерный сайт рестрикции (ССР) -полидромный участок ДНК, разделяющий транскриптоны, образуя так называемые «шпильки» в ДНК. Состоит из инвертированных нуклеотидов (чаще гуанин и цитозин) по принципу «КАЗАК» - Разделение транскриптонов Промотор -ЦААТ блок –активный участок, состоящий их 70 -80 -100 пар нуклеотидов и заканчивается ЦААТ - Узнавание РНК-полимеразы ТАТА блок (блок Хогнесса) – состоит из 30 пар нуклеотидов, обогащен последовательностями аденина и тимина - Присоединение РНК-полимеразы Сайт инициации транскрипции - который при трансляции будет соответствовать АК – метионин (ТАЦ на ДНК) - Точка инициации, стартовая точка Оператор - с него начинается синтез и-РНК и с ним взаимодействует особый белок репрессор или индуктор и - от этого будет зависеть будет или нет идти транскипция Структурный блок – экзоны, интроны, донорные сайты сплайсинга Терминатор - Поли А
Транскриптон Спенсерный сайт рестрикции (ССР) -полидромный участок ДНК, разделяющий транскриптоны, образуя так называемые «шпильки» в ДНК. Состоит из инвертированных нуклеотидов (чаще гуанин и цитозин) по принципу «КАЗАК» - Разделение транскриптонов Промотор -ЦААТ блок –активный участок, состоящий их 70 -80 -100 пар нуклеотидов и заканчивается ЦААТ - Узнавание РНК-полимеразы ТАТА блок (блок Хогнесса) – состоит из 30 пар нуклеотидов, обогащен последовательностями аденина и тимина - Присоединение РНК-полимеразы Сайт инициации транскрипции - который при трансляции будет соответствовать АК – метионин (ТАЦ на ДНК) - Точка инициации, стартовая точка Оператор - с него начинается синтез и-РНК и с ним взаимодействует особый белок репрессор или индуктор и - от этого будет зависеть будет или нет идти транскипция Структурный блок – экзоны, интроны, донорные сайты сплайсинга Терминатор - Поли А
 Инициация транскрипции — сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариот также и от более далеких участков генома — энхансеров и сайленсеров) и от наличия или отсутствия различных белковых факторов.
Инициация транскрипции — сложный процесс, зависящий от последовательности ДНК вблизи транскрибируемой последовательности (а у эукариот также и от более далеких участков генома — энхансеров и сайленсеров) и от наличия или отсутствия различных белковых факторов.
 Элонгация, терминация Белковые факторы элонгации обеспечивают продвижение РНК полимеразы со скоростью 40 -50 п. н. в секунду в направлении от 3’ к 5’ концу РНК-полимераза остановится, когда достигнет терминирующих кодонов. С помощью ρ-фактора (греч. ро - фактор терминации) РНК полимераза и первичный транскрипт отделяются от ДНК
Элонгация, терминация Белковые факторы элонгации обеспечивают продвижение РНК полимеразы со скоростью 40 -50 п. н. в секунду в направлении от 3’ к 5’ концу РНК-полимераза остановится, когда достигнет терминирующих кодонов. С помощью ρ-фактора (греч. ро - фактор терминации) РНК полимераза и первичный транскрипт отделяются от ДНК
 Пострепликативный процесс
Пострепликативный процесс
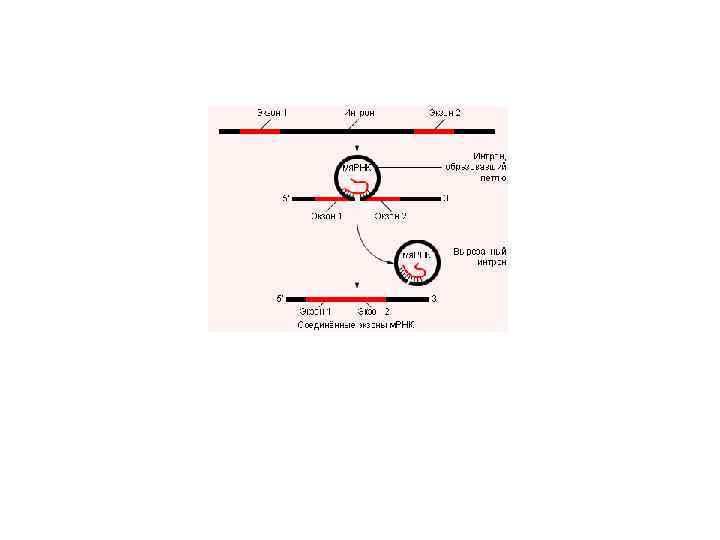
 сплайсинга, при котором из одного и того") В настоящее время доказана возможность альтернативного (взаимоисключающего) сплайсинга, при котором из одного и того же первичного транскрипта могут удаляться разные нуклеотидные последовательности и образовываться разные зрелые м. РНК. В результате одна и та же последовательность нуклеотидов ДНК может служить информацией для синтеза разных пептидов.
В настоящее время доказана возможность альтернативного (взаимоисключающего) сплайсинга, при котором из одного и того же первичного транскрипта могут удаляться разные нуклеотидные последовательности и образовываться разные зрелые м. РНК. В результате одна и та же последовательность нуклеотидов ДНК может служить информацией для синтеза разных пептидов.
 Кепирование Кэпирование 5’-конца транскрипта 7’-метил ГТФ, защищает от 5’-экзонуклеазы, защита 5’ м. РНК от экзонуклеаз, правильная установка РНК на рибосоме при взаимодействии белков с кэпом
Кепирование Кэпирование 5’-конца транскрипта 7’-метил ГТФ, защищает от 5’-экзонуклеазы, защита 5’ м. РНК от экзонуклеаз, правильная установка РНК на рибосоме при взаимодействии белков с кэпом
 Полиаденилирование После синтеза про-m. РНК на расстоянии примерно 20 н в направлении 3'-конца от последовательности 5'-AAУAA-3' происходит разрезание эндонуклеазой и к новому 3'-концу присоединяется от 30 до 300 остатков АМФ (безматричный синтез). Поли. А хвост защищает 3'-конец от гидролиза, т. к. покрыт поли. Асвязывающими белками. Время жизни m. РНК зависит от длинны поли. А-хвоста.
Полиаденилирование После синтеза про-m. РНК на расстоянии примерно 20 н в направлении 3'-конца от последовательности 5'-AAУAA-3' происходит разрезание эндонуклеазой и к новому 3'-концу присоединяется от 30 до 300 остатков АМФ (безматричный синтез). Поли. А хвост защищает 3'-конец от гидролиза, т. к. покрыт поли. Асвязывающими белками. Время жизни m. РНК зависит от длинны поли. А-хвоста.

") Трансляция (Биосинтез белка)
Трансляция (Биосинтез белка)
 ГЕНЕТИЧЕСКИЙ КОД
ГЕНЕТИЧЕСКИЙ КОД
 СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА • Триплетность • Специфичность • Универсальность • Вырожденность • Неперекрывающийся • Колинеарность
СВОЙСТВА ГЕНЕТИЧЕСКОГО КОДА • Триплетность • Специфичность • Универсальность • Вырожденность • Неперекрывающийся • Колинеарность
 Процесс трансляции включает в себя 3 основных этапа: 1. Инициация 2. Элонгация 3. Терминация
Процесс трансляции включает в себя 3 основных этапа: 1. Инициация 2. Элонгация 3. Терминация
 факторы инициации (их 3: IF-1 -IF-3) обеспечивают объединение рибосомы с и-РНК, отвечают за") а) факторы инициации (их 3: IF-1 -IF-3) обеспечивают объединение рибосомы с и-РНК, отвечают за взаимодействие кодона AUG на рибосоме со специальной инициирующей т. РНК; б) факторы элонгации (их тоже 3: EF-Tu, EF-Ts, EF-G) обеспечивают связывание антикодонов заряженных т-РНК с соответствующими кодонами и-РНК на рибосоме и продвижение рибосомы вдоль матрицы и-РНК; в) факторы терминации: считается, что не существует т-РНК, специфичных для трех кодонов-терминаторов (UAA, UAG, UGA), их опознают и с ними взаимодействуют специальные белки (RF 1; RF 2; RF 3); при этом RF 1 узнает UAA и UAG; RF 2 - UAA и UGA; а RF 3 не имеет кодоновой специфичности и взаимодействует с RF 1 и RF 2, делая процесс терминации более эффективным и зависимым от энергии расщепления ГТФ. Аппарат трансляции у эукариот несколько сложнее: белков в рибосоме больше, и факторов трансляции больше. Например одних факторов инициации не меньше десяти, тем не менее общая схема процесса трансляции принципиально такая же, как и у бактерий.
а) факторы инициации (их 3: IF-1 -IF-3) обеспечивают объединение рибосомы с и-РНК, отвечают за взаимодействие кодона AUG на рибосоме со специальной инициирующей т. РНК; б) факторы элонгации (их тоже 3: EF-Tu, EF-Ts, EF-G) обеспечивают связывание антикодонов заряженных т-РНК с соответствующими кодонами и-РНК на рибосоме и продвижение рибосомы вдоль матрицы и-РНК; в) факторы терминации: считается, что не существует т-РНК, специфичных для трех кодонов-терминаторов (UAA, UAG, UGA), их опознают и с ними взаимодействуют специальные белки (RF 1; RF 2; RF 3); при этом RF 1 узнает UAA и UAG; RF 2 - UAA и UGA; а RF 3 не имеет кодоновой специфичности и взаимодействует с RF 1 и RF 2, делая процесс терминации более эффективным и зависимым от энергии расщепления ГТФ. Аппарат трансляции у эукариот несколько сложнее: белков в рибосоме больше, и факторов трансляции больше. Например одних факторов инициации не меньше десяти, тем не менее общая схема процесса трансляции принципиально такая же, как и у бактерий.



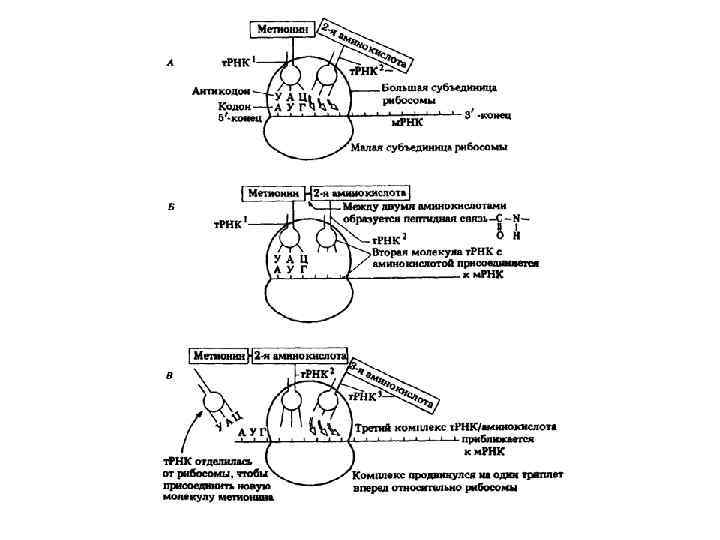
") Инициация Объединение двух субъединиц рибосом на определенном участке м. РНК (с инициирующим кодоном АУГ) и присоединение к ней первой аминоацил т-РНК с АК- метионин, которая является затравочной, и располагается в пептидильном участке рибосомы. Рибосома захлопывается и делает «шаг» . К концу инициации в пептидильном участке – АК-метионин, а в аминоацильном – следующая т-РНК.
Инициация Объединение двух субъединиц рибосом на определенном участке м. РНК (с инициирующим кодоном АУГ) и присоединение к ней первой аминоацил т-РНК с АК- метионин, которая является затравочной, и располагается в пептидильном участке рибосомы. Рибосома захлопывается и делает «шаг» . К концу инициации в пептидильном участке – АК-метионин, а в аминоацильном – следующая т-РНК.
.") Элонгация Удлинение по принципу колинеарности (перевод «языка» последовательности триплетов нуклеотидов на «язык» последовательности аминокислот). Пептидильный и аминоацильный участки рибосомы находятся очень близко, поэтому между двумя АК, расположенными в них образуется пептидная связь под действием пептидилтрансферазы.
Элонгация Удлинение по принципу колинеарности (перевод «языка» последовательности триплетов нуклеотидов на «язык» последовательности аминокислот). Пептидильный и аминоацильный участки рибосомы находятся очень близко, поэтому между двумя АК, расположенными в них образуется пептидная связь под действием пептидилтрансферазы.
 Непосредственная функция отдельного гена состоит в кодировании структуры определенного белка-фермента, который катализирует одну биохимическую реакцию, протекающую в определенных условиях среды. Ген (участок ДНК) → и. РНК → белок-фермент → биохимическая реакция → наследственный признак.
Непосредственная функция отдельного гена состоит в кодировании структуры определенного белка-фермента, который катализирует одну биохимическую реакцию, протекающую в определенных условиях среды. Ген (участок ДНК) → и. РНК → белок-фермент → биохимическая реакция → наследственный признак.
, который входит в акцепторный") Терминация Весь процесс идет до терминального кодона (УАА, УАГ, УГА), который входит в акцепторный участок рибосомы. К последней АК присоединяется вода, ее капрбоксильный конец отделяется от т. РНК и связь с рибосомой теряется, рибосома распадается на 2 субъединицы.
Терминация Весь процесс идет до терминального кодона (УАА, УАГ, УГА), который входит в акцепторный участок рибосомы. К последней АК присоединяется вода, ее капрбоксильный конец отделяется от т. РНК и связь с рибосомой теряется, рибосома распадается на 2 субъединицы.
 Модификация Образовавшийся первичный белок через ЭПС проходит в аппарат Гольджи, где осуществляется его модификация. Готовый белок покрывается мембраной и используется для собственных нужд клетки, либо экспортируется а пределы клетки.
Модификация Образовавшийся первичный белок через ЭПС проходит в аппарат Гольджи, где осуществляется его модификация. Готовый белок покрывается мембраной и используется для собственных нужд клетки, либо экспортируется а пределы клетки.
 Обобщенная схема процесса экспрессии генетической информации в эукариотической клетке
Обобщенная схема процесса экспрессии генетической информации в эукариотической клетке
 РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ ПРОКАРИОТ Негативный контроль – репрессор кодируется регуляторным геном и взаимодействуя с оператором регулирует транскрипцию. Позитивный контроль – регуляторный белок как индуктор присоединяется к оператору, обеспечивая соединение РНКполимеразы с промотором
РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ ПРОКАРИОТ Негативный контроль – репрессор кодируется регуляторным геном и взаимодействуя с оператором регулирует транскрипцию. Позитивный контроль – регуляторный белок как индуктор присоединяется к оператору, обеспечивая соединение РНКполимеразы с промотором
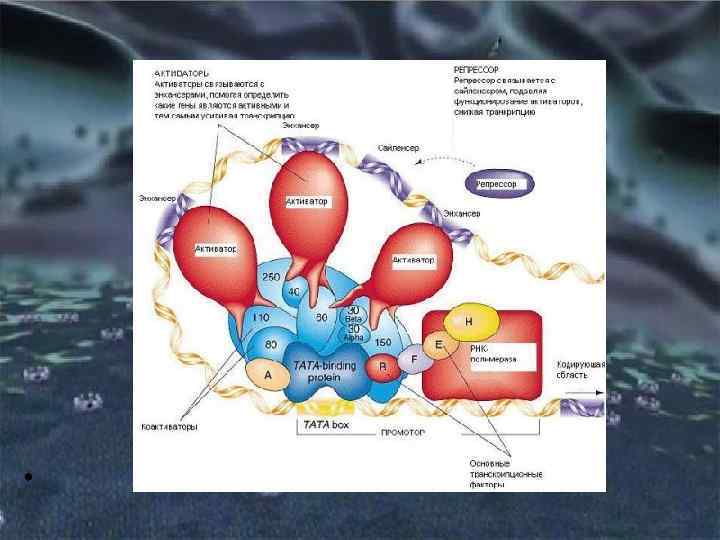
 Общие принципы генетического контроля экспрессии генов С участием белка-репрессора С участием белка-активатора
Общие принципы генетического контроля экспрессии генов С участием белка-репрессора С участием белка-активатора
 Если белок-регулятор взаимодействует с оператором, занимающим часть промотора или расположенным между ним и структурной частью гена, то это не дает возможности РНК-полимеразе соединиться с промоторной последовательностью и осуществить транскрипцию. Такой белок называют репрессором, и в этом случае осуществляется негативный контроль экспрессии гена со стороны генарегулятора Если промотор обладает слабой способностью соединяться с РНКполимеразой, а ему предшествует область, узнаваемая белкомрегулятором, присоединение последнего непосредственно перед промотором к молекуле ДНК облегчает связывание РНКполимеразы с промотором, вслед за чем следует транскрипция. Такие белки называют активаторами (или апоиндукторами), а контроль экспрессии гена со стороны генарегулятора — позитивным
Если белок-регулятор взаимодействует с оператором, занимающим часть промотора или расположенным между ним и структурной частью гена, то это не дает возможности РНК-полимеразе соединиться с промоторной последовательностью и осуществить транскрипцию. Такой белок называют репрессором, и в этом случае осуществляется негативный контроль экспрессии гена со стороны генарегулятора Если промотор обладает слабой способностью соединяться с РНКполимеразой, а ему предшествует область, узнаваемая белкомрегулятором, присоединение последнего непосредственно перед промотором к молекуле ДНК облегчает связывание РНКполимеразы с промотором, вслед за чем следует транскрипция. Такие белки называют активаторами (или апоиндукторами), а контроль экспрессии гена со стороны генарегулятора — позитивным
 Способыпосттрансляционной регуляции активности генов эукариот • увеличение количества продукта гена достигается за счет увеличения числа его копий. Наиболее известный пример амплификации (умножения) многократная избирательная репликация генов р. РНК в ооцитах амфибий. Такой тип регуляции репликации обеспечивает накопление большого количества копий определенных генов и их дальнейшую транскрипцию; • образование вторичных структур в виде шпилек в районе инициирующего кодона, верхняя часть которых - петля, образованная из неспаренных оснований. Кодоны AUG, находясь в районе стебля, блокируют трансляцию. • наличие регуляторных белков, которые, связываясь с инициирующим кодоном, репрессируют трансляцию; • ацетилирование гистонов, приводящее к развороту нуклеосомных частиц, модифицированные таким образом гистоны связываются с ДНК менее эффективно; • метилирование как молекул ДНК, так и гистонов, входящих в структуру нуклеосом. Метилирование усиливает основность гистонов и связь с ДНК. Эти изменения гистонов нарушают процесс деспирализации нуклеосом перед началом синтеза ДН К или ее транскрипции.
Способыпосттрансляционной регуляции активности генов эукариот • увеличение количества продукта гена достигается за счет увеличения числа его копий. Наиболее известный пример амплификации (умножения) многократная избирательная репликация генов р. РНК в ооцитах амфибий. Такой тип регуляции репликации обеспечивает накопление большого количества копий определенных генов и их дальнейшую транскрипцию; • образование вторичных структур в виде шпилек в районе инициирующего кодона, верхняя часть которых - петля, образованная из неспаренных оснований. Кодоны AUG, находясь в районе стебля, блокируют трансляцию. • наличие регуляторных белков, которые, связываясь с инициирующим кодоном, репрессируют трансляцию; • ацетилирование гистонов, приводящее к развороту нуклеосомных частиц, модифицированные таким образом гистоны связываются с ДНК менее эффективно; • метилирование как молекул ДНК, так и гистонов, входящих в структуру нуклеосом. Метилирование усиливает основность гистонов и связь с ДНК. Эти изменения гистонов нарушают процесс деспирализации нуклеосом перед началом синтеза ДН К или ее транскрипции.
 РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ ЭУКАРИОТ
РЕГУЛЯЦИЯ АКТИВНОСТИ ГЕНОВ ЭУКАРИОТ
 Благодарю за внимание
Благодарю за внимание