2 Физиология возбудимых тканей.ppt
- Количество слайдов: 36



Лекция № 2 ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ

Раздражимость – основное свойство живых клеток, способность реагировать в ответ на действие раздражителей изменением структурных и функциональных свойств. Раздражитель – любое изменение внешней или внутренней среды (если достаточно велико, возникло достаточно быстро, продолжается достаточно долго). Классификация раздражителей ( по природе) : - физические (механические, температ. , звуковые, световые, электрические) - химические (щелочи, кислоты, гормоны, продукты обмена) - физико-химические (изменение р. Н среды, ионного состава) Классификация раздражителей ( по степени приспособленности) : - адекватные (к восприятию которых биологическая структура адаптирована, н-р, видимый свет для рецепторов, нервный импульс для скелетной мышцы) - неадекватные (воздействующие на структуру, не приспособленную специально для их восприятия, н-р, электрич. ток для мышечной ткани, механич. удар); их пороговая сила в сотни и более раз превышает пороговую силу адекватного раздражителя В процессе эволюции произошла постепенная дифференциация тканей, которые осуществляют приспособительную деятельность. Раздражимость этих тканей достигла наивысшего развития и трансформировалась в новое свойство – возбудимость. Возбудимость – способность клеток отвечать на действие возбудителя возбуждением.

Возбуждение - сложный биологический процесс, который характеризуется специфическим изменением процессов обмена веществ, теплообразования, временной деполяризацией мембраны клеток и проявляющийся специализированной реакцией ткани (сокращение мышцы, отделение секрета железой и т. д. ) Возбуждение – ответная реакция возбудимой ткани на действие возбудителя, проявляющаяся в совокупности физических, физикохимических, метаболических процессов и изменений деятельности. Возбудимые ткани нервная железистая мышечная Возбуждение (распространяющийся волнообразный процесс) проявляется: в мышечной ткани - сокращением, в железистой – образованием и выделением секрета, в нервной – возникновением и проведением нервного импульса. Различные ткани обладают неодинаковой возбудимостью. Мера возбудимости – порог раздражения – минимальная сила раздражителя, которая способна вызвать возбуждение. Раздражители с силой меньше порога раздражения называются подпороговыми, выше – сверхпороговыми.

Природа возбуждения. Гальвани Луиджи. Впервые продемонстрировал высокую чувствительность возбудимых тканей к действию слабого электрического тока. Показал наличие электрических явлений в живых клетках. Конец 19 в. Герман Л. , Дюбуа-Раймон Э. , Бернштейн Ю. доказали, что электрические явления, возникающие в возбудимых тканях, обусловлены электрическими свойствами клеточных мембран. Дюбуа-Раймон Э. показал, что наружная поверхность мышцы заряжена положительно по отношению к внутреннему содержимому. В состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, кот. называется мембранным потенциалом покоя (МПП) или мембранным потенциалом.

Луи джи Гальва ни ( 9 сентября 1737 - Эмиль Генрих Дюбуа-Реймон (7 ноября 4 декабря 1798) — итальянский врач, 1818 - 26 декабря 1896) — немецкий анатом, физиолог и физик, один из физиолог, швейцарец по происхождению, основателей электрофизиологии и учения об философ, иностранный членэлектричестве, основоположник корреспондент Петербургской АН (1892). экспериментальной электрофизиологии. Основоположник электрофизиолоиги — Первым исследовал электрические явления установил ряд закономерностей, при мышечном сокращении ( «животное характеризующих электрические явления в электричество» ). Обнаружил возникновение мышцах и нервах. Автор молекулярной разности потенциалов при контакте разных теории биопотенциалов. видов металла и электролита.

Потенциал покоя Опыт Ходжкина – Хаксли : введение активного электрода в гигантский (1 мм в диаметре) аксон кальмара, помещенный в морскую воду. Второй электрод во внешней среде (морская вода). В момент введения электрода внутрь аксона регистрировали скачок отрицательного потенциала, т. е. внутренняя среда заряжена отрицательно относительно внешней среды. Факторы, влияющие на величину ПП: 1. Селективность проницаемости клеточной мембраны для различных ионов 2. Концентрация ионов цитоплазмы клетки и ионов окружающей среды 3. Работа механизмов активного транспорта ионов Потенциал покоя (ПП) — мембранный потенциал возбудимой клетки (нейрона, кардиомиоцита) в невозбужденном состоянии. Он представляет собой разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны и составляет у теплокровных от -55 до -100 м. В. У нейронов и нервных волокон обычно составляет -70 м. В.
 отрицательности внутри клетки за счёт")
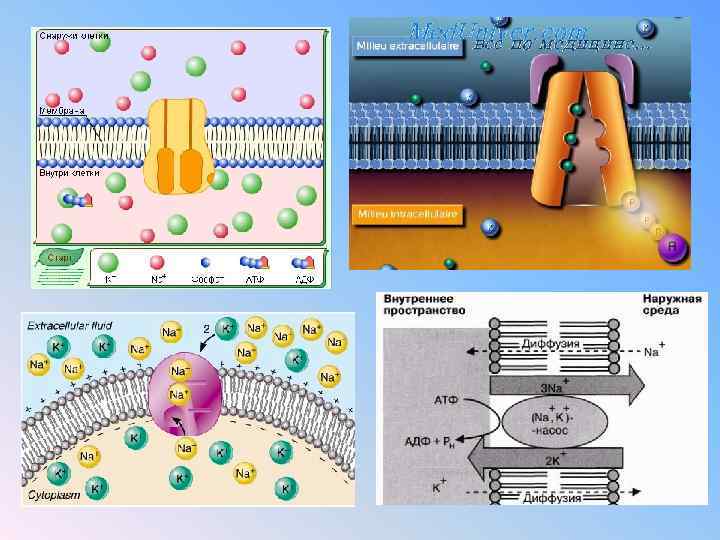
Формирование ПП. Первый этап: создание незначительной (-10 м. В) отрицательности внутри клетки за счёт неравного асимметричного обмена Na+ на K+ в соотношении 3 : 2. В результате этого клетку покидает больше положительных зарядов с натрием, чем возвращается в неё с калием. Такая особенность работы натрий-калиевого насоса, осуществляющего взаимообмен этих ионов через мембрану с затратами энергии АТФ, обеспечивает его электрогенность. Результаты деятельности мембранных ионных насосов-обменников на первом этапе формирования ПП таковы: • 1. Дефицит ионов натрия (Na+) в клетке. • 2. Избыток ионов калия (K+) в клетке. • 3. Появление на мембране слабого электрического потенциала (-10 м. В). Второй этап: создание значительной (-60 м. В) отрицательности внутри клетки за счёт утечки из неё через мембрану ионов K+. Ионы калия K+ покидают клетку и уносят с собой из неё положительные заряды, доводя отрицательность до -70 м. В. Итак, мембранный потенциал покоя - это дефицит положительных электрических зарядов внутри клетки, возникающий за счёт утечки из неё положительных ионов калия и электрогенного действия натрий-калиевого насоса.

В состоянии покоя мембрана клетки высокопроницаема для ионов К, Сl и малопроницаема для ионов Na. Мембраны живых клеток в определенной степени проницаемы для всех ионов. Без специальных механизмов невозможно поддерживать постоянный градиент концентрации. Для этой цели существуют системы активного транспорта, перемещающие ионы против градиента концентрации и работающие с затратой энергии.
 — быстрое колебание потенциала покоя, сопровождающееся, как правило, перезарядкой мембраны.")
Потенциал действия (ПД) — быстрое колебание потенциала покоя, сопровождающееся, как правило, перезарядкой мембраны. ПД — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного и мышечного сокращения.

Этапы процесса: 1. 2. 3. Под влиянием раздражителя пороговой или сверхпороговой величины проницаемость мембраны клетки для ионов возрастает, и они устремляются внутрь клетки (проницаемость Na в клетку в 500 раз больше, и в 20 раз больше, чем для К), что приводит к уменьшению величины мембранного ПП – возникает деполяризация мембраны. Разность потенциалов на мембране исчезает. Происходит перезарядка мембраны (инверсия заряда) – внутренняя поверхность заряжается положительно по отношению к наружной. Потенциал превышения достигает 30 -50 м. В, после чего закрываются быстрые натриевые каналы и открываются калиевые. Начинается процесс восстановления исходного уровня МПП – реполяризация мембраны.

Фазы одиночного цикла возбуждения (при внеклеточном отведении, т. е. с помощью электродов, приложенных к поверхности клетки) 1. Предспайк (предпотенциал) – процесс медленной деполяризации мембраны 2. Пиковый потенциал, или спайк (мембрана не способна отвечать возбуждением на раздражение) 3. Отрицательный следовой потенциал – от критического уровня деполяризации мембраны до исходного уровня поляризации мембраны (порог раздражения снижен, возбудимость повышена) 4. Положительный следовой потенциал – увеличение мембранного потенциала покоя и постепенное возвращение его к исходной величине (порог раздражения повышается).

Фазы одиночного цикла возбуждения (при внутриклеточном отведении, т. е. с помощью электродов, один из которых внутри клетки, а другой на ее поверхности) 1. Местное возбуждение (локальный ответ) – начальная деполяризация мембраны 2. Деполяризация мембраны (восходящая часть спайка, включая инверсию) 3. Реполяризация мембраны – нисходящая часть ПД 4. Следовая деполяризация (соответствует отрицательному следовому потенциалу) 5. Следовая гиперполяризация (соответствует положительному следовому потенциалу) Причины гиперполяризации: 1) продолжается выход ионов К 2) открываются каналы для Cl и Cl поступает в клетку 3) усиливается работа Na-K-насоса

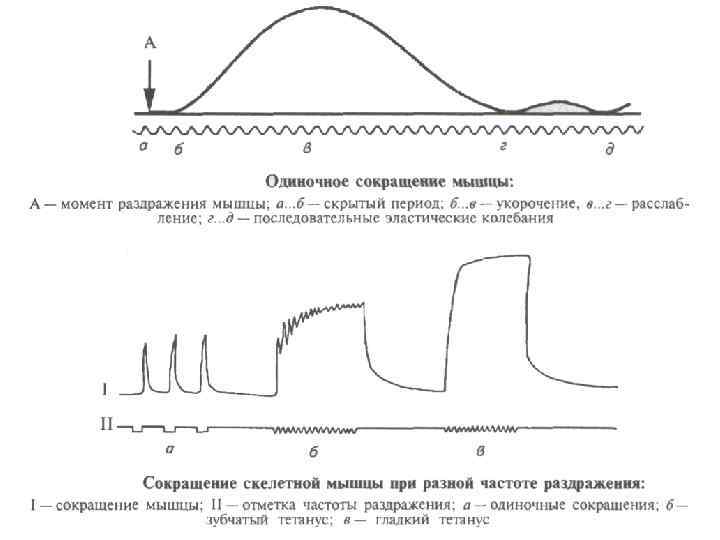
Лабильность Понятие введено Н. Е. Введенским как функциональная подвижность возбудимых тканей. Мера лабильности – количество ПД, которое способна генерировать возбудимая ткань в единицу времени. Например, наиболее лабильными являются волокна слухового нерва, генерация ПД достигает 1000 Гц. Николай Евгеньевич Введенский (16(28) апреля 1852 -16 сентября 1922) — русский физиолог, ученик И. М. Сеченова

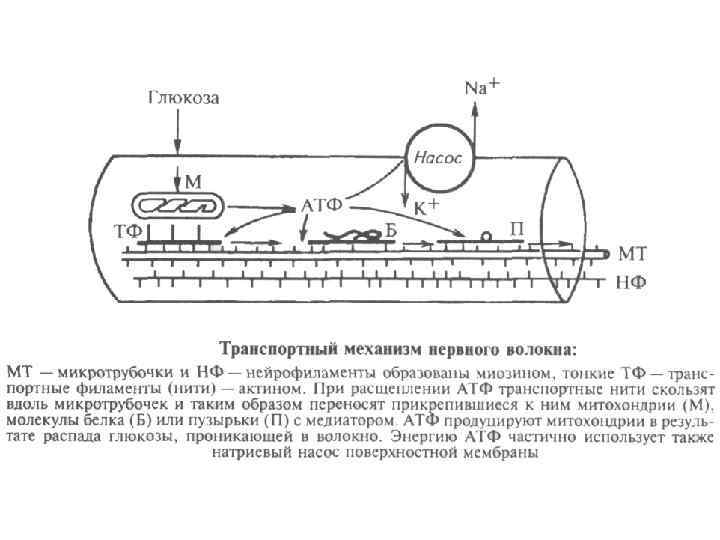
Механизм проведения возбуждения по нервным волокнам Миелиновая оболочка — электроизолирующая оболочка, покрывающая аксоны многих нейронов. Миелиновую оболочку образуют глиальные клетки. Миелиновая оболочка формируется из плоского выроста тела глиальной клетки, многократно оборачивающего аксон подобно изоляционной ленте. Цитоплазма в выросте практически отсутствует, в результате чего миелиновая оболочка представляет собой, по сути, множество слоёв клеточной мембраны. Миелин прерывается только в области перехватов Ранвье, которые встречаются через правильные промежутки длиной примерно 1 мм. В связи с тем, что ионные токи не могут проходить сквозь миелин, вход и выход ионов осуществляется лишь в области перехватов. Это ведёт к увеличению скорости проведения нервного импульса. Нервные волокна - миелинизированные немиелинизированные

Особенности проведения возбуждения по немиелинизированным волокнам 1. 2. 3. 4. 5. Потенциал действия распространяется непрерывно. Возбуждение последовательно охватывает соседние участки мембраны осевого цилиндра и распространяется вдоль оси аксона до его конца. Проведение нервного импульса начинается с распространением электрического поля. Возникший потенциал действия за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые потенциалы. Распространение возбуждения идет с постепенным ослаблением – декрементом, т. к. процессы метаболизма не обеспечивают быстрой компенсации расхода энергии. Сам потенциал действия не перемещается, он исчезает там же, где возник. Главную роль в возникновении нового потенциал действия играет предыдущий. Скорость распространения возбуждения по волокну определяется его диаметром.
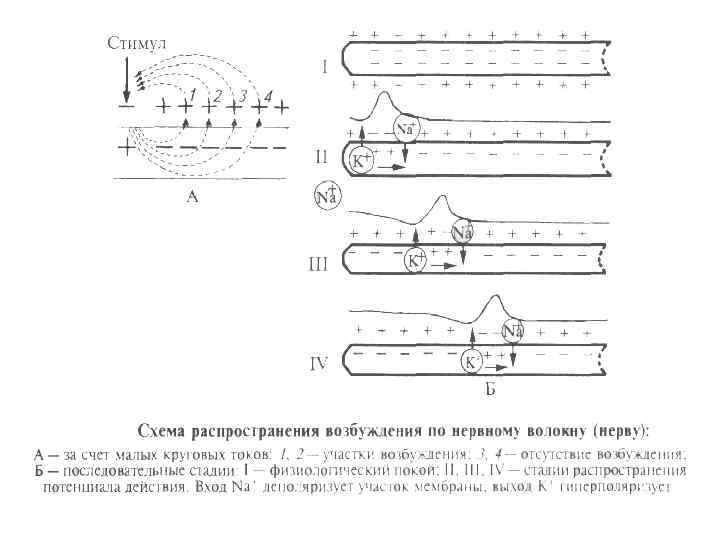

Особенности проведения возбуждения по миелинизированным волокнам 1. 2. 3. 4. 5. Возбуждение происходит не затухая, т. е. бездекрементно. Потенциал действия распространяется скачкообразно (сальтаторное проведение). Потенциал действия, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых потенциал действия. В случае повреждения одного перехвата Ранвье потенциал действия возбуждает 2 -й, 3 -й, 4 -й и даже 5 -й, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Экономичность сальтаторного проведения в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых меньше 1 % мембраны, и, следовательно, необходимо значительно меньше энергии для восстановления трансмембранных градиентов Na+ и K+, расходующихся в результате возникновения потенциал действия, что может иметь значение при высокой частоте разрядов, идущих по нервному волокну. Например, при диаметре волокна около 2 мкм и отсутствии миелиновой оболочки скорость проведения будет составлять ~1 м/с, а при наличии даже слабой миелинизации при том же диаметре волокна — 15 -20 м/с. В волокнах большего диаметра, обладающих толстой миелинововой оболочкой, скорость проведения может достигать 120 м/с. Скорость распространения потенциала действия по мембране отдельно взятого нервного волокна не является постоянной величиной — в зависимости от различных условий, эта скорость может очень значительно уменьшаться и, соответственно, увеличиваться, возвращаясь к некоему исходному уровню.

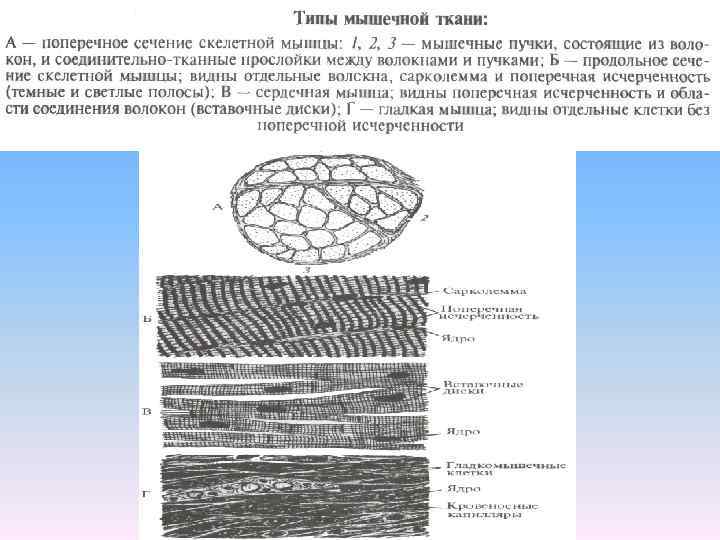
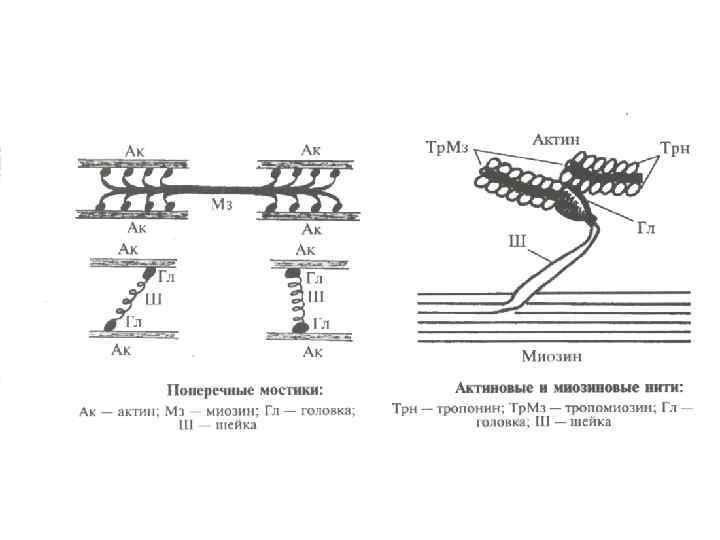
Физиология мышц



Физиология нервов

Нервно-мышечная передача возбуждения. Синапс. утолщенная часть мембраны клетки с которой контактирует пресинаптическая мембрана. Имеет электрически возбудимые ионные каналы и поэтому способна к генерации потенциала действия, а также специальные белковые структуры — рецепторы, воспринимающие действие медиаторов. пространство между пре - и постсинаптическими мембранами от 50 до 500 нм оно заполнено жидкостью, близкой по составу плазме крови.


Синапс - это структурно-физиологическое образование, обеспечивающее передачу возбуждения с нервного волокна на иннервируемую им клетку (мышечную, нервную или железистую). синапс клетки нервное волокно мышечная, нервная, железистая Передача происходит при выделении нервными окончаниями специфических химических соединений — медиаторов (передатчиков) нервного импульса. Медиатор у животных в скелетных мышцах - ацетилхолин.

Синапсы состоят из пресинаптической постсинаптической мембраны щели мембраны утолщенная часть мембраны покрывающей расширенное нервное окончание (нейросекрето рный аппарат). клетки с которой контактирует пресинаптическая мембрана. . Имеет электрически возбудимые ионные каналы и поэтому способна к генерации потенциала действия, а также специальные белковые структуры — рецепторы, воспринимающие действие медиаторов. пространство между пре- и постсинаптиче скими мембранами от 50 до 500 нм, оно заполнено жидкостью, близкой по составу плазме крови.
, достигнув пресинаптической мембраны,")
Возбуждение распространяется по нервному волокну в виде потенциала действия (нервного импульса), достигнув пресинаптической мембраны, вызывает ее деполяризацию, что приводит к открытию кальциевых каналов. Ионы Са 2+ входят внутрь нервного окончания и способствуют освобождению медиатора из синаптических пузырьков и выходу его в синаптическую щель. Медиатор быстро диффундирует через щель и воздействует на постсинаптическую мембрану — взаимодействует с рецептором (ацетилхолин — с холинорецептором, норадреналин — с адренорецептором и т. д. ).

На взаимодействие медиатора с рецептором мембрана отвечает изменением проницаемости для ионов Nа+ и К+, что приводит к ее деполяризации, возникновению потенциала действия, генерации возбудительного постсинаптического потенциала. Под влиянием этого потенциала происходит деполяризация соседних с синапсом участков мембраны. Таким образом потенциал действия распространяется по всему органу.

Медиаторы выбрасываются в синаптическую щель не только при возбуждении, но и в покое. В покое выделяются малые порции (кванты) медиатора, но под влиянием нервного импульса одновременно выбрасывается значительное количество таких квантов. Избыток медиатора разрушается специальными ферментами: ацетилхолин — холинэстеразой; норадреналин — моноаминооксидазой; катехол-О - метилтрансферазой.

Свойства синапсов 1. Одностороннее проведение возбуждения — 1. возбуждение проводится через синапс только в одну сторону (в одном направлении) с нерва (аксона) на мышцу (другой орган). 2. Замедленное проведение возбуждения — 2. возбуждение через синапсы проводится с задержкой в сравнении с нервным волокном. Объясняется это тем, что проведение возбуждения — многоэтапный процесс: время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, активацию мембраны, рост потенциала действия до пороговой величины.

3. Утомление — связано с уменьшением 3. резерва медиатора при длительном поступлении импульсов. 4. Низкая лабильность — обусловлена 4. тем, что проведение возбуждения через синапс сопряжено с затратой относительно большого периода времени.

В гладких мышцах и в центральной нервной системе имеются и тормозные синапсы. Нервные импульсы высвобождают из нервных окончаний содержащийся в них тормозной медиатор: норадреналин — в кишечнике, бронхах; ацетилхолин — в мышечных клетках сфинктера мочевого пузыря, водителя ритма сердца и других; гамма-аминомасляную кислоту — в центральной нервной системе и др.
2 Физиология возбудимых тканей.ppt