Лекция 2 нерв син мыш.ppt
- Количество слайдов: 71



Лекция № 2 Физиология возбудимых тканей Для студентов заочного отделения

Цель: ознакомится с функциями возбудимых тканей План: • Физиология нервных волокон • Нервно-мышечный синапс • Физиология скелетных мышц

ФИЗИОЛОГИЯ НЕРВНЫХ ВОЛОКОН Физиологические свойства: • раздражимость • возбудимость, • проводимость, • рефрактерность, • лабильность.
 и • безмиелиновые (безмякотные)")
В зависимости от гистологического строения различают • миелиновые (мякотные) и • безмиелиновые (безмякотные)

• Возбуждения по нервам осуществляется на основе ионных механизмов генерации ПД.

• Возбуждение по безмиелиновым волокнам идет непрерывно. Этот процесс происходит в каждой точке мембраны на всем протяжении волокна.
 и участки лишенные оболочки -перехваты Ранвье. Электрические")
• Миелиновые волокна имеют миелин (изолятор) и участки лишенные оболочки -перехваты Ранвье. Электрические токи не могут проходить через миелин, они возникают только между соседними перехватами Ранвье, Перехват Ранвье миелин Строение миелинового волокна. Возбуждение идет скачкообразно

В зависимости от скорости проведения возбуждения нервные волокна делятся на три основных типа: А, В, С. • Волокна типа А – это миелиновые волокна, скорость проведения возбуждения 5 -120 м/с, диаметр 12 -22 мкм • А α - иннервирующие скелетные мышцы; • А β - афферентные волокна от мышечных рецепторов, от рецепторов прикосновения и давления, • А- (дельта) афференты от температурных и болевых рецепторов;

• Волокна типа В - миелиновые преганглионарные волокна вегетативной нервной системы, скорость проведения возбуждения 3 -18 м/с, диаметр 1 -3 мкм • Волокна типа С - безмиелиновые нервные волокна со скоростью проведения возбуждения 0, 5 -3 м/с, диаметр 0, 5 -1 мкм. Это постганглионарные волокна вегетативной нервной системы, а так же афферентные волокна от рецепторов давления, боли.

Скорость распространения возбуждения по нервам

Синапс • —это специализированная структура, которая обеспечивает передачу возбуждения с одной возбудимой структуры на другую (Шеррингтон).

А Морфология синапсов. Б Salpeter M. 1983 А. Лягушка Б. Птица Slater C. R. 2003

Структурно-функциональная характеристика нервно-мышечного синапса • 1. Пресинаптическое окончание • 2. Синаптическая щель • 3. Постсинаптическая мембрана

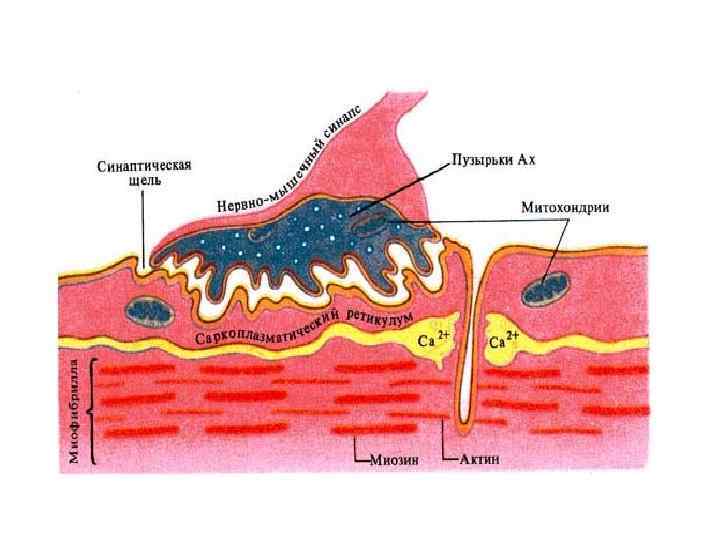
1. Пресинаптическое окончание • образуется расширением по ходу окончания аксона, иннервирующего мышечное волокно. • Пресинаптическая мембрана - это часть мембраны нервного окончания в области контакта его с мышечным волокном
 Синаптические пузырьки- везикулы, заполненные медиатором ацетилхолином и АТФ. 2) Митохондрии")
Структура пресинаптического окончания 1) Синаптические пузырьки- везикулы, заполненные медиатором ацетилхолином и АТФ. 2) Митохондрии – для энергетического обеспечения; 3)Цистерны гладкой эндоплазматической сети, содержащие депонированный Са;

• Через пресинаптическую мембрану осуществляется выделение - экзоцитоз - медиатора АХ в синаптическую щель.

2. Синаптическая щель содержит: • межклеточную жидкость, • фермент ацетилхолинэстеразу • мукополисахаридное плотное вещество в виде полосок, мостиков, которое обеспечивает связь между пре- и постсинаптической мембранами

3. Постсинаптическая мембрана • утолщенная часть клеточной мембраны мышечного волокна. • Содержит белковые рецепторы способные связать молекулы медиатора, активируя ионные каналы. рецептор Ионный канал

• Наличие множества складок, которые образуют слепые карманы, благодаря им резко увеличивается площадь постсинаптической мембраны и количество рецепторов. • Постсинаптическую мембрану нервномышечного синапса так же называют концевой пластинкой.

Механизм синаптической передачи и ее регуляция осуществляется в 2 этапа: 1. Преобразование электрического сигнала в химический. 2. Преобразование химического сигнала обратно в электрический

1. Преобразование электрического сигнала в химический. ПД, поступивший в пресинаптическое окончание, вызывает деполяризацию ее мембраны. Открываются потенциалзависимые Са 2+-каналы. Са 2+ входит внутрь клетки. Ионы Са 2+ активируют везикулы, и везикула перемещается к пресинаптической мембране.

При контакте везикулы с пресинаптической мембраной происходит «плавление» ее стенки и медиатор выходит в синаптическую щель посредством -экзоцитоза. В нервно-мышечном синапсе медиатор является АХ (ацетилхолин).

Молекулы АХ поступают в синаптическую щель, Направление ПД диффундируют к постсинаптической мембране и вступают везикулы во взаимодействие с ее рецепторами. Остальные разрушаются ферментом Синаптическая щель холинэстеразой. Пресинаптическое окончание митохондрии Постсинаптическая мембрана Строение синапса

• В нервно-мышечном синапсе АХ взаимодействует с Н-холинорецепторами, которые способны активироваться под влиянием никотина, вследствие чего они и получили свое название, • они имеют в своем составе Na/К канал.

2 этап: Преобразование химического сигнала обратно в электрический • Этот этап осуществляется на постсинаптической мембране (в мышце) АХ взаимодействует с Н-холинорецепторами происходит открытие каналов для ионов Na, Ионы натрия входят внутрь мышечного волокна через постсинаптическую мембрану, в результате происходит деполяризация. • Эта деполяризация называется возбуждающим постсинаптическим потенциалом (ВПСП), а в нервно-мышечном, потенциал концевой пластинки (ПКП).

Физиология мышц

Существует три типа мышц: Рис. Поперечнополосатая мышца Рис. Сердечная мышца Рис. Гладкая мышца

Физиологические свойства мышц: • • • 1. возбудимость 2. сократимость 3. рефрактерность 4. лабильность 5. проводимость 6. автоматизм

ФИЗИЧЕСКИЕ СВОЙСТВА МЫШЦ • 1. Растяжимость - способность изменять длину под влиянием приложенной силы. • 2. Эластичность - способность восстанавливать первоначальную форму после прекращения действия ее сил. • 3. Сила мышц - определяется тем максимальным грузом, которой мышца в состоянии чуть-чуть приподнять. • 4. Способность совершать работу - определяется произведением величины поднятого груза на высоту подъема.

Функциональная единица мышц – мышечное волоконо. • Мышечное волокно окружено мембраной - сарколеммой, сходной по электрическим свойствам с мембраной нервных клеток. • В состав мышечных волокон входит большое количество миофибрилл, которые и создают характерную поперечную исчерченность. • Каждая из миофибрилл состоит из повторяющихся в продольном направлении одинаковых участков саркомеров. • Расположение саркомеров совпадают, это и создает картину "полосатости", или поперечной исчерченности волокна.

Саркоплазматический ретикулум – депо кальция

Саркомеры отделены друг от друга Z-мебранами и представляют собой функциональные единицы поперечно-полосатой мышцы. Z-мембрана актин головки миозина Z-мембрана миозин саркомер Z-мембрана

К Z-мембране прикреплены тонкие актиновые нити, между ними – толстые миозиновые нити
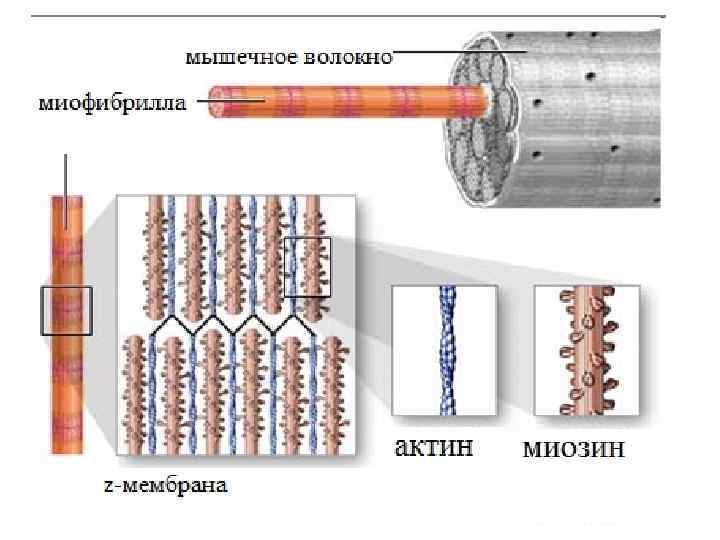

Структура миофибрилл • Сведения о белковой структуре компонентов поперечнополосатых мышц были получены еще в середине XIX века. • Вильгелм Куне показал, что в водных растворах солей из измельченной мышцы экстрагируется целый ряд белковых фракций.
 нить • образована двумя спиральными тяжами из глобулярных молекул актина (G -")
Актиновая (тонкая) нить • образована двумя спиральными тяжами из глобулярных молекул актина (G - актин), закрученных один вокруг другого. Мономером является молекула G - актина, названного так из-за своей глобулярной формы. Полимеризуясь они образуют длинную двойную спираль F - актин (от англ. filamentous - нитевидный).

В состав актиновой нити входят два вспомогательных белка - тропомиозин и тропонин • Тропомиозин - располагается в виде нитей в продольных бороздках F- актиновой спирали. В покое закрывает активные зоны актина • К тропомиозину прикреплен комплекс молекул тропонина, чувствительных к ионам Са 2+

Миозиновые нити имеют головки, которые отходят от нити биполярно. Головки обладает энергией АТФ

Теория мышечного сокращения • В 1954 г. А. Хаксли при использовании микроскопа установил, что актиновые и миозиновые филаменты не изменяют своей длины как при укорочении или удлинении саркомера. • В момент изменения длины саркомера происходит перекрывание актиновых и миозиновых филаментов, в результате их скольжения. • Теория скольжения нитей (Хаксли, 1954)

Сокращение. Теория скольжения Хаксли

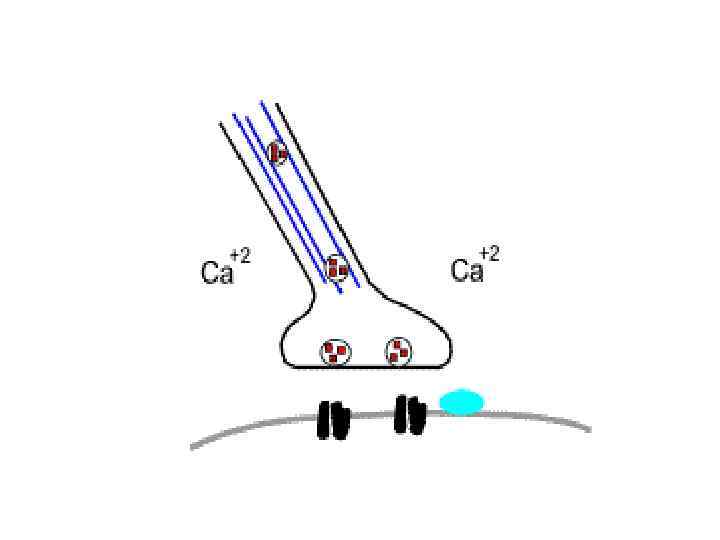
• При возбуждении мышечного волокна потенциал действия, возникающий в области концевой пластинки (мышечной мембраны), распространяется по Тсистеме вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазматического ретикулума и освобождает из них ионы Са 2+

Нервно-мышечный синапс

В состоянии покоя • головка миозина не прикреплена к активной зоне актиновой нити - мешает тропомиозин. • Белок тропомиозин блокирует активные участки белка актина, способные взаимодействовать с головкой миозина

Шаг 1. Ионы кальция, выходя из СР запускают процесс сокращения. • Под влиянием ионов Са 2+ молекула тропонина изменяет свою форму и выталкивает тропомиозин в желобок между двумя нитями актина, освобождая активные участки для прикрепления миозиновых головок (мостиков) к актину.

Шаг 2: Головка миозина прикрепляется к активному участку актина

Шаг 3: Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что обеспечивает энергией поперечные мостики. Мостики совершают движение к центру саркомера. Происходит укорочение саркомера.

Шаг 4: Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация ионов Са 2+ внутри миофибрилл не снизится.

Типы мышечного сокращения • 1. Изотоническое — волокна мышцы укорачиваются без изменения их напряжения • 2. Изометрическое - мышца развивает напряжение без изменения своей длины. • 3. Ауксотоническое - мышца развивает напряжение и укорачивается.

Режимы мышечных сокращений • Режим мышечных сокращений зависит от частоты импульсации мотонейронов.

Одиночное сокращение - • - механический ответ мышечного волокна на однократное раздражение. Выделяют три фазы сокращения • 1. латентная (срытая) фаза возбуждения это время, прошедшее от нанесения раздражения до момента начала механической реакции мышцы. • 2. фаза напряжения или укорочения • 3. фаза расслабления или удлинения (более продолжительна)
")
Фазы одиночного мышечного сокращения (ОМС)

Соотношения фаз ОМС с фазами возбудимости в скелетных мышцах Красный график- фазы сокращения: • 1 – латентный период • а –сокращения • б – расслабление 1 Синий график – фазы возбудимости: • в- АРФ соотв. фазе 1 • г -ОРФ соотв. фазе а • д – экзальтация соотв. пику сокращения

Тетанус • В естественных условиях к мышечным волокнам поступают не одиночные нервные импульсы, а их ряды. Когда мышца получает ряд часто следующих друг за другом нервных импульсов, она отвечает длительным сокращением. • Длительное, слитное сокращение мышцы получило название тетанического сокращения или тетануса. К тетаническому сокращению способны только скелетные мышцы. Гладкие мышцы и мышца сердца не способны к тетаническому сокращению, вследствие наличия продолжительного рефрактерного периода.

• Тетанус возникает вследствие суммации одиночных мышечных сокращений. • Для того, чтобы возник тетанус, необходимо действие повторных раздражителей (или нервных импульсов) на мышцу еще до того, как закончится ее одиночное сокращение.

Суммация мышечных сокращений

• Если раздражающие импульсы сближены и каждый из них приходится на расслабления одиночного сокращения, то возникает зубчатый тетанус (неполный несовершенный тетанус).

• Если раздражающие импульсы сближены настолько, что каждый последующий приходится на фазу сокращения, т. е. когда мышца не успела перейти к расслаблению, то возникает длительное непрерывное сокращение - гладкий тетанус.

При анализе графиков, обращает на себя внимание следующая особенность: амплитуда зубчатого и гладкого тетануса выше, чем амплитуда одиночного мышечного сокращения. • Введенский показал, что увеличение амплитуды тетануса связано с тем, что каждый последующий нервный импульс попадает в стадию экзальтации.

• Существуют оптимальные, т. е. наилучшие значения частоты раздражения, при которых возникает тетанус максимальной амплитуды – оптимум (импульс попадает в экзальтацию). • При значительном увеличении частоты раздражения амплитуда тетануса уменьшается - это наступает пессимум частоты (импульс попадает в АРФ).

• Гладкий тетанус -нормальное рабочее состояние скелетных мышц, обуславливается посылкой из ЦНС нервных импульсов с частотой 40 -50 имп. в 1 с. • Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1 с.
…. 2)… 3)…. . 4)….")
• Т. о. , выделяют режимы мышечных сокращений: 1)…. 2)… 3)…. . 4)…. 5)…. • Режимы мышечных сокращений зависят от частоты импульсации мотонейронов

Двигательная единица – совокупность мышечных волокон, иннервируемых разветвлениями одного мотонейрона.
 • 2. быстрые, легко утомляемые")
Виды двигательных единиц: • 1. медленные, малоутомляемые (красные волокна) • 2. быстрые, легко утомляемые (белые волокна) • 3. быстрые, устойчивые к утомлению.

Функциональные особенности медленных ДЕ • 1. Иннервируются высоко возбудимыми а -мотонейронами с низкой скоростью проведения возбуждения по аксону • 2. Количество мышечных волокон в ДЕ небольшое, и развивают меньшую силу сокращения. • 3. Имеют низкую активность миозиновой АТФазы и низкую скорость сокращения.

Функциональные особенности медленных ДЕ • 4. Имеют хорошее кровоснабжение, много митохондрий, миоглобина, высокий аэробный обмен, поэтому обладают низкой утомляемостью. Способны выполнять длительную маломощную работу. • 5. В регуляции движения обеспечивают мышечный тонус и позу, а также способность к длительной циклической работе - бег, плавание и др. (например у марафонцев их количество в мышцах достигает 85%).

Функциональные особенности быстрых ДЕ. • 1. Иннервируются крупными, менее возбудимыми а-мотонейронами с высокой скоростью проведения ПД по аксону. • 2. Количество мышечных волокон в ДЕ сравнительно больше, и они развивают большую силу сокращения. • 3. Имеют высокую активность миозиновой АТФ-азы и развивают высокую скорость сокращения.

Функциональные особенности быстрых ДЕ. • 4. Имеют слаборазвитую капиллярную сеть, мало митохондрий, миоглобина, но содержат много гликолитических ферментов, большой запас креатинфосфата и гликогена, анаэробный тип энергообеспечения. • 5. Способны развивать большую мощность, но быстро утомляются. • 6. В регуляции движения обеспечивают - перемещение организма в пространстве с большой скоростью и мощностью (например, у спринтеров и прыгунов количество быстрых ДЕ в мышцах достигает 90%).

Функциональные особенности быстрых, устойчивых к утомлению ДЕ. • По структурно-функциональным свойствам занимают среднее положение между медленными и быстрыми ДЕ • Вероятно, используются в быстрых ритмических движениях (ходьба, бег).

Благодарю за внимание
Лекция 2 нерв син мыш.ppt