Го-13 - 14 AA рус.ppt
- Количество слайдов: 75



Лекция № 13 РЕГУЛЯЦИЯ ОБМЕНА ВЕЩЕСТВ. БИОХИМИЯ ГОРМОНОВ. 1 МЕХАНИЗМ ДЕЙСТВИЯ ГОРМОНОВ ЧЕРЕЗ ц. АМФ и ц. ГМФ

Цель: Ознакомить с общими свойствами гормонов, первым механизмам действия гормонов, посредниками передачи действия гормонов внутри клетки

План: 1. Общие свойства гормонов 2. Первый механизм через ц. АМФ 3. Первый механизм через ц. ГМФ

Гормоны - это биологически активные вещества , образующиеся в железистых клетках, выделяющиеся в кровь или лимфу и регулирующие обмен веществ.

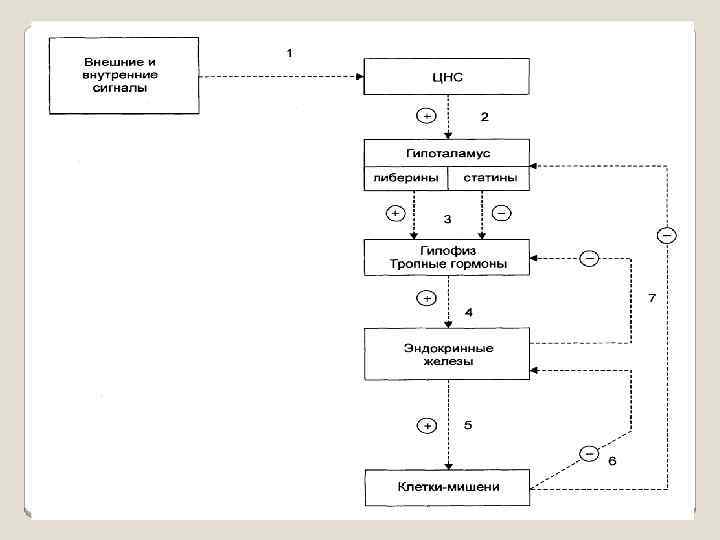
Ведущим звеном в адаптации организма является ЦНС и гипоталамо – гипофизарная система. ЦНС в ответ на раздражение посылает в гипоталамус и другие ткани в том числе на железы внутренней секреции, нервные импульсы в виде изменения концентрации ионов и медиаторов.

Гипоталамус выделяет особые вещества – нейросекретины или рилизинг- факторы двух видов: 1 Либерины, ускоряющие выделение тропных гипофизом 2: Статины угнетающие их выделение.

ГИПОТАЛАМУС окситоцин, вазопрессин АДЕНОГИПОФИЗ СТГ, ТТГ, АКТГ, ФСГ, ЛТГ, пролактин ЭПИФИЗ мелатонин ОКОЛОЩИТОВИД НАЯ ЖЕЛЕЗА паратгормон СЕРДЦЕ: натрий уретический фактор ЩИТОВИДНАЯ ЖЕЛЕЗА Т 3 , тироксин, кальцитонин ТИМУС тимозин НАДПОЧЕЧНИКИ катехоламиндер, кортикостероидтар, жыныс гормондары ПОЧКИ Эритропоэтин, ренин, простагландин ПИЩЕВАРИТЕЛЬ НЫЙ ТРАКТ Гастрин, секретин ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА инсулин , глюкагон ПОЛОВЫЕ ЖЕЛЕЗЫ Эстрадиол, прогестерон, тестостерон, релаксин, ингибин, хорионический гонадотропин Эндокринная система
 Гормоны -простые белки (инсулин, гормон роста, ЛТГ, паратгормон)")
Классификация гормонов I. Белково-пептидные гормоны 1) Гормоны -простые белки (инсулин, гормон роста, ЛТГ, паратгормон) 2) Гормоны - сложные белки (ТТГ, ФСГ, ЛГ) 3)Гормоны- полипептиды (глюкагон, АКТГ, МСГ, кальцитонин, вазопрессин, окситоцин) Некоторые из перечисленных гормонов образуются из неактивных предшественников - прогормонов (например, инсулин и глюкагон).
. III. Гормоны –")
II. Стероидные гормоны – производные холестерина (кортикостероиды, половые гормоны: мужские, женские). III. Гормоны – производные аминокислот (тироксин, трийодтиронин, адреналин, норадреналин).

Общие свойства гормонов -строгая специфичность биологического действия; -высокая биологическая активность; секретируемость; - дистантность действия; - гормоны могут находиться в крови, как в свободном, так и в соединенном с определёнными белками состоянии; - кратковременность действия; - все гормоны проявляют свое действие через рецепторы.
 По химической природе рецепторы – это белки, истинные гликопротеиды Ткани, в")
Рецепторы гормонов (РЦ) По химической природе рецепторы – это белки, истинные гликопротеиды Ткани, в которых имеются рецепторы для данного гормона называются ткани-мишени (клетки-мишени).

Биологическое действие гормона зависит не только от его содержания в крови, но и от количества и функционального состояния рецепторов, а также от уровня функционирования пострецепторного механизма
 Мембранно-цитозольный механизм гормоны,")
Все известные гормоны по механизму действия подразделяются на 3 группы: I) Мембранно-цитозольный механизм гормоны, действующие путем изменения активности внутриклеточных ферментов. Эти гормоны язываются с рецепторами на наружной поверхности мембраны клетки-мишени, внутри клетки не входят и действуют через вторичных посредников (мессенджеров): ц-АМФ, ц-ГМФ, ионы кальция, инозитолтрифосфат.
 Эти")
2. Гормоны, действующие путем изменения скорости синтеза белков и ферментов. (Цитозольный. ) Эти гормоны связываются с внутриклеточными рецепторами: цитозольными, ядерными или рецепторами органоидов. К данным гормонам относятся стероидные и тироидные гормоны
 К таким гормонам относятся")
3. Гормоны, действующие путем изменения проницаемости плазматической мембраны (мембранный. ) К таким гормонам относятся инсулин, СТГ, ЛТГ, АДГ.

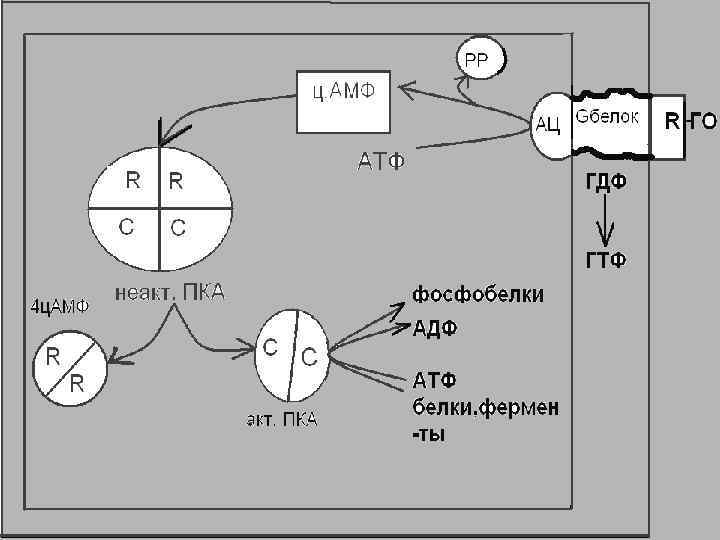
1 -й МЕХАНИЗМ Аденилатциклазная система состоит из 3 -х частей: I - узнающая часть, представленная рецептором, расположенным на наружной поверхности клеточной мембраны, . II часть - сопрягающий белок (G-белок). В неактивном виде G-белок связан своей субъединицей с ГДФ.

III часть - каталитическая является ферментом аденилатциклазой аденилатциклаза АТФ Н 4 Р 2 О 7 + ц. АМФ взаимодействует с протеинкиназой А, которая состоит из 4 -х субъединиц: 2 -х регуляторных, 2 -х каталитических.

ц-АМФ связывается с регуляторными субъединицами, две каталитические освобождаются, протеинкиназа А становится активной.

Протеинкиназа А катализирует перенос от АТФ фосфатной группы на ОН-группы серина и треонина ряда белков и ферментов клетокмишеней, т. е. является серинтреонин-киназой АТФ АДФ Белок белок-Р

Белками, на которые будут переноситься остатки фосфорной кислоты при фосфорилировании с участием протеинкиназы А, могут быть некоторые ферменты (например, фосфорилаза, липаза, гликогенсинтетаза, метилтрансферазы), белки рибосом, ядер, мембран. При фосфорилировании неактивных форм фосфорилазы и липазы наблюдаются конформационные изменения в их молекулах, что ведет к повышению их активности.

Фосфорилирование гликогенсинтетазы , наоборот, тормозит ее активность. Присоединение фосфорной кислоты к белкам рибосом повышает синтез белка.
 и ДНК")
Если фосфорная кислота присоединяется к ядерным белкам, то связь между белком (гистоном) и ДНК ослабляется, что ведет к усилению транскрипции, а значит и к повышенному синтезу белков. Фосфорилирование белков мембран повышает их проницаемость для ряда веществ, в частности для ионов.

Под действием гормонов, действующих через ц. АМФ, ускоряется: 1. Гликогенолиз путем фосфоролиза, 2. липолиз, 3. синтез белков, 4. транспорт ионов через мембраны, 5. Ингибируется гликогенез

Первый механизм через ц. ГМФ

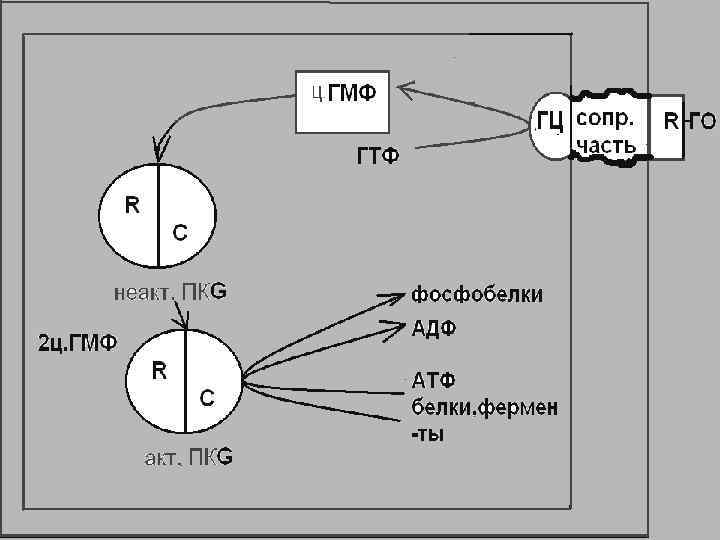
По этому механизму действуют гормоны через гуанилатциклазную систему. Гуанилатциклаза имеет мембраносвязанную и растворимую (цитозольную) формы Мембраносвязанная форма состоит из 3 -х участков: 1 - узнающего (на внешней стороне плазматической мембраны)

2 - го - Трансмембранного 3 -го - Каталитического Мембранносвязанная форма фермента активируется через рецепторы короткими пептидами, например, предсердным натрий-уретическим фактором.

Натрий-уретический фактор синтезируется в предсердии в ответ на повышение объема циркулирующий крови , поступает в почки, активирует в них гуанилатциклазу, что приводит к повышению экскреции натрия и воды

Гладкомышечные клетки также содержат гуанилатциклазную систему через которую осуществляется их расслабление. Действуют через эту систему вазодилятаторы, как эндогенные (оксид азота), так и экзогенные

В эпителиальных клетках кишечника активатором гуанилатциклазы может быть бактериальный эндотоксин, который приводит к замедлению всасывания воды и диареи. Цитозольная форма гуанилатциклазыгемсодержащий фермент
 продукты ПОЛ Под")
В регуляции его активности участвуют нитровазодилятаторы, активные формы кислорода (оксид азота) продукты ПОЛ Под действием гуанилатциклазы из ГТФ образуется ц. ГМФ Ц-ГМФ действует на протеинкиназу G состоящую из двух субъединиц

ц. ГМФ связывается с регуляторными участками ПК G активируя ее. ПКА и ПК G являются серин- треонинкиназами, и ускоряя фосфорилирование серина и треонина разных белков и ферментов оказывают различное биологическое действие.
 под действием натрий-уретического фактора усиливается диурез (этот гормон-пептид образуется в предсердиях) 2) под")
1) под действием натрий-уретического фактора усиливается диурез (этот гормон-пептид образуется в предсердиях) 2) под действием бактериальных эндотоксинов развивается диарея

Один и тот же гормон может действовать и через ц. ГМФ и через ц. АМФ. Эффект зависит от того с каким рецептором связывается гормон. Например, адреналин может связываться как с альфа , так и с бетта – рецепторами.

Образование комплекса адреналина с бетта- рецепторами ведет к образованию ц. АМФ. Образование комплекса адреналина с альфа- рецепторами ведет к образованию ц. ГМФ. Эффекты , оказываемые адреналином будут различными.

ПК G повышает активность гликогенситетазы, тормозит агрегацию тромбоцитов, активирует фосфолипазу С, освобождая Са из его депо. Т. о. , по своему действию ц. ГМФ является антагонистом ц. АМФ
 под действием оксида азота происходит расслабление гладкомышечных клеток сосудов (что используется в")
3) под действием оксида азота происходит расслабление гладкомышечных клеток сосудов (что используется в медицине, так как ряд нитропрепаратов, таких как нитроглицерин, используются для снятия спазмов сосудов)

Снятие сигнала гормона, действующего через ц. АМФ и ц. ГМФ, происходит следующим образом: 1. гормон быстро разрушается, и, следовательно, разрушается комплекс гормон-рецептор

2. для снятия гормонального сигнала в клетках существует особый фермент фосфодиэстераза, который превращает циклические нуклеотиды в нуклеозидмонофосфаты (адениловую и гуаниловую кислоты соответственно)

3. фосфорилированные ферменты под влиянием протеинфосфатаз инактивируются

Т. Ш. Шарманов, С. М. Плешкова «Метаболические основы питания с курсом общей биохимии» , Алматы, 1998 г. С. Тапбергенов «Медицинская биохимия» , Астана, 2001 г. С. Сеитов «Биохимия» , Алматы, 2001 г. Стр 342 -352, 369 - 562 В. Дж. Маршал «Клиническая биохимия» , 2000 г. Н. Р. Аблаев Биохимия в схемах и рисунках, Алматы 2005 г. Стр 199 -212 Биохимия. Краткий курс с упражнениями и задачами. Под ред. проф. Е. С. Северина, А. Я. Николаева, М. , 2002 г. Северин Е. С. «Биохимия» 2008, Москва, стр 534 -603 Березов Т. Т. , Коровкин Б. Ф. 2002 «Биологическая химия» , стр 248 -298.

Контрольные вопросы: 1. Общие свойства гормонов 2. Классификация гормонов 3. Посредники действия гормонов первого механизма 4. Роль ц АМФ и ц ГМФ

Лекция № 14 Регуляция обмена веществ Первый механизм действия гормонов через ионы кальция, ДАГ и ИТФ. Второй и третий механизмы действия.

Ознакомить с особенностями действия гормонов через посредники: ионы кальция, ДАГ, ИТФ, действие стероидных гормонов- второй механизм, мембранным механизмом Цель:

Посредники действия гормонов – ионы кальция, ДАГ, ИТФ Второй механизм действия Особенности действия гормонов по третьему механизму. План:

Внутриклеточными посредниками гормонов могут служить ионы кальция, играющие роль регулятора благодаря изменению своей концентрации.
, а снаружи клетки и")
Внутри клетки концентрация ионов кальция ничтожно мала (10¯ 7 моль/л), а снаружи клетки и внутри органоидов выше (10¯ 3 моль/л).

Поступление кальция из внешней среды внутрь клетки осуществляется по кальциевым каналам мембраны. Поток кальция регулируется Са-зависимой АТФазой мембраны, в осуществлении ее функции регулирующую роль могут выполнять инозитолтрифосфат (IP 3) и инсулин.

Внутри клетки ионы Са 2+ депонируются в матриксе митохондрий и эндоплазматическом ретикулуме. Са 2+, поступающий в цитоплазму из внешней среды или из внутриклеточных депо, взаимодействует с Са 2+-зависимой кальмодулинкиназой.

Кальций связывается с регуляторной частью фермента, это кальцийсвязывающий белок – кальмодулин, при этом происходит активация фермента.
 для связывания с ионами кальция или")
Кальмодулин имеет несколько центров (до 4 -х) для связывания с ионами кальция или магния. В покое кальмодулин связан с магнием, при увеличении концентрации кальция в клетке, кальций вытесняет магний.
 для связывания с ионами кальция или")
Кальмодулин имеет несколько центров (до 4 -х) для связывания с ионами кальция или магния. В покое кальмодулин связан с магнием, при увеличении концентрации кальция в клетке, кальций вытесняет магний.

При значительном увеличении кальция образуется комплекс 4 Са 2+кальмодулин, который активирует гуанилатциклазу и фосфодиэстеразу ц. АМФ.

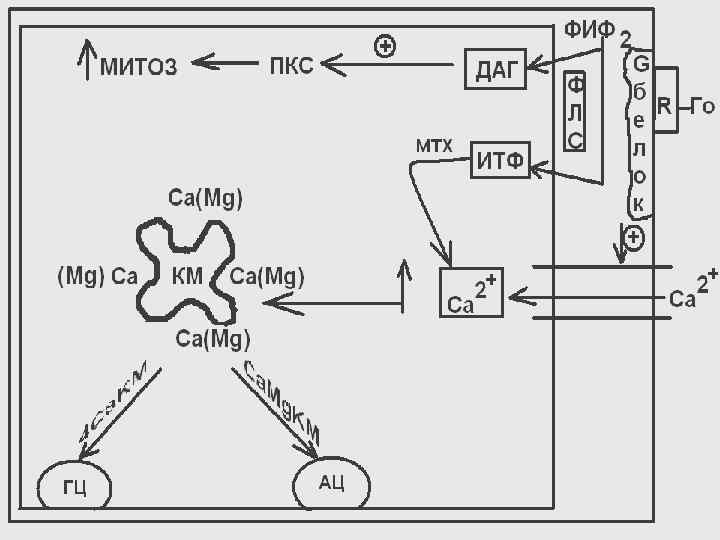
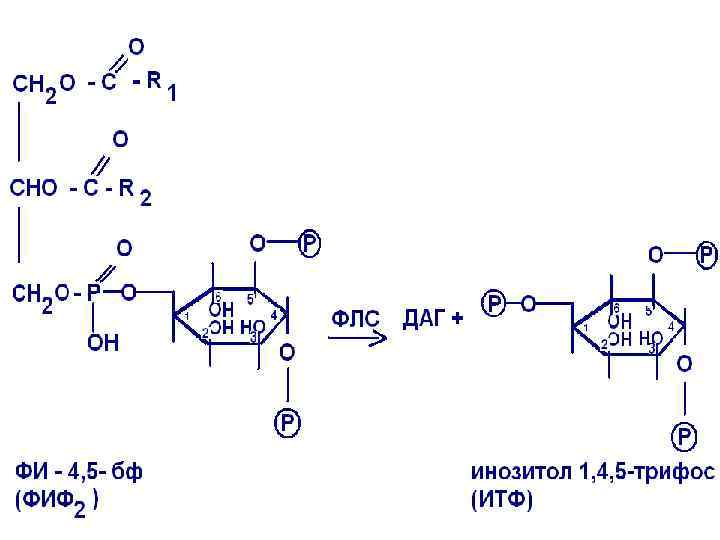
Действие гормонов через ионы кальция часто сочетается с использованием в качестве посредника производных фосфатидилинозитола. Рецептор в таких случаях находится в комплексе с Gбелком и при взаимодействии рецептора с гормоном (например, ТТГ, пролактин, СТГ)

происходит активация мембраносвязанного фермента фосфолипазы С, которая ускоряет реакцию распада фосфатидилинозитол 4, 5 -дифосфата с образованием ДАГ и инозитол-1, 4, 5 -трифосфата.

ДАГ и инозитолтрифосфат являются вторичными посредниками в действии соответствующих гормонов. ДАГ вызывает активацию протеинкиназы С, которая, в свою очередь, вызывает фосфорилирование белков ядер, тем самым, усиливается пролиферация клеток-мишеней.
.")
Под действием IР 3 усиливается выход кальция из внутриклеточных депо (эндоплазматического ретикулума).

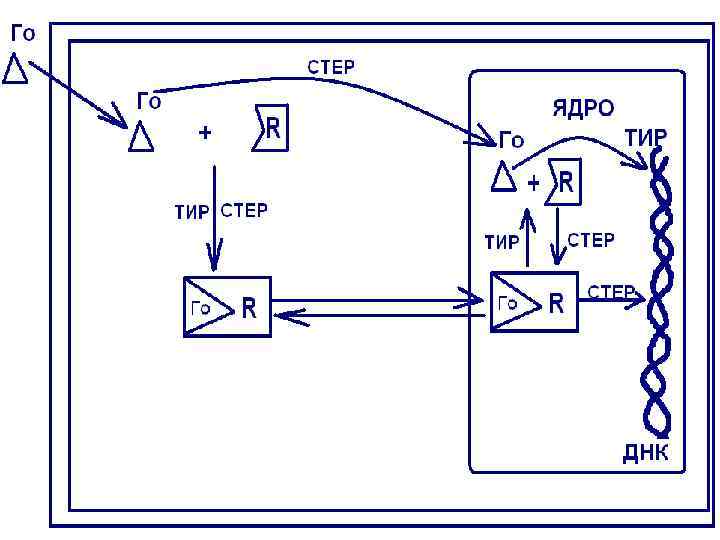
Второй механизм Этот механизм действия характерен для гормонов, которые могут легко проникать через мембрану(стероидные и тироидные)
, где и образуется гормон рецепторный комплекс.")
Возможно проникновение гормона сразу в ядро (стероидные гормоны), где и образуется гормон рецепторный комплекс. В ядре стероидрецепторный комплекс связывается с регуляторными белками хроматина, регулируя, таким образом, транскрипцию.

Образуется м-РНК, она поступает в цитозоль, формируя матрицу, где идет синтез белков – ферментов. Действие ферментов активируется за счет увеличения их концентрации. По этому механизму действуют гормоны адаптации: стероидные и тиреоидные

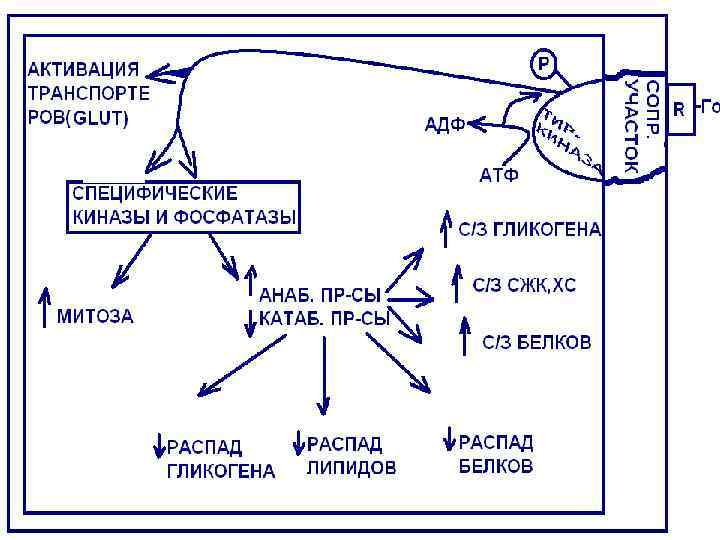
Третий механизм действия гормонов
 для различных субстратов (")
Гормоны, действующие путем изменения проницаемости плазматической мембраны (мембранный. ) для различных субстратов ( аминокислоты, глюкоза, глицерин и др. )

Эти гормоны связываются с рецепторами плазматических мембран и свое действие опосредуют через тирозинкиназно-фосфатазную систему.

При этом происходит изменение активности внутриклеточных ферментов, сопровождающееся активацией белков-транспортеров и ионных каналов. К таким гормонам относятся инсулин, СТГ, ЛТГ, АДГ.

Инсулин действует через мембраносвязанную тирозин- киназу, Активируя транспортные белки, переносящие глюкозу, активируются специфические ферменты.

Гормоны СТГ, ЛДГ образуя гормонрецепторный комплекс активируют цитозольную тирозин-киназу, которая действует подобно мембраносвязанной, активируется фосфолипаза С, что ведет к мобилизации Са +2 и активированию протеинкиназы С.
, усиливается реабсорбция воды")
АДГ действуя через ц. АМФ, вызывает перемещение водных каналов (белковаквапоринов), усиливается реабсорбция воды в почках, снижается выделение мочи, т. е АДГ увеличивает проницаемость мембран клетокмишеней для воды.

Т. Ш. Шарманов, С. М. Плешкова «Метаболические основы питания с курсом общей биохимии» , Алматы, 1998 г. С. Тапбергенов «Медицинская биохимия» , Астана, 2001 г. С. Сеитов «Биохимия» , Алматы, 2001 г. Стр 342 -352, 369 - 562 В. Дж. Маршал «Клиническая биохимия» , 2000 г. Н. Р. Аблаев Биохимия в схемах и рисунках, Алматы 2005 г. Стр 199 -212 Биохимия. Краткий курс с упражнениями и задачами. Под ред. проф. Е. С. Северина, А. Я. Николаева, М. , 2002 г. Северин Е. С. «Биохимия» 2008, Москва, стр 534 -603 Березов Т. Т. , Коровкин Б. Ф. «Биологическая химия» , стр 248298. Литература:

Контрольные вопросы: 1. Роль ц. ГМФ в механизме действия гормонов 2. Роль Са и ИТФ в механизме действия гормонов 3. Второй механизм – изменение скорости синтеза белков- ферментов 4. Третий механизм – изменение механизма проницаемости клеточной мембраны.
Го-13 - 14 AA рус.ppt