Лекция 11-1-такт.ан.ppt
- Количество слайдов: 44
. Рецепторные образования кожи. Тельца") Лекция 11. Кожный и двигательный анализатор. (Часть 1 -Тактильная чувствительность). Рецепторные образования кожи. Тельца Паччини, Мейснера, диски Меркеля, свободные нервные окончания. Характеристики рецепторов. Проводящие пути тактильной чувствительности.
Лекция 11. Кожный и двигательный анализатор. (Часть 1 -Тактильная чувствительность). Рецепторные образования кожи. Тельца Паччини, Мейснера, диски Меркеля, свободные нервные окончания. Характеристики рецепторов. Проводящие пути тактильной чувствительности.
 Кожно-мышечное чувство объединяет все сенсорные анализаторы, чьи рецепторы находятся в коже, мышцах, а также связках и сухожилиях. Выделяют несколько типов чувствительности: тактильную (осязание), температурную, чувствительность мышц и сухожилий (проприорецепцию), а также болевую. Отличительная особенность этой системы – отсутствие специализированных сложных структур.
Кожно-мышечное чувство объединяет все сенсорные анализаторы, чьи рецепторы находятся в коже, мышцах, а также связках и сухожилиях. Выделяют несколько типов чувствительности: тактильную (осязание), температурную, чувствительность мышц и сухожилий (проприорецепцию), а также болевую. Отличительная особенность этой системы – отсутствие специализированных сложных структур.
 Кожа представляет собой огромное рецепторное поле, чувствительные нервные окончания постоянно осуществляют связь с ЦНС. В среднем на 1 см 2 кожи приходится до 5000 чувствительных окончаний, 200 болевых, 12 холодовых точек, 2 тепловых и 25 точек, реагирующих на давление.
Кожа представляет собой огромное рецепторное поле, чувствительные нервные окончания постоянно осуществляют связь с ЦНС. В среднем на 1 см 2 кожи приходится до 5000 чувствительных окончаний, 200 болевых, 12 холодовых точек, 2 тепловых и 25 точек, реагирующих на давление.
 Рецепторные образования кожи Рецепторы, расположенные в коже, обеспечивают три типа чувствительности: тактильную температурную и болевую. В зависимости от структуры все рецепторные образования кожи делят на две группы: свободные и несвободные (инкапсулированные и неинкапсулированные). Свободные — это нервные окончания, не имеющие вспомогательных структур и свободно расположенные в ткани. Инкапсулированные рецепторы включаются в комплекс со вспомогательными клетками.
Рецепторные образования кожи Рецепторы, расположенные в коже, обеспечивают три типа чувствительности: тактильную температурную и болевую. В зависимости от структуры все рецепторные образования кожи делят на две группы: свободные и несвободные (инкапсулированные и неинкапсулированные). Свободные — это нервные окончания, не имеющие вспомогательных структур и свободно расположенные в ткани. Инкапсулированные рецепторы включаются в комплекс со вспомогательными клетками.
: выделяют прикосновение, давление, вибрацию. Эти ощущения обеспечиваются наличием в коже механорецепторов.") Тактильные ощущения (осязание): выделяют прикосновение, давление, вибрацию. Эти ощущения обеспечиваются наличием в коже механорецепторов. Кожные рецепторы: 1 — тельце Мейснера; 2 — диски Меркеля; 3 — тельце Паччини; 4 — рецептор волосяного фолликула; 5 — тактильный диск (тельце Пинкуса—Игго); 6 — окончание Руффини Механорецепторы - это вид рецепторов, которые воспринимают механическое воздействие, деформацию, действующую извне, или возникающие во внутренних органах.
Тактильные ощущения (осязание): выделяют прикосновение, давление, вибрацию. Эти ощущения обеспечиваются наличием в коже механорецепторов. Кожные рецепторы: 1 — тельце Мейснера; 2 — диски Меркеля; 3 — тельце Паччини; 4 — рецептор волосяного фолликула; 5 — тактильный диск (тельце Пинкуса—Игго); 6 — окончание Руффини Механорецепторы - это вид рецепторов, которые воспринимают механическое воздействие, деформацию, действующую извне, или возникающие во внутренних органах.
 В коже человека, лишенной волос, существует четыре основных типа таких рецепторов: тельце Пачини, тельце Мейснера, диски Меркеля и окончания Руффини. Имеются также рецепторы волосяных луковиц, которые реагируют в случае отклонения волоса. Кожные рецепторы: 1 — тельце Мейснера; 2 — диски Меркеля; 3 — тельце Паччини; 4 — рецептор волосяного фолликула; 5 — тактильный диск (тельце Пинкуса—Игго); 6 — окончание Руффини Инкапсулированные рецепторы включаются в комплекс со вспомогательными клетками. Среди инкапсулированных механорецепторов кожи различают: диски Меркеля, которые могут группироваться в тельца Пинкуса—Игго (рецепторы давления); окончания Руффини (также рецепторы давления); тельца Мейснера (рецепторы прикосновения); тельца Паччини (рецепторы смещения). Для участков кожи, покрытых волосками, характерны рецепторы волосяных фолликулов — свободные нервные окончания, обвивающие основание кожного волоска.
В коже человека, лишенной волос, существует четыре основных типа таких рецепторов: тельце Пачини, тельце Мейснера, диски Меркеля и окончания Руффини. Имеются также рецепторы волосяных луковиц, которые реагируют в случае отклонения волоса. Кожные рецепторы: 1 — тельце Мейснера; 2 — диски Меркеля; 3 — тельце Паччини; 4 — рецептор волосяного фолликула; 5 — тактильный диск (тельце Пинкуса—Игго); 6 — окончание Руффини Инкапсулированные рецепторы включаются в комплекс со вспомогательными клетками. Среди инкапсулированных механорецепторов кожи различают: диски Меркеля, которые могут группироваться в тельца Пинкуса—Игго (рецепторы давления); окончания Руффини (также рецепторы давления); тельца Мейснера (рецепторы прикосновения); тельца Паччини (рецепторы смещения). Для участков кожи, покрытых волосками, характерны рецепторы волосяных фолликулов — свободные нервные окончания, обвивающие основание кожного волоска.
 Верхний ряд - схемы рецептивных полей, средний - морфология рецепторов, нижний - электрическая активность рецепторов. (а) Быстро адаптирующиеся рецепторы: тельца Мейснера и тельца Пачини. (б) Медленно адаптирующиеся рецепторы: диски Меркеля и тельца Руффини. В нижнем ряду - ответы на синусоидальные стимулы (а) и на надавливание на кожу (б). Тактильные рецепторы различаются по скорости своей адаптации. Есть быстро адаптирующиеся (фазные) и медленно адаптирующиеся (статические) рецепторы. Быстро адаптирующиеся рецепторы регистрируют силу и появление сигнала, а медленно адаптирующиеся рецепторы регистрируют наличие сигнала на всем протяжении стимуляции, то есть они дают информацию о длительности сигнала.
Верхний ряд - схемы рецептивных полей, средний - морфология рецепторов, нижний - электрическая активность рецепторов. (а) Быстро адаптирующиеся рецепторы: тельца Мейснера и тельца Пачини. (б) Медленно адаптирующиеся рецепторы: диски Меркеля и тельца Руффини. В нижнем ряду - ответы на синусоидальные стимулы (а) и на надавливание на кожу (б). Тактильные рецепторы различаются по скорости своей адаптации. Есть быстро адаптирующиеся (фазные) и медленно адаптирующиеся (статические) рецепторы. Быстро адаптирующиеся рецепторы регистрируют силу и появление сигнала, а медленно адаптирующиеся рецепторы регистрируют наличие сигнала на всем протяжении стимуляции, то есть они дают информацию о длительности сигнала.
 Предполагают, что скорость адаптации фазных и статических рецепторов зависит от изменения проницаемости мембраны нейронов для ионов К+ вследствие продолжительной стимуляции. Проницаемость мембран нейронов быстро адаптирующихся рецепторов меняется быстрее, чем проницаемость мембран нейронов медленно адаптирующихся нейронов. При каждом нервном импульсе некоторое количество ионов Са²+ поступает внутрь клетки из внешней среды. При этом довольно скоро концентрация этих ионов около внутренней поверхности наружной мембраны становится высокой. Ионы Са²+ активируют калиевые ионные каналы и ионы калия выходят наружу. Это затрудняет процесс деполяризации мембраны, а следовательно, дальнейшее формирование нервных импульсов.
Предполагают, что скорость адаптации фазных и статических рецепторов зависит от изменения проницаемости мембраны нейронов для ионов К+ вследствие продолжительной стимуляции. Проницаемость мембран нейронов быстро адаптирующихся рецепторов меняется быстрее, чем проницаемость мембран нейронов медленно адаптирующихся нейронов. При каждом нервном импульсе некоторое количество ионов Са²+ поступает внутрь клетки из внешней среды. При этом довольно скоро концентрация этих ионов около внутренней поверхности наружной мембраны становится высокой. Ионы Са²+ активируют калиевые ионные каналы и ионы калия выходят наружу. Это затрудняет процесс деполяризации мембраны, а следовательно, дальнейшее формирование нервных импульсов.
 Тельца Паччини, Мейснера, диски Меркеля, свободные нервные окончания. Тельца Паччини Тельца Пачини были подробно изучены в 1835 году итальянским ученым Ф. Пачини. Тельца Пачини – это инкапсулированные рецепторы овальной формы, длиной 0, 5 - 2 мм. Находятся по всей дерме и подкожной жировой ткани. На срезах - это луковицеобразная структура, состоящая из множества слоев соединительной ткани, окружающих немиелинизированное нервное волокно.
Тельца Паччини, Мейснера, диски Меркеля, свободные нервные окончания. Тельца Паччини Тельца Пачини были подробно изучены в 1835 году итальянским ученым Ф. Пачини. Тельца Пачини – это инкапсулированные рецепторы овальной формы, длиной 0, 5 - 2 мм. Находятся по всей дерме и подкожной жировой ткани. На срезах - это луковицеобразная структура, состоящая из множества слоев соединительной ткани, окружающих немиелинизированное нервное волокно.
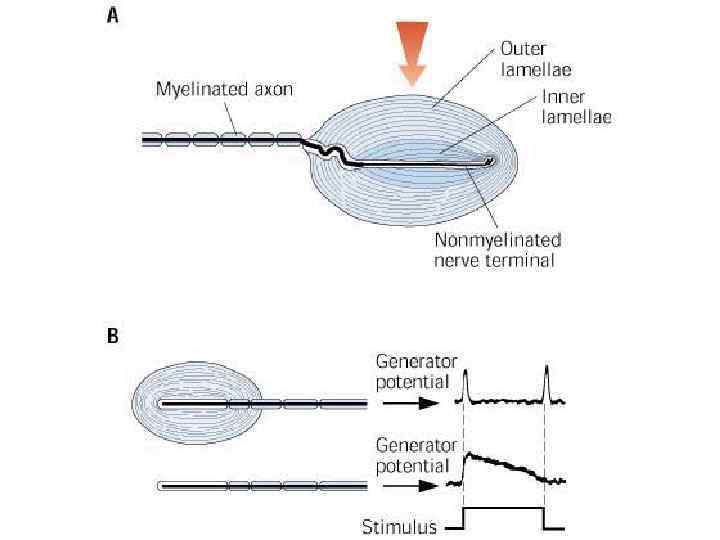
, т. е. регистрируют") Тельца Пачини являются быстроадаптирующимися (реагируют только в момент начала воздействия), т. е. регистрируют силу давления. Обладают большими рецептивными полями, т. о. представляют грубую чувствительность. Полагают, что слоистая структура преобразует постоянное по величине сдавливание в кратковременные стимулы. При сдавливании это достигается быстрым скольжением слоев тельца относительно друга до достижения вскоре некоторого нового равновесия, устраняющего сдавливание. Таким образом, пачиниевы тельца способны детектировать вибрацию даже тогда, когда находятся под постоянным давлением. разрез тельца Пачиниева: 1 - внутренняя колбочка; 2 - нервное волокно
Тельца Пачини являются быстроадаптирующимися (реагируют только в момент начала воздействия), т. е. регистрируют силу давления. Обладают большими рецептивными полями, т. о. представляют грубую чувствительность. Полагают, что слоистая структура преобразует постоянное по величине сдавливание в кратковременные стимулы. При сдавливании это достигается быстрым скольжением слоев тельца относительно друга до достижения вскоре некоторого нового равновесия, устраняющего сдавливание. Таким образом, пачиниевы тельца способны детектировать вибрацию даже тогда, когда находятся под постоянным давлением. разрез тельца Пачиниева: 1 - внутренняя колбочка; 2 - нервное волокно
 может обнаруживаться в немиелинизированном окончании, когда тельце Пачини сдавлено. При") Генераторный потенциал (деполяризация) может обнаруживаться в немиелинизированном окончании, когда тельце Пачини сдавлено. При этом возникает короткий залп импульсов в сенсорном волокне, который угасает за 1 - 2 секунды до нуля или очень малой частоты. Предполагают, что основной функцией пачиниевых телец кожи является детектирование вибрации. Частоты вибраций, на которые они реагируют, находятся в диапазоне 70 - 1000 Гц. Наиболее чувствительны они, однако, в диапазоне 200 - 400 Гц, где деформация кожи всего на 1 мкм является достаточным стимулом. разрез тельца Пачиниева: 1 - внутренняя колбочка; 2 - нервное волокно
Генераторный потенциал (деполяризация) может обнаруживаться в немиелинизированном окончании, когда тельце Пачини сдавлено. При этом возникает короткий залп импульсов в сенсорном волокне, который угасает за 1 - 2 секунды до нуля или очень малой частоты. Предполагают, что основной функцией пачиниевых телец кожи является детектирование вибрации. Частоты вибраций, на которые они реагируют, находятся в диапазоне 70 - 1000 Гц. Наиболее чувствительны они, однако, в диапазоне 200 - 400 Гц, где деформация кожи всего на 1 мкм является достаточным стимулом. разрез тельца Пачиниева: 1 - внутренняя колбочка; 2 - нервное волокно
 Кожные рецепторы: 1 — тельце Мейснера; 2 — диски Меркеля; 3 — тельце Паччини; 4 — рецептор волосяного фолликула; 5 — тактильный диск (тельце Пинкуса—Игго); 6 — окончание Руффини
Кожные рецепторы: 1 — тельце Мейснера; 2 — диски Меркеля; 3 — тельце Паччини; 4 — рецептор волосяного фолликула; 5 — тактильный диск (тельце Пинкуса—Игго); 6 — окончание Руффини
, подробно изучены русским гистологом А.") Тельца Мейснера — открыты немецким ученым Г. Мейснером (18291905), подробно изучены русским гистологом А. С. Догелем (1892). Это маленькие рецепторы, их длина 40 -180 мкм, а ширина 30 -60 мкм. Представляют собой слоистую структуру с нервным окончанием, проходящим между слоями. Снаружи тельце покрыто соединительнотканной капсулой. Располагаются в дерме. В зависимости от размера и упаковки глиальных клеток вокруг нервного отростка имеется несколько типов инкапсулированных телец. Капсула прикреплена к вышележащим слоям эпителия коллагеновыми волокнами.
Тельца Мейснера — открыты немецким ученым Г. Мейснером (18291905), подробно изучены русским гистологом А. С. Догелем (1892). Это маленькие рецепторы, их длина 40 -180 мкм, а ширина 30 -60 мкм. Представляют собой слоистую структуру с нервным окончанием, проходящим между слоями. Снаружи тельце покрыто соединительнотканной капсулой. Располагаются в дерме. В зависимости от размера и упаковки глиальных клеток вокруг нервного отростка имеется несколько типов инкапсулированных телец. Капсула прикреплена к вышележащим слоям эпителия коллагеновыми волокнами.
 Это обеспечивает эффективную механическую связь между поверхностью кожи и органом чувства. Нервные окончания образуют в капсуле спираль, ветви которой изолированы друг от друга оболочками из шванновских клеток, и стимулируются движением кожи, в частности, вибрацией. Частотный диапазон этих вибрационных стимулов ниже, чем диапазон, характерный для телец Пачини: тельца Мейснера предпочтительно отвечают на стимулы, поступающие с частотой примерно 30 -40 Гц.
Это обеспечивает эффективную механическую связь между поверхностью кожи и органом чувства. Нервные окончания образуют в капсуле спираль, ветви которой изолированы друг от друга оболочками из шванновских клеток, и стимулируются движением кожи, в частности, вибрацией. Частотный диапазон этих вибрационных стимулов ниже, чем диапазон, характерный для телец Пачини: тельца Мейснера предпочтительно отвечают на стимулы, поступающие с частотой примерно 30 -40 Гц.
 Это рецепторы давления. У человека этих клеток больше всего в коже пальцев рук и ног. Тельца Мейснера являются быстроадаптирующимися. Обладают малыми рецептивными полями, т. е. представляют тонкую чувствительность.
Это рецепторы давления. У человека этих клеток больше всего в коже пальцев рук и ног. Тельца Мейснера являются быстроадаптирующимися. Обладают малыми рецептивными полями, т. е. представляют тонкую чувствительность.
 — немецкий врачанатом, гистолог и физиолог.") Тельца Меркеля Меркель Фридрих Зигмунд (1845 — 1919) — немецкий врачанатом, гистолог и физиолог. Ректор университета в Ростоке. Тельца Меркеля — рецепторные клетки, окруженные тончайшими окончаниями чувствительных нервов. Открыты в 1875 году немецким гистологом Ф. Меркелем и подробно изучены А. С. Догелем (1903). Находятся в глубоких слоях эпидермиса кожи. Эти клетки найдены в особо чувствительных участках кожи, например в губах. У оснований клеток Меркеля располагаются дисковидные окончания сенсорных аксонов - диски Меркеля. Группа из 10 -20 клеток Меркеля образует синаптические контакты с окончаниями одного сенсорного аксона.
Тельца Меркеля Меркель Фридрих Зигмунд (1845 — 1919) — немецкий врачанатом, гистолог и физиолог. Ректор университета в Ростоке. Тельца Меркеля — рецепторные клетки, окруженные тончайшими окончаниями чувствительных нервов. Открыты в 1875 году немецким гистологом Ф. Меркелем и подробно изучены А. С. Догелем (1903). Находятся в глубоких слоях эпидермиса кожи. Эти клетки найдены в особо чувствительных участках кожи, например в губах. У оснований клеток Меркеля располагаются дисковидные окончания сенсорных аксонов - диски Меркеля. Группа из 10 -20 клеток Меркеля образует синаптические контакты с окончаниями одного сенсорного аксона.
 Клетки Меркеля реагируют на внезапные смещения кожи, например при поглаживании. Являются медленноадаптирующимися (регистрируют продолжительность давления). Диски Меркеля имеют точечные рецептивные поля.
Клетки Меркеля реагируют на внезапные смещения кожи, например при поглаживании. Являются медленноадаптирующимися (регистрируют продолжительность давления). Диски Меркеля имеют точечные рецептивные поля.
 Тельца Руффини лежат в соединительнотканной части кожи, они имеют вид веретеновидных структур. Войдя в капсулу, нервное волокно теряет миелин и разветвляется на множество ветвей, которые заканчиваются колбообразными вздутиями, окруженными глиальными клетками. Окончания плотно прилегают к коллагеновым волокнам, формирующим основу тельца. Тельца Руффини являются механорецепторами, они также воспринимают тепло и служат проприорецепторами. Активируются при растяжении кожи, являются медленноадаптирующимися рецепторами.
Тельца Руффини лежат в соединительнотканной части кожи, они имеют вид веретеновидных структур. Войдя в капсулу, нервное волокно теряет миелин и разветвляется на множество ветвей, которые заканчиваются колбообразными вздутиями, окруженными глиальными клетками. Окончания плотно прилегают к коллагеновым волокнам, формирующим основу тельца. Тельца Руффини являются механорецепторами, они также воспринимают тепло и служат проприорецепторами. Активируются при растяжении кожи, являются медленноадаптирующимися рецепторами.
 Свободные нервные окончания представлены конечными разветвлениями дендритов сенсорных нейронов. Они теряют миелин, проникают между клетками эпителия и располагаются в эпидермисе и дерме. В некоторых случаях конечные разветвления осевого цилиндра окутывают измененные эпителиальные клетки, образуя осязательные мениски. Несвободные нервные окончания состоят не только из ветвлений волокна, потерявшего миелин, но и из клеток глии.
Свободные нервные окончания представлены конечными разветвлениями дендритов сенсорных нейронов. Они теряют миелин, проникают между клетками эпителия и располагаются в эпидермисе и дерме. В некоторых случаях конечные разветвления осевого цилиндра окутывают измененные эпителиальные клетки, образуя осязательные мениски. Несвободные нервные окончания состоят не только из ветвлений волокна, потерявшего миелин, но и из клеток глии.
 Свободные нервные окончания иннервируют волосяные фолликулы волос (покрывающих большую часть тела человека, а также грубых волос, растущих на голове, в подмышечных впадинах, у мужчин - на лице). Свободные нервные окончания волосяных фолликулов являются механорецепторами и возбуждаются при смещении волос или их подергивании. Типы рецепторов кожи: А — тельце Пачини; Б — тельце Мейснера; В — нервное сплетение у основания волосяной луковицы; Г — колба Краузе; Д — нервное сплетение роговой оболочки.
Свободные нервные окончания иннервируют волосяные фолликулы волос (покрывающих большую часть тела человека, а также грубых волос, растущих на голове, в подмышечных впадинах, у мужчин - на лице). Свободные нервные окончания волосяных фолликулов являются механорецепторами и возбуждаются при смещении волос или их подергивании. Типы рецепторов кожи: А — тельце Пачини; Б — тельце Мейснера; В — нервное сплетение у основания волосяной луковицы; Г — колба Краузе; Д — нервное сплетение роговой оболочки.
 Другая разновидность свободных нервных окончаний имеется в эпидермисе и в сосочковом слое дермы, большинство из них являются ноцицепторами или терморецепторами, но некоторые принадлежат к механорецепторам, которые специфически реагируют на слабое околопороговое раздражение. Предполагается, что при раздражении этой разновидности рецепторов возникают ощущения щекотки и зуда.
Другая разновидность свободных нервных окончаний имеется в эпидермисе и в сосочковом слое дермы, большинство из них являются ноцицепторами или терморецепторами, но некоторые принадлежат к механорецепторам, которые специфически реагируют на слабое околопороговое раздражение. Предполагается, что при раздражении этой разновидности рецепторов возникают ощущения щекотки и зуда.
 Типы рецепторов кожи: А — тельце Пачини; Б — тельце Мейснера; В — нервное сплетение у основания волосяной луковицы; Г — колба Краузе; Д — нервное сплетение роговой оболочки. Тонкость различения деталей тактильных стимулов зависит от плотности расположения рецепторов в коже. Так, на губах, языке, кончиках пальцев плотность расположения рецепторов наибольшая и соответственно различение тактильных особенностей наилучшее.
Типы рецепторов кожи: А — тельце Пачини; Б — тельце Мейснера; В — нервное сплетение у основания волосяной луковицы; Г — колба Краузе; Д — нервное сплетение роговой оболочки. Тонкость различения деталей тактильных стимулов зависит от плотности расположения рецепторов в коже. Так, на губах, языке, кончиках пальцев плотность расположения рецепторов наибольшая и соответственно различение тактильных особенностей наилучшее.
 Характеристики рецепторов Исследования уровня тактильной чувствительности можно проводить с помощью «волосков Фрея» , а пространственных порогов, которые характеризуют плотность распределения тактильных рецепторов, – «циркулем Вебера» . При использовании метода Фрея применяется набор специальных градуированных волосков и щетинок, прикрепленных под прямым углом к ручке. Метод позволяет определить количество чувствительных точек на 1 см 2 поверхности и установить порог раздражения точек. Измерение кожной чувствительности. А. Набором волосков Фрея разной толщины можно измерить порог восприятия прикосновения. Результаты выражаются количественно путем калибровки на шкале чувствительности силы, при которой волосок начинает сгибаться. Б. Электромеханический прибор для точно градуированной механической стимуляции кожи. Ток, проходящий через катушку в магнитном поле, вызывает ее смещение на строго определенную величину (как в громкоговорителе). Электрические волны произвольно выбранной формы преобразуются в движения стерженька
Характеристики рецепторов Исследования уровня тактильной чувствительности можно проводить с помощью «волосков Фрея» , а пространственных порогов, которые характеризуют плотность распределения тактильных рецепторов, – «циркулем Вебера» . При использовании метода Фрея применяется набор специальных градуированных волосков и щетинок, прикрепленных под прямым углом к ручке. Метод позволяет определить количество чувствительных точек на 1 см 2 поверхности и установить порог раздражения точек. Измерение кожной чувствительности. А. Набором волосков Фрея разной толщины можно измерить порог восприятия прикосновения. Результаты выражаются количественно путем калибровки на шкале чувствительности силы, при которой волосок начинает сгибаться. Б. Электромеханический прибор для точно градуированной механической стимуляции кожи. Ток, проходящий через катушку в магнитном поле, вызывает ее смещение на строго определенную величину (как в громкоговорителе). Электрические волны произвольно выбранной формы преобразуются в движения стерженька
, пространственным и временным разрешениями. Абсолютный порог") Тактильная чувствительность характеризуется порогом интенсивности (абсолютным и относительным), пространственным и временным разрешениями. Абсолютный порог тактильной чувствительности определяется давлением, при котором испытуемый чувствует воздействие. Вот соответствующие данные для разных участков тела (в мг/мм). Кончик языка. . . . . 2 Кончик пальцев. . . . . 3 Поверхность десны. . . . 4 Тыльная сторона ладони. . . . . 5 Ладонная поверхность предплечья. . 8 Тыл кисти. . . 12 Икры ног. . . 15 Поверхность живота. . . . 26 Тыльная поверхность предплечья. . . 35 Поясница. . . 48 Плотная часть подошвы. . . . . 250
Тактильная чувствительность характеризуется порогом интенсивности (абсолютным и относительным), пространственным и временным разрешениями. Абсолютный порог тактильной чувствительности определяется давлением, при котором испытуемый чувствует воздействие. Вот соответствующие данные для разных участков тела (в мг/мм). Кончик языка. . . . . 2 Кончик пальцев. . . . . 3 Поверхность десны. . . . 4 Тыльная сторона ладони. . . . . 5 Ладонная поверхность предплечья. . 8 Тыл кисти. . . 12 Икры ног. . . 15 Поверхность живота. . . . 26 Тыльная поверхность предплечья. . . 35 Поясница. . . 48 Плотная часть подошвы. . . . . 250
 и") Клинические тактильные тесты. При рутинном клиническом тестировании к коже слегка прикасаются (например, ваткой) и пациента спрашивают о том, что он ощущает и где находится раздражитель. Пространственное разрешение оценивается по распознаванию цифр или букв разной величины, вычерчиваемых на коже тупым стержнем. Вибрационная чувствительность определяется электромагнитным вибратором. Врач измеряет как абсолютный порог ощущения вибрации, так и порог различения изменений в ее частоте. Двухточечный порог кожи. А. Кончики циркуля, разведенные на разные расстояния, неоднократно прикасаются к коже. Двухточечный порог–наименьшее расстояние, при котором испытуемый может определить, что стимулируются две точки, а не одна. Б. Изменчивость двухточечного порога кожи в зависимости от участка тела человека (с изменениями по [22]; данные Е. Н. Weber, Archiv fur Anatomie, Physiologic und wissenschaftliche Medizin, 1835, p. 152)
Клинические тактильные тесты. При рутинном клиническом тестировании к коже слегка прикасаются (например, ваткой) и пациента спрашивают о том, что он ощущает и где находится раздражитель. Пространственное разрешение оценивается по распознаванию цифр или букв разной величины, вычерчиваемых на коже тупым стержнем. Вибрационная чувствительность определяется электромагнитным вибратором. Врач измеряет как абсолютный порог ощущения вибрации, так и порог различения изменений в ее частоте. Двухточечный порог кожи. А. Кончики циркуля, разведенные на разные расстояния, неоднократно прикасаются к коже. Двухточечный порог–наименьшее расстояние, при котором испытуемый может определить, что стимулируются две точки, а не одна. Б. Изменчивость двухточечного порога кожи в зависимости от участка тела человека (с изменениями по [22]; данные Е. Н. Weber, Archiv fur Anatomie, Physiologic und wissenschaftliche Medizin, 1835, p. 152)
 Пространственное разрешение определяется минимальным расстоянием между двумя точками, прикосновения к которым ощущаются как отдельные. Расстояние для разных участков тела составляет (в мм): Кончик языка. . . . 1 Кожа сгибателя пальца. . . 2 Губы. . . . . 4, 5 Кожа разгибателя пальца. . 7 Затылок. . . . . 27 Шея. . . 54 Бедро, спина. . . . 67 Пространственное разрешение зависит не только от расположения рецепторов: если раздражать две точки не одновременно, а по очереди, то разрешение возрастает. Следовательно, оно зависит и от обработки сигналов.
Пространственное разрешение определяется минимальным расстоянием между двумя точками, прикосновения к которым ощущаются как отдельные. Расстояние для разных участков тела составляет (в мм): Кончик языка. . . . 1 Кожа сгибателя пальца. . . 2 Губы. . . . . 4, 5 Кожа разгибателя пальца. . 7 Затылок. . . . . 27 Шея. . . 54 Бедро, спина. . . . 67 Пространственное разрешение зависит не только от расположения рецепторов: если раздражать две точки не одновременно, а по очереди, то разрешение возрастает. Следовательно, оно зависит и от обработки сигналов.
 Относительный порог интенсивности для тактильных ощущений: человек замечает, что давление изменилось, если это изменение составило не меньше 6%. Наибольший же разрыв между порогом чувствительности и порогом боли имеет место на кончиках пальцев, которые, будучи весьма чувствительными к прикосновению, относительно мало чувствительны к боли.
Относительный порог интенсивности для тактильных ощущений: человек замечает, что давление изменилось, если это изменение составило не меньше 6%. Наибольший же разрыв между порогом чувствительности и порогом боли имеет место на кончиках пальцев, которые, будучи весьма чувствительными к прикосновению, относительно мало чувствительны к боли.
 Классификация кожных механорецепторов по скорости адаптации и адекватным стимулам
Классификация кожных механорецепторов по скорости адаптации и адекватным стимулам
 Количество и плотность размещения рецепторов: - Общее количество тактильных рецепторов оценивается в 10 миллионов штук, сгруппированных в 1 миллион афферентов. - Количество свободных афферентных окончаний в коже: 50% от общего количества кожных афферентов. - Плотность размещения свободных нервных окончаний в коже: 170 шт/см 2 Размеры и прочие структурные параметры: - Тельца Мейснера: 50 -140 мкм - Колбы Крауза: 40 -150 мкм - Тельца Руффини: 1 -2 мм - Тельца Фатера-Пачини: 0. 5 -5 мм - Нервно-сухожильные веретёна: - Размеры: 0. 5 -1 мм - Количество мышечных волокон в веретене: 15 штук - Нервно-мышечные веретёна - Количество мышечных волокон в веретене: 1 -8 штук - Диаметр выходящих нервных волокон: 17 мкм (первичные) и 8 мкм (вторичные)
Количество и плотность размещения рецепторов: - Общее количество тактильных рецепторов оценивается в 10 миллионов штук, сгруппированных в 1 миллион афферентов. - Количество свободных афферентных окончаний в коже: 50% от общего количества кожных афферентов. - Плотность размещения свободных нервных окончаний в коже: 170 шт/см 2 Размеры и прочие структурные параметры: - Тельца Мейснера: 50 -140 мкм - Колбы Крауза: 40 -150 мкм - Тельца Руффини: 1 -2 мм - Тельца Фатера-Пачини: 0. 5 -5 мм - Нервно-сухожильные веретёна: - Размеры: 0. 5 -1 мм - Количество мышечных волокон в веретене: 15 штук - Нервно-мышечные веретёна - Количество мышечных волокон в веретене: 1 -8 штук - Диаметр выходящих нервных волокон: 17 мкм (первичные) и 8 мкм (вторичные)
 Скорость передачи: - Психофизическая оценка ёмкости тактильных каналов: 5 бит/c - Скорость передачи импульсов из свободных нервных окончаний: 1 м/c (С-волокна, IV тип) - Скорость передачи импульсов из инкапсулированных рецепторов: 50 м/c (А-волокна, II тип) Разрешающая способность и пороги ощущений: - Порог ощущения кожных рецепторов при надавливании: 10 мкм - Одновременный пространственный порог кожных рецепторов: - На губах и кончиках пальцев: 1 -3 мм - На спине, плечах, бёдрах: 50 -100 мм - Последовательный пространственный порог кожных рецепторов: - На губах и кончиках пальцев: 1 мм - На спине, плечах, бёдрах: 10 -20 мм - Порог ощущения вибрации: 150 -300 Гц - Минимальная ощущаемая амплитуда вибрации: 1 мкм
Скорость передачи: - Психофизическая оценка ёмкости тактильных каналов: 5 бит/c - Скорость передачи импульсов из свободных нервных окончаний: 1 м/c (С-волокна, IV тип) - Скорость передачи импульсов из инкапсулированных рецепторов: 50 м/c (А-волокна, II тип) Разрешающая способность и пороги ощущений: - Порог ощущения кожных рецепторов при надавливании: 10 мкм - Одновременный пространственный порог кожных рецепторов: - На губах и кончиках пальцев: 1 -3 мм - На спине, плечах, бёдрах: 50 -100 мм - Последовательный пространственный порог кожных рецепторов: - На губах и кончиках пальцев: 1 мм - На спине, плечах, бёдрах: 10 -20 мм - Порог ощущения вибрации: 150 -300 Гц - Минимальная ощущаемая амплитуда вибрации: 1 мкм
: - Тельца Руффини и колбы Краузе помимо механических воздействий детектируют") Модальность (специфичность к воздействиям): - Тельца Руффини и колбы Краузе помимо механических воздействий детектируют температуру, т. е. полимодальны. Тельца Мейснера могут ощущать вибрацию. Также полимодальны неинкапсулированные свободные афференты. - Остальные рецепторы унимодальны. Рецепторные поля (площадь рецепции): - Для тактильных дисков площадь равна анатомическим размерам. - Для дисков Меркеля это аггломерация из 30 -50 дисков, обслуживаемых одним нервным волокном. - Для телец Фатера-Паччини эта область больше анатомически размером.
Модальность (специфичность к воздействиям): - Тельца Руффини и колбы Краузе помимо механических воздействий детектируют температуру, т. е. полимодальны. Тельца Мейснера могут ощущать вибрацию. Также полимодальны неинкапсулированные свободные афференты. - Остальные рецепторы унимодальны. Рецепторные поля (площадь рецепции): - Для тактильных дисков площадь равна анатомическим размерам. - Для дисков Меркеля это аггломерация из 30 -50 дисков, обслуживаемых одним нервным волокном. - Для телец Фатера-Паччини эта область больше анатомически размером.
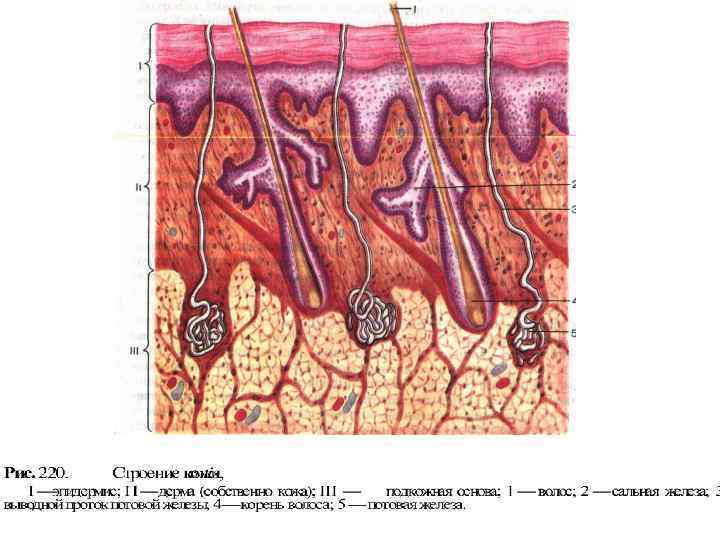
 Рецепторный аппарат кожи 1 – нервное сплетение луковицы волоса; 2 – свободные нервные окончания; 3 – диски Меркеля; 4 – тельца Мейснера; 5 – тельца Пачини; 6 – тельца Руффини (рецепторы тепла); 7 – колбочки Краузе (рецепторы холода); 8 – эпидермис; 9 – дерма.
Рецепторный аппарат кожи 1 – нервное сплетение луковицы волоса; 2 – свободные нервные окончания; 3 – диски Меркеля; 4 – тельца Мейснера; 5 – тельца Пачини; 6 – тельца Руффини (рецепторы тепла); 7 – колбочки Краузе (рецепторы холода); 8 – эпидермис; 9 – дерма.
 Проводниковый отдел От большинства механорецепторов в спинной мозг информация поступает в ЦНС по А-волокнам и лишь от рецепторов щекотки – по С-волокнам. Первый нейрон находится в спинальных ганглиях. В заднем роге спинного мозга происходит первое переключение на интернейроны (второй нейрон), от них восходящий путь в составе заднего столба достигает ядер заднего столба в продолговатом мозге (третий нейрон), где происходит второе переключение, далее через медиальную петлю путь следует к ядрам зрительного бугра (четвертый нейрон), центральные отростки нейронов зрительного бугра идут в кору больших полушарий.
Проводниковый отдел От большинства механорецепторов в спинной мозг информация поступает в ЦНС по А-волокнам и лишь от рецепторов щекотки – по С-волокнам. Первый нейрон находится в спинальных ганглиях. В заднем роге спинного мозга происходит первое переключение на интернейроны (второй нейрон), от них восходящий путь в составе заднего столба достигает ядер заднего столба в продолговатом мозге (третий нейрон), где происходит второе переключение, далее через медиальную петлю путь следует к ядрам зрительного бугра (четвертый нейрон), центральные отростки нейронов зрительного бугра идут в кору больших полушарий.
 1 — пучок Голля; 2 — пучок Бурдаха; 3 — задний корешок; 4 — передний корешок; 5 — спиноталамический тракт (проведение болевой чувствительности); 6 — двигательные аксоны; 7 — симпатические аксоны; 8 — передний рог; 9 — проприоспинальный путь; 10 — задний рог; И — висцерорецепторы; 12 — проприорецепторы; 13 — терморецепторы; 14 — ноцицепторы; 15 — механорецепторы Все кожные рецепторы являются окончаниями псевдоуниполярных чувствительных нейронов, расположенных в спинномозговых ганглиях. По афферентным волокнам (дендритам) этих нейронов информация поступает сначала к телу нейрона, а затем по его аксону в задние рога соответствующего сегмента спинного мозга.
1 — пучок Голля; 2 — пучок Бурдаха; 3 — задний корешок; 4 — передний корешок; 5 — спиноталамический тракт (проведение болевой чувствительности); 6 — двигательные аксоны; 7 — симпатические аксоны; 8 — передний рог; 9 — проприоспинальный путь; 10 — задний рог; И — висцерорецепторы; 12 — проприорецепторы; 13 — терморецепторы; 14 — ноцицепторы; 15 — механорецепторы Все кожные рецепторы являются окончаниями псевдоуниполярных чувствительных нейронов, расположенных в спинномозговых ганглиях. По афферентным волокнам (дендритам) этих нейронов информация поступает сначала к телу нейрона, а затем по его аксону в задние рога соответствующего сегмента спинного мозга.
 В каждый задний корешок спинного мозга входят афференты, собирающие информацию с определенного участка кожи. Такой участок кожи называется дерматомом. Дерматомы организма человека
В каждый задний корешок спинного мозга входят афференты, собирающие информацию с определенного участка кожи. Такой участок кожи называется дерматомом. Дерматомы организма человека
 Дерматомы лица и шеи иннервируются тройничным и лицевым нервами. Первичная обработка сигнала производится нейронами задних рогов сегмента спинного мозга (или соответствующими ядрами черепно-мозговых нервов). От этих нейронов информация может поступать к мотонейронам и вегетативным (симпатическим) нейронам своего сегмента; далее короткими путями к соседним сегментам и, наконец, в протяженные восходящие пути спинного мозга (Голля и Бурдаха для тактильных и температурных воздействий и спиноталамические для болевых воздействий).
Дерматомы лица и шеи иннервируются тройничным и лицевым нервами. Первичная обработка сигнала производится нейронами задних рогов сегмента спинного мозга (или соответствующими ядрами черепно-мозговых нервов). От этих нейронов информация может поступать к мотонейронам и вегетативным (симпатическим) нейронам своего сегмента; далее короткими путями к соседним сегментам и, наконец, в протяженные восходящие пути спинного мозга (Голля и Бурдаха для тактильных и температурных воздействий и спиноталамические для болевых воздействий).
 По трактам Голля и Бурдаха сигналы достигают одноименных ядер продолговатого мозга, затем переключаются в таламусе (вентробазальное ядро) и соматотопически проецируются в контрлатеральную постцентральную извилину. Спиноталамические пути, к которым присоединяются болевые афференты тройничного и лицевого нервов, переключаются в таламусе и проецируются также в постцентральную кору.
По трактам Голля и Бурдаха сигналы достигают одноименных ядер продолговатого мозга, затем переключаются в таламусе (вентробазальное ядро) и соматотопически проецируются в контрлатеральную постцентральную извилину. Спиноталамические пути, к которым присоединяются болевые афференты тройничного и лицевого нервов, переключаются в таламусе и проецируются также в постцентральную кору.
 Схема проводящих путей тактильной чувствительности рецепторы пучок Голля, Бурдаха Спиной мозг Ядра пучка Голля, Бурдаха Вентральные ядра Продолговатый мозг Зрительный бугор Задняя центр. Извилина Кора
Схема проводящих путей тактильной чувствительности рецепторы пучок Голля, Бурдаха Спиной мозг Ядра пучка Голля, Бурдаха Вентральные ядра Продолговатый мозг Зрительный бугор Задняя центр. Извилина Кора
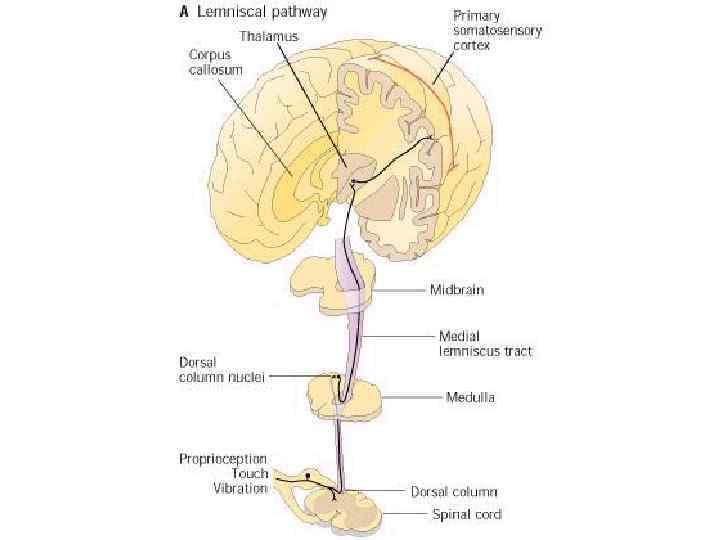
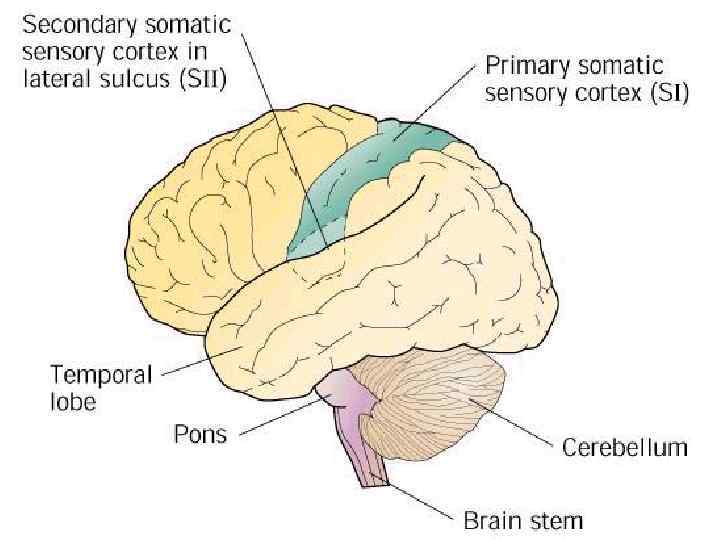
 Центральный отдел тактильного анализатора локализуется в 1 и II зонах соматосенсорной области коры большого мозга (задняя центральная извилина).
Центральный отдел тактильного анализатора локализуется в 1 и II зонах соматосенсорной области коры большого мозга (задняя центральная извилина).