

lekzia11-функции отделов мозга.ppt
- Количество слайдов: 70
 Лекция 11. ФУНКЦИИ ПРОДОЛГОВАТОГО МОЗГА, МОСТА, СРЕДНЕГО МОЗГА Дыхательный центр. Сосудодвигательный центр. Центры, обеспечивающие врожденное пищевое поведение. Слуховые и вестибулярные ядра. Центры сна и бодрствования, стадии сна. Средний мозг и ориентировочный рефлекс. Экстрапирамидная система.
Лекция 11. ФУНКЦИИ ПРОДОЛГОВАТОГО МОЗГА, МОСТА, СРЕДНЕГО МОЗГА Дыхательный центр. Сосудодвигательный центр. Центры, обеспечивающие врожденное пищевое поведение. Слуховые и вестибулярные ядра. Центры сна и бодрствования, стадии сна. Средний мозг и ориентировочный рефлекс. Экстрапирамидная система.
 Эпифиз Таламус Верхние холмики Средний мозг Нижние холмики Мост Ножки мозжечка Дно 4 -го желудочка Продолговатый мозг и мост выполняют ряд функций. Здесь находятся центры: дыхательный, сосудодвигательный, пищевого поведения и др.
Эпифиз Таламус Верхние холмики Средний мозг Нижние холмики Мост Ножки мозжечка Дно 4 -го желудочка Продолговатый мозг и мост выполняют ряд функций. Здесь находятся центры: дыхательный, сосудодвигательный, пищевого поведения и др.
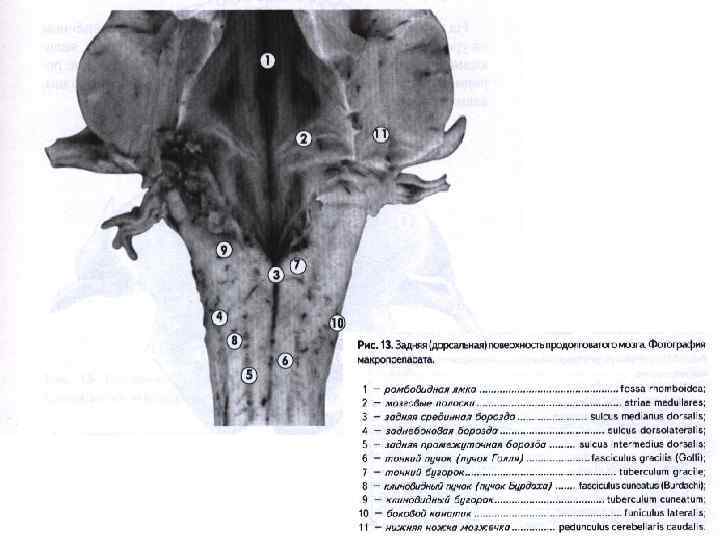
,") От продолговатого мозга отходят IX-XII пары черепномозговых нервов: IX — языкоглоточный нерв (n. glossopharingeus), X — блуждающий нерв (п. vagus), XI — добавочный нерв (п. accessorius), XII — подъязычный нерв (п. hypoglossus).
От продолговатого мозга отходят IX-XII пары черепномозговых нервов: IX — языкоглоточный нерв (n. glossopharingeus), X — блуждающий нерв (п. vagus), XI — добавочный нерв (п. accessorius), XII — подъязычный нерв (п. hypoglossus).
 и проводящих") Поперечный разрез продолговатого мозга Левая половина — схема расположения ядер (обозначены светлым) и проводящих путей (обозначены темным): 1 — задний продольный пучок; 2 — шов; 3 — пирамида; 4 — корешок подъязычного нерва; 5 — медиальная петля (осязательный путь); 6 — ядро оливы; 7 — ретикулярная формация; 8 — спиноталамический тракт; 9 — двойное ядро; 10— корешок блуждающего нерва; 11 — дорсальный продольный пучок; 12 — нижние ножки мозжечка; 13 — ядро Дейтерса; 14 — одиночный (солитарный) тракт и его ядро; 15 — верхнее вестибулярное ядро; 16 — дорсальное ядро блуждающего нерва; 17 — ядро подъязычного нерва
Поперечный разрез продолговатого мозга Левая половина — схема расположения ядер (обозначены светлым) и проводящих путей (обозначены темным): 1 — задний продольный пучок; 2 — шов; 3 — пирамида; 4 — корешок подъязычного нерва; 5 — медиальная петля (осязательный путь); 6 — ядро оливы; 7 — ретикулярная формация; 8 — спиноталамический тракт; 9 — двойное ядро; 10— корешок блуждающего нерва; 11 — дорсальный продольный пучок; 12 — нижние ножки мозжечка; 13 — ядро Дейтерса; 14 — одиночный (солитарный) тракт и его ядро; 15 — верхнее вестибулярное ядро; 16 — дорсальное ядро блуждающего нерва; 17 — ядро подъязычного нерва
 Оливные ядра связаны с зубчатыми ядрами мозжечка и являются промежуточными ядрами равновесия. Оливные ядра достигают наибольшей выраженности у человека в связи с вертикальным положением тела, так как прямохождение нуждается в совершенном вестибулярном аппарате.
Оливные ядра связаны с зубчатыми ядрами мозжечка и являются промежуточными ядрами равновесия. Оливные ядра достигают наибольшей выраженности у человека в связи с вертикальным положением тела, так как прямохождение нуждается в совершенном вестибулярном аппарате.
, образованная переплетением нервных волокон и") В продолговатом мозге находится ретикулярная формация (reticulum – сеть), образованная переплетением нервных волокон и лежащих между ними нервных клеток. Двигательная часть ретикулярной формации содержит центры, обеспечивающие контроль за кровообращением и дыханием. 7 - ретикулярная формация
В продолговатом мозге находится ретикулярная формация (reticulum – сеть), образованная переплетением нервных волокон и лежащих между ними нервных клеток. Двигательная часть ретикулярной формации содержит центры, обеспечивающие контроль за кровообращением и дыханием. 7 - ретикулярная формация
 ретикулярная формация
ретикулярная формация
 олива пирамида
олива пирамида
 ретикулярная формация Ретикулоспинальный тракт Через него ретикулярная формация оказывает влияние как на двигательную деятельность спинного мозга (осуществление спинальных рефлексов), так и на вегетативную регуляцию (сосудодвигательная, дыхательная, пищеварительная функции).
ретикулярная формация Ретикулоспинальный тракт Через него ретикулярная формация оказывает влияние как на двигательную деятельность спинного мозга (осуществление спинальных рефлексов), так и на вегетативную регуляцию (сосудодвигательная, дыхательная, пищеварительная функции).
 Дыхательный центр Регуляция дыхательной функции осуществляется иерархической системой нервных структур, расположенных на разных этажах нервной системы: - мотонейроны спинного мозга; - продолговатый мозг; - варолиев мост; - средний мозг; - гипоталамус; - лимбико-ретикулярный комплекс; - кора больших полушарий. В продолговатом мозге выделяют дыхательный центр совокупность нейронов, способных генерировать дыхательный ритм
Дыхательный центр Регуляция дыхательной функции осуществляется иерархической системой нервных структур, расположенных на разных этажах нервной системы: - мотонейроны спинного мозга; - продолговатый мозг; - варолиев мост; - средний мозг; - гипоталамус; - лимбико-ретикулярный комплекс; - кора больших полушарий. В продолговатом мозге выделяют дыхательный центр совокупность нейронов, способных генерировать дыхательный ритм
, Е –") Схема расположения дыхательных нейронов продолг. мозга и моста: I – инспираторные (вдох), Е – экспираторные (выдох).
Схема расположения дыхательных нейронов продолг. мозга и моста: I – инспираторные (вдох), Е – экспираторные (выдох).
 Импульсы от нейронов продолговатого мозга идут к мотонейронам спинного мозга, которые посылают информацию к дыхательным мышцам по диафрагмальным и межреберным нервам. Центр диафрагмальных нервов находится в основном в 3 -4 -ом шейных сегментах спинного мозга. Центры межреберных нервов, иннервирующих мускулатуру грудной клетки, локализуются в грудном отделе спинного мозга.
Импульсы от нейронов продолговатого мозга идут к мотонейронам спинного мозга, которые посылают информацию к дыхательным мышцам по диафрагмальным и межреберным нервам. Центр диафрагмальных нервов находится в основном в 3 -4 -ом шейных сегментах спинного мозга. Центры межреберных нервов, иннервирующих мускулатуру грудной клетки, локализуются в грудном отделе спинного мозга.
 Вышележащие отделы ЦНС, принимающие участие в регуляции дыхания В регуляции дыхания принимают участие также варолиев мост, средний мозг, гипоталамус, лимбикоретикулярный комплекс, кора большого мозга. Периодичность в работе дыхательного центра продолговатого мозга может обеспечиваться за счет регулирующих влияний со стороны т. н. пневмотаксического центра, который находится в области варолиева моста и состоит из двух типов нейронов - инспираторных и экспираторных.
Вышележащие отделы ЦНС, принимающие участие в регуляции дыхания В регуляции дыхания принимают участие также варолиев мост, средний мозг, гипоталамус, лимбикоретикулярный комплекс, кора большого мозга. Периодичность в работе дыхательного центра продолговатого мозга может обеспечиваться за счет регулирующих влияний со стороны т. н. пневмотаксического центра, который находится в области варолиева моста и состоит из двух типов нейронов - инспираторных и экспираторных.
 Таламус Эпифиз Верхние холмики Средний мозг Нижние холмики Мост Ножки мозжечка Дно 4 -го желудочка Пневмотаксический центр периодически затормаживает инспираторную часть дыхательного центра и стимулирует экспираторные нейроны, осуществляя таким образом прекращение вдоха и начало выдоха. Мост играет важную роль в регуляции продолжительности фаз вдоха, выдоха и паузы между ними. Нейроны моста при взаимодействии с нейронами продолговатого мозга обеспечивают нормальный цикл дыхания.
Таламус Эпифиз Верхние холмики Средний мозг Нижние холмики Мост Ножки мозжечка Дно 4 -го желудочка Пневмотаксический центр периодически затормаживает инспираторную часть дыхательного центра и стимулирует экспираторные нейроны, осуществляя таким образом прекращение вдоха и начало выдоха. Мост играет важную роль в регуляции продолжительности фаз вдоха, выдоха и паузы между ними. Нейроны моста при взаимодействии с нейронами продолговатого мозга обеспечивают нормальный цикл дыхания.
 Средний мозг играет важную роль в регуляции тонуса всей мускулатуры организма, в том числе и дыхательной. Гипоталамус выполняет интегрирующую роль в вегетативном обеспечении соматической деятельности, в том числе участвует в регуляции частоты и глубины дыхания при физической деятельности, повышении температуры внешней и внутренней среды.
Средний мозг играет важную роль в регуляции тонуса всей мускулатуры организма, в том числе и дыхательной. Гипоталамус выполняет интегрирующую роль в вегетативном обеспечении соматической деятельности, в том числе участвует в регуляции частоты и глубины дыхания при физической деятельности, повышении температуры внешней и внутренней среды.
 Кора большого мозга – произвольное изменение частоты и глубины дыхания. Благодаря коре большого мозга при выполнении физических упражнений интенсивность дыхания становится адекватной потребностям организма (более экономное дыхание).
Кора большого мозга – произвольное изменение частоты и глубины дыхания. Благодаря коре большого мозга при выполнении физических упражнений интенсивность дыхания становится адекватной потребностям организма (более экономное дыхание).
 Автоматия дыхательного центра Основная часть нейронов дыхательного центра в продолговатом мозге относится к ретикулярной формации, эти нейроны обладают свойством спонтанной активности - способность к автоматизму. Даже при отсутствии афферентных воздействий активность этих нейронов характеризуется периодичностью, которая определяется спецификой ионных механизмов их клеточной мембраны. Саморегуляция вдоха и выдоха обеспечивается циркуляцией возбуждения в нейронах дыхательного центра.
Автоматия дыхательного центра Основная часть нейронов дыхательного центра в продолговатом мозге относится к ретикулярной формации, эти нейроны обладают свойством спонтанной активности - способность к автоматизму. Даже при отсутствии афферентных воздействий активность этих нейронов характеризуется периодичностью, которая определяется спецификой ионных механизмов их клеточной мембраны. Саморегуляция вдоха и выдоха обеспечивается циркуляцией возбуждения в нейронах дыхательного центра.
 Периодичность разрядов дыхательных нейронов может быть обусловлена также наличием взаимных тормозных связей между инспираторными и экспираторными нейронами. Тормозные связи создают реципрокность разрядов дыхательных нейронов, когда появление активности инспираторных нейронов сопровождается торможением разрядов экспираторных и наоборот. В результате реципрокного взаимодействия дыхательных нейронов происходит смена фаз дыхательного цикла.
Периодичность разрядов дыхательных нейронов может быть обусловлена также наличием взаимных тормозных связей между инспираторными и экспираторными нейронами. Тормозные связи создают реципрокность разрядов дыхательных нейронов, когда появление активности инспираторных нейронов сопровождается торможением разрядов экспираторных и наоборот. В результате реципрокного взаимодействия дыхательных нейронов происходит смена фаз дыхательного цикла.
 Автоматии дыхательного центра способствуют гуморальные влияния непосредственно на центр, главным образом, со стороны СО 2. Автоматическая деятельность нейронов дыхательного центра изменяется под влиянием импульсации, поступающей из различных афферентных систем. Самым мощным источником рефлекторной регуляции активности нейронов дыхательного аппарата является афферентная система самого дыхательного аппарата. Плевра, дыхательные пути и сама ткань легких содержит большое количество рецепторных структур (рефлексогенные зоны: хемо- и механорецепторы). Механорецепторы - активируются при растяжении или, наоборот, при сжатии легких. Импульсы от них поступают в мозг по афферентным волокнам блуждающего нерва и в значительной мере изменяют деятельность нейронов дыхательного центра.
Автоматии дыхательного центра способствуют гуморальные влияния непосредственно на центр, главным образом, со стороны СО 2. Автоматическая деятельность нейронов дыхательного центра изменяется под влиянием импульсации, поступающей из различных афферентных систем. Самым мощным источником рефлекторной регуляции активности нейронов дыхательного аппарата является афферентная система самого дыхательного аппарата. Плевра, дыхательные пути и сама ткань легких содержит большое количество рецепторных структур (рефлексогенные зоны: хемо- и механорецепторы). Механорецепторы - активируются при растяжении или, наоборот, при сжатии легких. Импульсы от них поступают в мозг по афферентным волокнам блуждающего нерва и в значительной мере изменяют деятельность нейронов дыхательного центра.
 Нейроны дыхательных центров подвержены также влиянию рецептивных зон сосудов: увеличение содержания СО 2 в крови возбуждает хеморецепторы аортальной и синокаротидной рефлексогенных зон, при этом увеличивается поток импульсов по соответствующим нервам в ЦНС, а оттуда – к дыхательной мускулатуре, что ведет к учащению и углублению дыхания. На деятельность нейронов дыхательного центра влияют различные неспецифические рефлекторные факторы. Так, любое болевое воздействие сопровождается изменением дыхательной ритмики. Погружение в холодную воду тормозит выдох, в результате чего возникает затяжной вдох. Нельзя сделать вдох одновременно с глотанием: возникновение глотательного рефлекса затормаживает деятельность дыхательного центра.
Нейроны дыхательных центров подвержены также влиянию рецептивных зон сосудов: увеличение содержания СО 2 в крови возбуждает хеморецепторы аортальной и синокаротидной рефлексогенных зон, при этом увеличивается поток импульсов по соответствующим нервам в ЦНС, а оттуда – к дыхательной мускулатуре, что ведет к учащению и углублению дыхания. На деятельность нейронов дыхательного центра влияют различные неспецифические рефлекторные факторы. Так, любое болевое воздействие сопровождается изменением дыхательной ритмики. Погружение в холодную воду тормозит выдох, в результате чего возникает затяжной вдох. Нельзя сделать вдох одновременно с глотанием: возникновение глотательного рефлекса затормаживает деятельность дыхательного центра.
 Функция дыхания теснейшим образом связана с эмоциональными процессами. Почти все эмоциональные проявления человека (смех, плач) связаны с изменением дыхания. В этом случае функция дыхательного центра изменяется импульсацией из центров ствола головного мозга, которые особенно тесно связаны с эмоциональным поведением.
Функция дыхания теснейшим образом связана с эмоциональными процессами. Почти все эмоциональные проявления человека (смех, плач) связаны с изменением дыхания. В этом случае функция дыхательного центра изменяется импульсацией из центров ствола головного мозга, которые особенно тесно связаны с эмоциональным поведением.
 Сосудодвигательный центр - совокупность нейронов, расположенных на различных уровнях центральной нервной системы и осуществляющих регуляцию сосудистого тонуса. В составе ЦНС есть следующие уровни: - спинальный; - бульбарный (продолговатый мозг); - гипоталамический; - корковый.
Сосудодвигательный центр - совокупность нейронов, расположенных на различных уровнях центральной нервной системы и осуществляющих регуляцию сосудистого тонуса. В составе ЦНС есть следующие уровни: - спинальный; - бульбарный (продолговатый мозг); - гипоталамический; - корковый.
 Роль спинного мозга в регуляции сосудистого тонуса Спинальный уровень сосудодвигательного центра открыт в 1870 г. Овсянниковым. Он осуществлял перерезку ЦНС на различных уровнях и обнаружил, что у спинального животного снижается давление (АД) крови, но затем постепенно восстанавливается, хотя не до исходного уровня, и поддерживается на постоянном уровне. Спинной мозг играет определенную роль в регуляции сосудистого тонуса, но не имеет большого самостоятельного значения. Он передаёт импульсы от выше лежащих отделов сосудодвигательного центра.
Роль спинного мозга в регуляции сосудистого тонуса Спинальный уровень сосудодвигательного центра открыт в 1870 г. Овсянниковым. Он осуществлял перерезку ЦНС на различных уровнях и обнаружил, что у спинального животного снижается давление (АД) крови, но затем постепенно восстанавливается, хотя не до исходного уровня, и поддерживается на постоянном уровне. Спинной мозг играет определенную роль в регуляции сосудистого тонуса, но не имеет большого самостоятельного значения. Он передаёт импульсы от выше лежащих отделов сосудодвигательного центра.
 Таламус Эпифиз Верхние холмики Средний мозг Нижние холмики Мост Ножки мозжечка Дно 4 -го желудочка Роль продолговатого мозга в регуляции сосудистого тонуса Более детальный анализ показал, что сосудодвигательный центр продолговатого мозга расположен на дне IV желудочка и состоит из двух отделов — прессорного и депрессорного. Раздражение прессорного отдела сосудодвигательного центра вызывает сужение артерий и подъем, а раздражение второго — расширение артерий и падение АД.
Таламус Эпифиз Верхние холмики Средний мозг Нижние холмики Мост Ножки мозжечка Дно 4 -го желудочка Роль продолговатого мозга в регуляции сосудистого тонуса Более детальный анализ показал, что сосудодвигательный центр продолговатого мозга расположен на дне IV желудочка и состоит из двух отделов — прессорного и депрессорного. Раздражение прессорного отдела сосудодвигательного центра вызывает сужение артерий и подъем, а раздражение второго — расширение артерий и падение АД.
 Влияния, идущие от сосудосуживающего центра продолговатого мозга, приходят к нервным центрам симпатической части вегетативной нервной системы, расположенным в боковых рогах грудных сегментов спинного мозга, регулирующих тонус сосудов отдельных участков тела. Спинномозговые центры способны через некоторое время после выключения сосудосуживающего центра продолговатого мозга немного повысить давление крови, снизившееся вследствие расширения артерий и артериол.
Влияния, идущие от сосудосуживающего центра продолговатого мозга, приходят к нервным центрам симпатической части вегетативной нервной системы, расположенным в боковых рогах грудных сегментов спинного мозга, регулирующих тонус сосудов отдельных участков тела. Спинномозговые центры способны через некоторое время после выключения сосудосуживающего центра продолговатого мозга немного повысить давление крови, снизившееся вследствие расширения артерий и артериол.
 Гипоталамический уровень регуляции сосудистого тонуса При возбуждении передних групп ядер гипоталамуса активируется парасимпатическая нервная система - понижение тонуса. Раздражение задних ядер даёт, в основном, сосудосуживающий эффект.
Гипоталамический уровень регуляции сосудистого тонуса При возбуждении передних групп ядер гипоталамуса активируется парасимпатическая нервная система - понижение тонуса. Раздражение задних ядер даёт, в основном, сосудосуживающий эффект.
 – нейронытерморецепторы, постоянно измеряют температуру крови, 80% из") В передней части гипоталамуса (преоптическая область) – нейронытерморецепторы, постоянно измеряют температуру крови, 80% из них реагируют на перегрев, 20% – на охлаждение. Дополнительно (но в меньшей степени) учитываются сигналы от тепловых и холодовых рецепторов кожи. При перегреве – расширение сосудов кожи, потоотделение, поведенческие реакции. При переохлаждении – сужение сосудов кожи, дрожь и пилоэрекция, поведенческие реакции.
В передней части гипоталамуса (преоптическая область) – нейронытерморецепторы, постоянно измеряют температуру крови, 80% из них реагируют на перегрев, 20% – на охлаждение. Дополнительно (но в меньшей степени) учитываются сигналы от тепловых и холодовых рецепторов кожи. При перегреве – расширение сосудов кожи, потоотделение, поведенческие реакции. При переохлаждении – сужение сосудов кожи, дрожь и пилоэрекция, поведенческие реакции.
 Особенности гипоталамической регуляции: осуществляется как компонент терморегуляции; просвет сосудов изменяется в соответствии с изменениями t окружающей среды. Гипоталамический отдел сосудодвигательного центра обеспечивает изменение окраски кожи при эмоциональных реакциях. Гипоталамический отдел сосудодвигательного центра тесно связан с бульбарным и корковым отделами сосудодвигательного центра.
Особенности гипоталамической регуляции: осуществляется как компонент терморегуляции; просвет сосудов изменяется в соответствии с изменениями t окружающей среды. Гипоталамический отдел сосудодвигательного центра обеспечивает изменение окраски кожи при эмоциональных реакциях. Гипоталамический отдел сосудодвигательного центра тесно связан с бульбарным и корковым отделами сосудодвигательного центра.
 Корковый отдел сосудодвигательного центра Раздражение передней центральной извилины, лобной и височной зоны коры головного мозга меняют сосудистый тонус. Кора головного мозга обеспечивает выработку условных рефлексов и на расширение и на сужение сосудов. Условные рефлексы быстрее вырабатываются на сужение, чем на расширение. За счёт коркового отдела сосудодвигательного центра происходит приспособление сосудистой реакции к изменению условий окружающей среды.
Корковый отдел сосудодвигательного центра Раздражение передней центральной извилины, лобной и височной зоны коры головного мозга меняют сосудистый тонус. Кора головного мозга обеспечивает выработку условных рефлексов и на расширение и на сужение сосудов. Условные рефлексы быстрее вырабатываются на сужение, чем на расширение. За счёт коркового отдела сосудодвигательного центра происходит приспособление сосудистой реакции к изменению условий окружающей среды.
 5 6") 1 2 Нервная регуляция сердечных сокращений: 3 4 барорецепторы (растяжение стенок сосудов) 5 6 7 сосуды, мозговое в-во надпочечников хеморецепторы растяжения стенок внутренних органов 1, 2 – сосудодвигательный центр продолговатого мозга и моста и поступающие из него команды; 3 – регулирующие влияния гипоталамуса, больших полушарий и других структур ЦНС, а также рецепторов; 4, 5 – ядра блуждающ. нерва и их парасимпатич. действие; 6, 7 – симпатические эффекты (спинной мозг и ганглии): более обширные проекции. Параллельно развивается влияние симпатической нервной системы на сосуды (сужение) и мозговое вещество надпочечников (выброс адреналина). 32
1 2 Нервная регуляция сердечных сокращений: 3 4 барорецепторы (растяжение стенок сосудов) 5 6 7 сосуды, мозговое в-во надпочечников хеморецепторы растяжения стенок внутренних органов 1, 2 – сосудодвигательный центр продолговатого мозга и моста и поступающие из него команды; 3 – регулирующие влияния гипоталамуса, больших полушарий и других структур ЦНС, а также рецепторов; 4, 5 – ядра блуждающ. нерва и их парасимпатич. действие; 6, 7 – симпатические эффекты (спинной мозг и ганглии): более обширные проекции. Параллельно развивается влияние симпатической нервной системы на сосуды (сужение) и мозговое вещество надпочечников (выброс адреналина). 32
 Барорецепторный рефлекс – компенсаторная реакция на изменение растяжения стенок дуги аорты и каротидного синуса. Еще один рефлекс запускается избыточным растяжением стенок предсердий (если желудочки не успевают откачивать кровь): происходит усиление работы сердца. Если давление оказывается ниже нормы, то активируется симпатическая система, сердце начинает биться чаще и сильнее; если давление выше нормы – активируется блуждающий нерв, работа сердца тормозится.
Барорецепторный рефлекс – компенсаторная реакция на изменение растяжения стенок дуги аорты и каротидного синуса. Еще один рефлекс запускается избыточным растяжением стенок предсердий (если желудочки не успевают откачивать кровь): происходит усиление работы сердца. Если давление оказывается ниже нормы, то активируется симпатическая система, сердце начинает биться чаще и сильнее; если давление выше нормы – активируется блуждающий нерв, работа сердца тормозится.
 5 4 Основные связи сосудодвигательного центра продолговатого мозга и моста (на выходе показаны только симпат. эффекты): 3 1 2 1. Барорецепторы сосудов. 2. Периферические хеморецепторы (хемо. РЦ). 3. Центральные хемо. РЦ. 4. Дыхательные центры. 5. Влияния гипоталамуса (терморегуляция, боль и другие врожденно значимые стимулы, эмоции) и коры больших полушарий (переключаются через гипоталамус и средний мозг; эмоции, связанные с оценкой ситуации как потенциально значимой, опасной и т. п. ; центр таких эмоций – поясная изв. ). 34
5 4 Основные связи сосудодвигательного центра продолговатого мозга и моста (на выходе показаны только симпат. эффекты): 3 1 2 1. Барорецепторы сосудов. 2. Периферические хеморецепторы (хемо. РЦ). 3. Центральные хемо. РЦ. 4. Дыхательные центры. 5. Влияния гипоталамуса (терморегуляция, боль и другие врожденно значимые стимулы, эмоции) и коры больших полушарий (переключаются через гипоталамус и средний мозг; эмоции, связанные с оценкой ситуации как потенциально значимой, опасной и т. п. ; центр таких эмоций – поясная изв. ). 34
") Дыхательная аритмия: результат влияния дыхательного центра на сосудодвигательный (на примере частоты сердечных сокращений (ЧСС) собаки): во время вдоха интервал между сокращениями сердца уменьшается (ЧСС растет); во время выдоха – наоборот. вдох – рост ЧСС выдох – снижение ЧСС Интервал между сердечн. сокращениями, мс сек Выдох Вдох
Дыхательная аритмия: результат влияния дыхательного центра на сосудодвигательный (на примере частоты сердечных сокращений (ЧСС) собаки): во время вдоха интервал между сокращениями сердца уменьшается (ЧСС растет); во время выдоха – наоборот. вдох – рост ЧСС выдох – снижение ЧСС Интервал между сердечн. сокращениями, мс сек Выдох Вдох
 регулирует тонус сосудов; 2) распределяет кровь в организме; 3) регулирует") Значение сосудодвигательного центра: 1) регулирует тонус сосудов; 2) распределяет кровь в организме; 3) регулирует кровяное давление; 4) участвует в терморегуляции; 5) обеспечивает эмоциональные реакции.
Значение сосудодвигательного центра: 1) регулирует тонус сосудов; 2) распределяет кровь в организме; 3) регулирует кровяное давление; 4) участвует в терморегуляции; 5) обеспечивает эмоциональные реакции.
 Сосательный рефлекс – возникает при") Центры, обеспечивающие врожденное пищевое поведение (сосание, жевание, слюноотделение, глотание) Сосательный рефлекс – возникает при раздражении чувствительных окончаний тройничного нерва, возбуждение которого переключается в продолговатом мозге на моторные ядра лицевого и подъязычного нервов.
Центры, обеспечивающие врожденное пищевое поведение (сосание, жевание, слюноотделение, глотание) Сосательный рефлекс – возникает при раздражении чувствительных окончаний тройничного нерва, возбуждение которого переключается в продолговатом мозге на моторные ядра лицевого и подъязычного нервов.
 Совокупность управляющих жеванием нейронов различных отделов мозга называют центром жевания. Акт жевания осуществляется рефлекторно, имеет цепной характер, автоматизированные и произвольные компоненты. Импульсы от рецепторов полости рта по волокнам тройничного нерва передаются в сенсорные ядра продолговатого мозга, ядра зрительного бугра, оттуда — в кору большого мозга. От ствола мозга и зрительного бугра коллатерали отходят к ретикулярной формации. В регуляции жевания принимают участие двигательные ядра продолговатого мозга, красное ядро, черное вещество, подкорковые ядра и кора большого мозга.
Совокупность управляющих жеванием нейронов различных отделов мозга называют центром жевания. Акт жевания осуществляется рефлекторно, имеет цепной характер, автоматизированные и произвольные компоненты. Импульсы от рецепторов полости рта по волокнам тройничного нерва передаются в сенсорные ядра продолговатого мозга, ядра зрительного бугра, оттуда — в кору большого мозга. От ствола мозга и зрительного бугра коллатерали отходят к ретикулярной формации. В регуляции жевания принимают участие двигательные ядра продолговатого мозга, красное ядро, черное вещество, подкорковые ядра и кора большого мозга.
 Импульсы от продолговатого мозга по двигательным волокнам тройничного нерва поступают к жевательным мышцам. Они осуществляют движения нижней челюсти, языка, щек и губ. В координации жевания большую роль играют импульсы от рецепторов жевательных мышц и зубов.
Импульсы от продолговатого мозга по двигательным волокнам тройничного нерва поступают к жевательным мышцам. Они осуществляют движения нижней челюсти, языка, щек и губ. В координации жевания большую роль играют импульсы от рецепторов жевательных мышц и зубов.
 Слюноотделение Слюна продуцируется тремя парами крупных слюнных желез и множеством мелких железок языка, слизистой оболочки неба и щек. Из желез по выводным протокам слюна поступает в полость рта. Разные железы выделяют слюну разного состава. Возбуждение от рецепторов полости рта передается в ЦНС по афферентным волокнам тройничного (5), лицевого (7), языкоглоточного (9) и блуждающего (10) нервов.
Слюноотделение Слюна продуцируется тремя парами крупных слюнных желез и множеством мелких железок языка, слизистой оболочки неба и щек. Из желез по выводным протокам слюна поступает в полость рта. Разные железы выделяют слюну разного состава. Возбуждение от рецепторов полости рта передается в ЦНС по афферентным волокнам тройничного (5), лицевого (7), языкоглоточного (9) и блуждающего (10) нервов.
 Импульсы достигают продолговатого и других отделов мозга, включая кору мозга. Основной центр слюноотделения расположен в продолговатом мозге, сюда и в боковые рога верхних грудных сегментов спинного мозга поступают импульсы из вышерасположенных отделов мозга. К слюнным железам импульсы следуют по эфферентным парасимпатическим и симпатическим нервным волокнам.
Импульсы достигают продолговатого и других отделов мозга, включая кору мозга. Основной центр слюноотделения расположен в продолговатом мозге, сюда и в боковые рога верхних грудных сегментов спинного мозга поступают импульсы из вышерасположенных отделов мозга. К слюнным железам импульсы следуют по эфферентным парасимпатическим и симпатическим нервным волокнам.
 Глотание Жевание завершается глотанием — переходом пищевого комка из полости рта в желудок. Глотание возникает в результате раздражения чувствительных нервных окончаний тройничного (5) и языкоглоточного (9) нервов. По афферентным волокнам этих нервов импульсы поступают в продолговатый мозг, где расположен центр глотания. От него импульсы по эфферентным двигательным волокнам тройничного (5), языкоглоточного (9), подъязычного (12) и блуждающего (10) нервов достигают мышц, обеспечивающих глотание.
Глотание Жевание завершается глотанием — переходом пищевого комка из полости рта в желудок. Глотание возникает в результате раздражения чувствительных нервных окончаний тройничного (5) и языкоглоточного (9) нервов. По афферентным волокнам этих нервов импульсы поступают в продолговатый мозг, где расположен центр глотания. От него импульсы по эфферентным двигательным волокнам тройничного (5), языкоглоточного (9), подъязычного (12) и блуждающего (10) нервов достигают мышц, обеспечивающих глотание.
 Деятельность центра глотания в продолговатом мозге координируется двигательными центрами среднего мозга, коры больших полушарий. Бульбарный центр находится в тесной связи с центром дыхания, тормозя его при глотании, что предотвращает попадание пищи в воздухоносные пути.
Деятельность центра глотания в продолговатом мозге координируется двигательными центрами среднего мозга, коры больших полушарий. Бульбарный центр находится в тесной связи с центром дыхания, тормозя его при глотании, что предотвращает попадание пищи в воздухоносные пути.
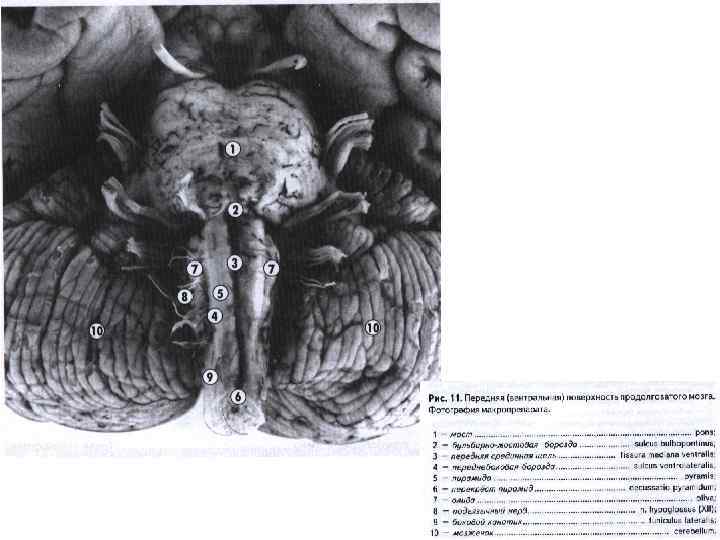
: III — глазодвигательный нерв; IV — блоковый нерв; V") Ствол мозга с вентральной стороны): III — глазодвигательный нерв; IV — блоковый нерв; V — тройничный нерв; VI — отводящий нерв; VII — лицевой нерв; VIII — предверно-улитковый нерв (слуховой); IX — языкоглоточный нерв; X — блуждающий нерв; XI — добавочный нерв; XII — подъязычный нерв; 1 — ножка мозга (средний мозг); 2 — перекрест пирамид (верхний шейный отдел спинного мозга); 3 — боковой канатик; 4 — передняя боковая борозда; 5 — олива; 6 — пирамида продолговатого мозга; 7 — мост
Ствол мозга с вентральной стороны): III — глазодвигательный нерв; IV — блоковый нерв; V — тройничный нерв; VI — отводящий нерв; VII — лицевой нерв; VIII — предверно-улитковый нерв (слуховой); IX — языкоглоточный нерв; X — блуждающий нерв; XI — добавочный нерв; XII — подъязычный нерв; 1 — ножка мозга (средний мозг); 2 — перекрест пирамид (верхний шейный отдел спинного мозга); 3 — боковой канатик; 4 — передняя боковая борозда; 5 — олива; 6 — пирамида продолговатого мозга; 7 — мост
 Продолговатый мозг Черепные нервы Вкусовые центры продолговатого мозга и моста (сигналы от языка VII и IX н. ; от глотки – X н. ): в зависимости от «хорошего» и «плохого» вкуса запускаются пищевые либо оборонительные реакции. горький кислый соленый сладкий «Хороший» вкус (рецепторы глюкозы; биологически полезные вещества): сосание, жевание, глотание, выделение желудочного сока и «густой» слюны с пищеварительными ферментами (парасимпатическая реакция). «Плохой» вкус (рецепторы горького – растительные токсины; избыток кислого и соленого): выплевывание, плач, рвота, выделение большого количества жидкой слюны (смыть «эту гадость» ; симпатическая реакция). 45
Продолговатый мозг Черепные нервы Вкусовые центры продолговатого мозга и моста (сигналы от языка VII и IX н. ; от глотки – X н. ): в зависимости от «хорошего» и «плохого» вкуса запускаются пищевые либо оборонительные реакции. горький кислый соленый сладкий «Хороший» вкус (рецепторы глюкозы; биологически полезные вещества): сосание, жевание, глотание, выделение желудочного сока и «густой» слюны с пищеварительными ферментами (парасимпатическая реакция). «Плохой» вкус (рецепторы горького – растительные токсины; избыток кислого и соленого): выплевывание, плач, рвота, выделение большого количества жидкой слюны (смыть «эту гадость» ; симпатическая реакция). 45
 Слуховые и вестибулярные ядра Вестибулярная информация необходима для коррекции движений; в связи с этим она расходится по 4 -м направлениям: через таламус в кору (управление произвольными движениями); в мозжечок (управление автоматизированными движениями); в глазодвигательные центры среднего мозга; в спинной мозг (вестибулоспинальные тракты).
Слуховые и вестибулярные ядра Вестибулярная информация необходима для коррекции движений; в связи с этим она расходится по 4 -м направлениям: через таламус в кору (управление произвольными движениями); в мозжечок (управление автоматизированными движениями); в глазодвигательные центры среднего мозга; в спинной мозг (вестибулоспинальные тракты).
 Вестиб. мешочки и каналы Улитка Вестибулярные ядра: информация от мешочков и полукружных каналов Слуховые ядра: от улитки Вестибулярные и слуховые ядра – по углам ромбовидной ямки; вестибулярные ядра медиальнее, как эволюционно более древние
Вестиб. мешочки и каналы Улитка Вестибулярные ядра: информация от мешочков и полукружных каналов Слуховые ядра: от улитки Вестибулярные и слуховые ядра – по углам ромбовидной ямки; вестибулярные ядра медиальнее, как эволюционно более древние
 и таламус; у человека") Далее слуховая информация идет в средний мозг (нижние холмики ч/х) и таламус; у человека – сравнение сигналов от правого и левого уха, определения направления на источник звука.
Далее слуховая информация идет в средний мозг (нижние холмики ч/х) и таламус; у человека – сравнение сигналов от правого и левого уха, определения направления на источник звука.
 В последнем случае возможен запуск ряда врожденных рефлексов: ровная установка головы ( «ребенок начинает держать голову» , органы зрения и слуха приводятся в оптимальное положение).
В последнем случае возможен запуск ряда врожденных рефлексов: ровная установка головы ( «ребенок начинает держать голову» , органы зрения и слуха приводятся в оптимальное положение).
 Центры сна и бодрствования, стадии сна Центры сна и бодрствования эволюционно очень древние, постоянно конкурируют друг с другом, учитывают значительное число факторов (прежде всего, сенсорных). 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг Мозжечок
Центры сна и бодрствования, стадии сна Центры сна и бодрствования эволюционно очень древние, постоянно конкурируют друг с другом, учитывают значительное число факторов (прежде всего, сенсорных). 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг Мозжечок
 1 - Главный центр бодрствования: ретикулярные ядра моста; сюда поступает часть информации от всех сенсорных систем; далее происходит оценка общего уровня «сенсорного давления» на ЦНС, и чем оно больше, тем мозг активнее (нас будит сигнал, поступивший от любой сенсорной системы); аксоны расходятся по всей ЦНС, задавая ее тонус ( «блок питания» мозга); при торможении сенсорных потоков мы засыпаем. Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг Мозжечок
1 - Главный центр бодрствования: ретикулярные ядра моста; сюда поступает часть информации от всех сенсорных систем; далее происходит оценка общего уровня «сенсорного давления» на ЦНС, и чем оно больше, тем мозг активнее (нас будит сигнал, поступивший от любой сенсорной системы); аксоны расходятся по всей ЦНС, задавая ее тонус ( «блок питания» мозга); при торможении сенсорных потоков мы засыпаем. Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг Мозжечок
 2 - Главный центр сна: центральное серое вещество среднего мозга и ядра шва; аксоны нейронов ядер шва также расходятся по всей ЦНС, снижая ее тонус и тормозя, в числе прочего, центры бодрствования. Торможение коры происходит за счет снижения активности нейронов таламуса, чьи аксоны идут в большие полушария 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг Мозжечок
2 - Главный центр сна: центральное серое вещество среднего мозга и ядра шва; аксоны нейронов ядер шва также расходятся по всей ЦНС, снижая ее тонус и тормозя, в числе прочего, центры бодрствования. Торможение коры происходит за счет снижения активности нейронов таламуса, чьи аксоны идут в большие полушария 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг Мозжечок
![3 - Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2]. При](https://present5.com/presentation/26067410_63064117/image-53.jpg "3 - Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2]. При") 3 - Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2]. При стрессе, приближении потенциально опасной ситуации трудно заснуть 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг 3 Мозжечок
3 - Голубое пятно: вспомогательный центр бодрствования, получив сигнал из [1], тормозит [2]. При стрессе, приближении потенциально опасной ситуации трудно заснуть 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг 3 Мозжечок
 4 - Супрахиазменные ядра переднего гипоталамуса: находятся напротив перекреста зрительных нервов, получают информацию об общем уровне освещенности и настраиваются на суточный ритм ( «биологические часы» ; часть нейронов активны днем и влияют на [1], часть – ночью и влияют на [2]. В яркой форме эффект «биологических часов» проявляется при резкой смене часового пояса. 4 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг 3 5 Мозжечок
4 - Супрахиазменные ядра переднего гипоталамуса: находятся напротив перекреста зрительных нервов, получают информацию об общем уровне освещенности и настраиваются на суточный ритм ( «биологические часы» ; часть нейронов активны днем и влияют на [1], часть – ночью и влияют на [2]. В яркой форме эффект «биологических часов» проявляется при резкой смене часового пояса. 4 2 Гипоталамус 1 Ножки мозга Четверохолмие Мост Продолговатый мозг 3 5 Мозжечок
 5 - Вспомогательный центр сна – ретикулярные ядра продолговатого мозга: реакция на химический состав крови, появление «отходов обмена веществ» , токсинов (при заболеваниях и отравлениях), рост концентрации инсулина и глюкозы (после еды хочется спать); оказывает постоянное возбуждающее действие на [2]. 4 2 Гипоталамус 1 Ножки мозга 3 Четверохолмие Мост Продолговатый мозг Мозжечок 5
5 - Вспомогательный центр сна – ретикулярные ядра продолговатого мозга: реакция на химический состав крови, появление «отходов обмена веществ» , токсинов (при заболеваниях и отравлениях), рост концентрации инсулина и глюкозы (после еды хочется спать); оказывает постоянное возбуждающее действие на [2]. 4 2 Гипоталамус 1 Ножки мозга 3 Четверохолмие Мост Продолговатый мозг Мозжечок 5
 вся ЦНС СЕНСОРНЫЕ СИСТЕМЫ Ретикулярные ядра моста СТРЕСС Голубое пятно Центральное серое вещество и ядра шва Супрахиаз 4. Супрахиазменные ядра переднего гипоталамуса: находятся напротив менные перекреста зрительных нервов, получают информацию об общем уровне ядра освещенности и настраиваются на суточный ритм ( «биологические часы» ; часть нейронов активны днем и влияют на [1], часть – ночью и влияют на Ретикулярные в яркой Ц Н С эффект «биологич. часов» с я форме [2], намекая, что пора спать). В ядра продолпроявляется при резкой смене часового пояса. В основе поддержания говатого мозга общий суточного ритма – медленные цепи внутриклеточных химических реакций. В норме центры сна и бодрствоуровень вания постоянно конкурируют, 5. Вспомогательный центр сна – ретикулярные ядра продолговатого освещенмозга: реакция на часто возникают «промежуточхимический состав крови, появление аденозина и других ности «отходов обмена ные» состояния, мы (при заболеваниях и Химический веществ» , токсинов засыпаем и отравлениях), просыпаемся постепенно. рост концентрации инсулина и глюкозы (после еды хочется спать); 56 состав крови оказывает постоянное возбуждающее действие на [2].
вся ЦНС СЕНСОРНЫЕ СИСТЕМЫ Ретикулярные ядра моста СТРЕСС Голубое пятно Центральное серое вещество и ядра шва Супрахиаз 4. Супрахиазменные ядра переднего гипоталамуса: находятся напротив менные перекреста зрительных нервов, получают информацию об общем уровне ядра освещенности и настраиваются на суточный ритм ( «биологические часы» ; часть нейронов активны днем и влияют на [1], часть – ночью и влияют на Ретикулярные в яркой Ц Н С эффект «биологич. часов» с я форме [2], намекая, что пора спать). В ядра продолпроявляется при резкой смене часового пояса. В основе поддержания говатого мозга общий суточного ритма – медленные цепи внутриклеточных химических реакций. В норме центры сна и бодрствоуровень вания постоянно конкурируют, 5. Вспомогательный центр сна – ретикулярные ядра продолговатого освещенмозга: реакция на часто возникают «промежуточхимический состав крови, появление аденозина и других ности «отходов обмена ные» состояния, мы (при заболеваниях и Химический веществ» , токсинов засыпаем и отравлениях), просыпаемся постепенно. рост концентрации инсулина и глюкозы (после еды хочется спать); 56 состав крови оказывает постоянное возбуждающее действие на [2].
 REM: rapid eye movements На основе анализа ЭЭГвыделяют стадии сна: Бодрствование: альфа-ритм – 10 -12 Гц, бета-ритм – 15 -30 Гц. Стадия 1: появление тета-ритма – 48 Гц. Стадия 2: сонные веретена и К-комплексы. Стадии 3 и 4: все более медленный дельта-ритм – 1 -3 Гц. REM-сон: «бодрствующая» ЭЭГ.
REM: rapid eye movements На основе анализа ЭЭГвыделяют стадии сна: Бодрствование: альфа-ритм – 10 -12 Гц, бета-ритм – 15 -30 Гц. Стадия 1: появление тета-ритма – 48 Гц. Стадия 2: сонные веретена и К-комплексы. Стадии 3 и 4: все более медленный дельта-ритм – 1 -3 Гц. REM-сон: «бодрствующая» ЭЭГ.
 СНЫ – «окно в бессознательное» , «упорядочивание» накопленной информации, продолжение ментальных процессов в ином состоянии (творческие сны, вещие сны и т. п. ). Если лишать REM-сна, то человек не высыпается, а на следующую ночь «добирает» REM-сон. Развитый REM-сон – только у млекопитающих. «Сон навеянный полётом пчелы вокруг граната за миг до пробуждения» , 1943
СНЫ – «окно в бессознательное» , «упорядочивание» накопленной информации, продолжение ментальных процессов в ином состоянии (творческие сны, вещие сны и т. п. ). Если лишать REM-сна, то человек не высыпается, а на следующую ночь «добирает» REM-сон. Развитый REM-сон – только у млекопитающих. «Сон навеянный полётом пчелы вокруг граната за миг до пробуждения» , 1943
 – физиологический отдых мозга разной степени глубины. REM-сон (парадоксальный: «бодрствующая»") Стадии 1 -4 (не-REM-сон) – физиологический отдых мозга разной степени глубины. REM-сон (парадоксальный: «бодрствующая» ЭЭГ, но порог пробуждения выше) – стадия сновидений, обработка накопленной информации (в первую очередь, за текущие сутки). Около 20% времени сна; 4 -5 раз за ночь примерно по 20 мин; в первые 3 года жизни – 3050%. «Сон навеянный полётом пчелы вокруг граната за миг до пробуждения» , 1943
Стадии 1 -4 (не-REM-сон) – физиологический отдых мозга разной степени глубины. REM-сон (парадоксальный: «бодрствующая» ЭЭГ, но порог пробуждения выше) – стадия сновидений, обработка накопленной информации (в первую очередь, за текущие сутки). Около 20% времени сна; 4 -5 раз за ночь примерно по 20 мин; в первые 3 года жизни – 3050%. «Сон навеянный полётом пчелы вокруг граната за миг до пробуждения» , 1943
 Средний мозг и ориентировочный рефлекс Центральное серое вещество: собирает большое число информационных потоков и влияет на уровень бодрствования, болевой чувствительности и др. Четверохолмие: реакция на новизну; верхние холмики – на новые зрительные стимулы; нижние холмики – на новые слуховые стимулы. При появлении новых стимулов четверохолмие запускает ориентировочный рефлекс – поворот глаз, головы и всего тела в сторону источника сигнала ( «любопытство» , исследовательское поведение). Верхние холмики Мозговой водопровод Центральное серое вещество Черная субстанция Красное ядро Ножки мозга
Средний мозг и ориентировочный рефлекс Центральное серое вещество: собирает большое число информационных потоков и влияет на уровень бодрствования, болевой чувствительности и др. Четверохолмие: реакция на новизну; верхние холмики – на новые зрительные стимулы; нижние холмики – на новые слуховые стимулы. При появлении новых стимулов четверохолмие запускает ориентировочный рефлекс – поворот глаз, головы и всего тела в сторону источника сигнала ( «любопытство» , исследовательское поведение). Верхние холмики Мозговой водопровод Центральное серое вещество Черная субстанция Красное ядро Ножки мозга
 Зрительная информация: волокна II н. Таламус Слуховая Средний мозг информация: Мост ядра VIII н. Эпифиз Верхние холмики Нижние холмики Кожная чувстви. Ножки мозжечка тельность: Дно 4 -го желудочка спинной мозг и ядра V н.
Зрительная информация: волокна II н. Таламус Слуховая Средний мозг информация: Мост ядра VIII н. Эпифиз Верхние холмики Нижние холмики Кожная чувстви. Ножки мозжечка тельность: Дно 4 -го желудочка спинной мозг и ядра V н.
 – сравнение текущего сигнала с тем, который был «только что»") Четверохолмие: нейроны-детекторы новизны (ДН) – сравнение текущего сигнала с тем, который был «только что» (доли секунды назад, передается через тормозный интернейрон: ТИ). При несовпадении – запуск ориентировочного рефлекса (через глазодвигательные центры и тектоспинальный тракт; у животных – отдельно двигаются ушные раковины). ДН ТИ Ориентировочн. рефлекс (если возб-е > торм-я)
Четверохолмие: нейроны-детекторы новизны (ДН) – сравнение текущего сигнала с тем, который был «только что» (доли секунды назад, передается через тормозный интернейрон: ТИ). При несовпадении – запуск ориентировочного рефлекса (через глазодвигательные центры и тектоспинальный тракт; у животных – отдельно двигаются ушные раковины). ДН ТИ Ориентировочн. рефлекс (если возб-е > торм-я)
 Движения глаз: с каждым глазом связано по 6 мышц, управляемых III, IV и IV нервами; два основных типа движений глаз – слежения и саккады (быстрые скачки); в основе - врожденные программы, но мы учимся ими управлять.
Движения глаз: с каждым глазом связано по 6 мышц, управляемых III, IV и IV нервами; два основных типа движений глаз – слежения и саккады (быстрые скачки); в основе - врожденные программы, но мы учимся ими управлять.
![Чтение: [1] – скачок в начало строки; [2] мини-саккады (5 -7 скачков вдоль строки,](https://present5.com/presentation/26067410_63064117/image-64.jpg "Чтение: [1] – скачок в начало строки; [2] мини-саккады (5 -7 скачков вдоль строки,") Чтение: [1] – скачок в начало строки; [2] мини-саккады (5 -7 скачков вдоль строки, текст читается «в несколько приемов» ). 1 2
Чтение: [1] – скачок в начало строки; [2] мини-саккады (5 -7 скачков вдоль строки, текст читается «в несколько приемов» ). 1 2
 И. Е. Репин «Не ждали»
И. Е. Репин «Не ждали»
 Чёрная субстанция. Медиальная «компактная» часть , DA-нейроны, аксоны идут в базальные ганглии (полосатое тело = скорлупа, хвостатое ядро); общий уровень двигательной активности и положительные эмоции, связанные с движениями. Латеральная «ретикулярная» часть , ГАМК-нейроны, контролирующие движения глаз Верхние холмики Мозговой водопровод Центральное серое вещество Черная субстанция Красное ядро Ножки мозга
Чёрная субстанция. Медиальная «компактная» часть , DA-нейроны, аксоны идут в базальные ганглии (полосатое тело = скорлупа, хвостатое ядро); общий уровень двигательной активности и положительные эмоции, связанные с движениями. Латеральная «ретикулярная» часть , ГАМК-нейроны, контролирующие движения глаз Верхние холмики Мозговой водопровод Центральное серое вещество Черная субстанция Красное ядро Ножки мозга
 Чёрная субстанция. Медиальная «компактная» часть , DA-нейроны, аксоны идут в базальные ганглии (полосатое тело = скорлупа, хвостатое ядро); общий уровень двигат. активности и положит. эмоции, связанные с движениями. Латеральная «ретикулярная» часть , ГАМК-нейроны, контролирующие движения глаз (торможение «несанкционированных» реакций). ГАМКнейроны черной субстанции Глазодвигательные центры (ядра III, IV, VI н. ) Движения глаз ГАМКнейроны полосатого тела Сигнал о несовпадении (четверохолмие) Кора больших полушарий (произвольные движения): для запуска нужно активи-ровать нейроны полосатого тела, которые затормозят клетки черной субстанции ( «торможение торможения» ) Верхние холмики Мозговой водопровод Центральное серое вещество Черная субстанция Красное ядро 67 Ножки мозга
Чёрная субстанция. Медиальная «компактная» часть , DA-нейроны, аксоны идут в базальные ганглии (полосатое тело = скорлупа, хвостатое ядро); общий уровень двигат. активности и положит. эмоции, связанные с движениями. Латеральная «ретикулярная» часть , ГАМК-нейроны, контролирующие движения глаз (торможение «несанкционированных» реакций). ГАМКнейроны черной субстанции Глазодвигательные центры (ядра III, IV, VI н. ) Движения глаз ГАМКнейроны полосатого тела Сигнал о несовпадении (четверохолмие) Кора больших полушарий (произвольные движения): для запуска нужно активи-ровать нейроны полосатого тела, которые затормозят клетки черной субстанции ( «торможение торможения» ) Верхние холмики Мозговой водопровод Центральное серое вещество Черная субстанция Красное ядро 67 Ножки мозга
 Красное ядро Передняя мелкоклеточная часть: вместе с нижней оливой передает сигналы от коры больших полушарий к мозжечку и участвует в двигательном обучении. Задняя (крупноклеточная) часть эволюционно более древняя, содержит нейроны, аксоны которых идут в спинной мозг (руброспинальный тракт; поддержание тонуса мышц, ряд сгибательных рефлексов и сгибание конечностей при локомоции).
Красное ядро Передняя мелкоклеточная часть: вместе с нижней оливой передает сигналы от коры больших полушарий к мозжечку и участвует в двигательном обучении. Задняя (крупноклеточная) часть эволюционно более древняя, содержит нейроны, аксоны которых идут в спинной мозг (руброспинальный тракт; поддержание тонуса мышц, ряд сгибательных рефлексов и сгибание конечностей при локомоции).
 тракта; еще не способен обеспечить тонкое управление моторикой") Руброспинальный тракт – предшественник кортико-спинального (пирамидного) тракта; еще не способен обеспечить тонкое управление моторикой пальцев, может лишь вызвать совместное (синергичное) их сгибание Руброспиный тракт – часть так называемой экстрапирамидной системы управления движениями, в которую входят также вестибулоспинальный и ретикуло-спинальный тракты.
Руброспинальный тракт – предшественник кортико-спинального (пирамидного) тракта; еще не способен обеспечить тонкое управление моторикой пальцев, может лишь вызвать совместное (синергичное) их сгибание Руброспиный тракт – часть так называемой экстрапирамидной системы управления движениями, в которую входят также вестибулоспинальный и ретикуло-спинальный тракты.
 Руброспинальный тракт – сгибание конечностей (в том числе при локомоции – то есть ходьбе, беге и т. п. ). Вестибулоспинальный тракт – разгибание конечностей (рефлексы, локомоция). Ретикулоспинальные тракты – идут от РФ, сгибание и разгибание туловища (самые древние двигательные тракты, с их помощью плавают рыбы). Кроме этого, на схеме представлены кортикоспинальный (пирамидный) тракт и тектоспинальные тракты. 70
Руброспинальный тракт – сгибание конечностей (в том числе при локомоции – то есть ходьбе, беге и т. п. ). Вестибулоспинальный тракт – разгибание конечностей (рефлексы, локомоция). Ретикулоспинальные тракты – идут от РФ, сгибание и разгибание туловища (самые древние двигательные тракты, с их помощью плавают рыбы). Кроме этого, на схеме представлены кортикоспинальный (пирамидный) тракт и тектоспинальные тракты. 70