Лек 10. 2014.ppt
- Количество слайдов: 45
 Лекция 10 Ядро Структурная организация наследственного материала Клеточный цикл Вольты Протеасома
Лекция 10 Ядро Структурная организация наследственного материала Клеточный цикл Вольты Протеасома
 Прокариотические клетки бактерий Ядра нет Геном замкнутый, циркулярный, организован в виде одной хромосомы
Прокариотические клетки бактерий Ядра нет Геном замкнутый, циркулярный, организован в виде одной хромосомы
 Эукариотическая клетка, интерфаза Есть ядро Геном линейный сегментированный и упакован в большое количество хромосом
Эукариотическая клетка, интерфаза Есть ядро Геном линейный сегментированный и упакован в большое количество хромосом
 цитоплазма ЭПР Ядерные поры Ядро внутренняя мембрана хроматин Ядерная пластинка наружная мембрана цитоплазма
цитоплазма ЭПР Ядерные поры Ядро внутренняя мембрана хроматин Ядерная пластинка наружная мембрана цитоплазма
 Интерфазное ядро
Интерфазное ядро
 Ядерная оболочка: внутренняя и внешняя мембраны, различаются по составу белков. Наружная мембрана – часть ЭПР, несет рибосомы. Внутренняя мембрана – производное ЭПР, взаимодействует с белками ядерной пластинки. Перинуклеарное пространство
Ядерная оболочка: внутренняя и внешняя мембраны, различаются по составу белков. Наружная мембрана – часть ЭПР, несет рибосомы. Внутренняя мембрана – производное ЭПР, взаимодействует с белками ядерной пластинки. Перинуклеарное пространство
 ламины транс-мембранные белки связь с цитоскелетом Белки, формирующие пластинку, - ламины,") Ядерная пластинка (ламина) ламины транс-мембранные белки связь с цитоскелетом Белки, формирующие пластинку, - ламины, - относятся к классу промежуточных филаментов. Пластинка служит местом прикрепления хроматина.
Ядерная пластинка (ламина) ламины транс-мембранные белки связь с цитоскелетом Белки, формирующие пластинку, - ламины, - относятся к классу промежуточных филаментов. Пластинка служит местом прикрепления хроматина.
 Ядерная пластинка ооцита морского ежа. Реплика, ядро было обработано тритоном Х-100
Ядерная пластинка ооцита морского ежа. Реплика, ядро было обработано тритоном Х-100
 Диаметр около 120 нм Негативное контрастирование Сканирующая электронная микроскопия Реплика,") Ядерные поры (поровые комплексы) Диаметр около 120 нм Негативное контрастирование Сканирующая электронная микроскопия Реплика, замораживание-травление
Ядерные поры (поровые комплексы) Диаметр около 120 нм Негативное контрастирование Сканирующая электронная микроскопия Реплика, замораживание-травление
 50 нм
50 нм
 нуклеопорины экспериментально доказанные связи гипотетические связи
нуклеопорины экспериментально доказанные связи гипотетические связи
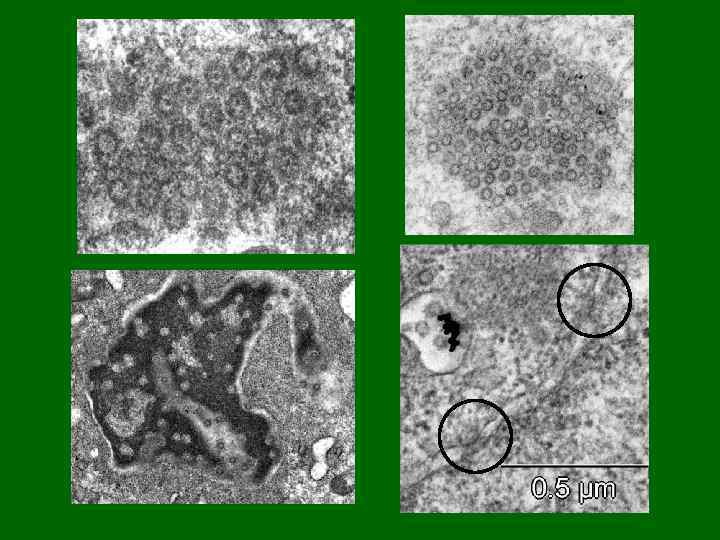
 Ядерные поры Ультратонкие срезы
Ядерные поры Ультратонкие срезы
 (Annulated lamellae)") Кольчатые пластинки (цитоплазматические поры) (Annulated lamellae)
Кольчатые пластинки (цитоплазматические поры) (Annulated lamellae)
 Ядрышко ГЗ ЯО Фибриллярная зона = ядрышковый организатор ПФЗ
Ядрышко ГЗ ЯО Фибриллярная зона = ядрышковый организатор ПФЗ
 Ядрышковый организатор – участки хромосом, где транскрибируются рибосомальные РНК, здесь локализованы гены р. РНК (около 400 генов у человека) ГЗ ПФЗ ЯО Фибриллярная зона – место транскрипции генов р. РНК Плотная ФЗ: процессинг и начало сборки рибосом Гранулярная зона - место сборки и накопления рибосом. Малая и большая субъединицы транспортируются в цитоплазму по отдельности, через поровые комплексы.
Ядрышковый организатор – участки хромосом, где транскрибируются рибосомальные РНК, здесь локализованы гены р. РНК (около 400 генов у человека) ГЗ ПФЗ ЯО Фибриллярная зона – место транскрипции генов р. РНК Плотная ФЗ: процессинг и начало сборки рибосом Гранулярная зона - место сборки и накопления рибосом. Малая и большая субъединицы транспортируются в цитоплазму по отдельности, через поровые комплексы.
 Структурная организация наследственного материала в клетках эукариот Деспирализованный хроматин Хроматин – комплекс ДНК и белков Длина ДНК человека около 2 м, упакована в ядро диаметром 5 -10 мкм Деспирализованный хроматин 4 -нм нити ДНК. Получаются при удалении белков из ядра
Структурная организация наследственного материала в клетках эукариот Деспирализованный хроматин Хроматин – комплекс ДНК и белков Длина ДНК человека около 2 м, упакована в ядро диаметром 5 -10 мкм Деспирализованный хроматин 4 -нм нити ДНК. Получаются при удалении белков из ядра
 нуклеосома 200 пар оснований Хроматосома- 166 П. О. гистоны
нуклеосома 200 пар оснований Хроматосома- 166 П. О. гистоны
 2, 7 нм между стрелками 10 -нм нить – транскрипция 30 -нм нить Основная форма эухроматина в интерфазе 10 -нм нить – транскрипция
2, 7 нм между стрелками 10 -нм нить – транскрипция 30 -нм нить Основная форма эухроматина в интерфазе 10 -нм нить – транскрипция
 Максимальная спирализация хроматина – в период митоза и мейоза Центромера
Максимальная спирализация хроматина – в период митоза и мейоза Центромера
 Митотические хромосомы в электронном микроскопе Нейтрофил Лимфоцит Клетка гепатомы А-1
Митотические хромосомы в электронном микроскопе Нейтрофил Лимфоцит Клетка гепатомы А-1
 В интерфазе: Около 10% эухроматина находится в состоянии 10 -нм нитей – активная транскрипция Около 10% хроматина высококонденсировано – гетерохроматин Гетерохроматин: – конститутивный, или структурный никогда не транскрибируется (центромеры); - факультативный – не транскрибируется в момент исследования Главные принципы пространственной организации хромосом в интерфазном ядре: - полярное расположение центромеров и теломеров - контакты с ядерной оболочкой в определенных участках
В интерфазе: Около 10% эухроматина находится в состоянии 10 -нм нитей – активная транскрипция Около 10% хроматина высококонденсировано – гетерохроматин Гетерохроматин: – конститутивный, или структурный никогда не транскрибируется (центромеры); - факультативный – не транскрибируется в момент исследования Главные принципы пространственной организации хромосом в интерфазном ядре: - полярное расположение центромеров и теломеров - контакты с ядерной оболочкой в определенных участках
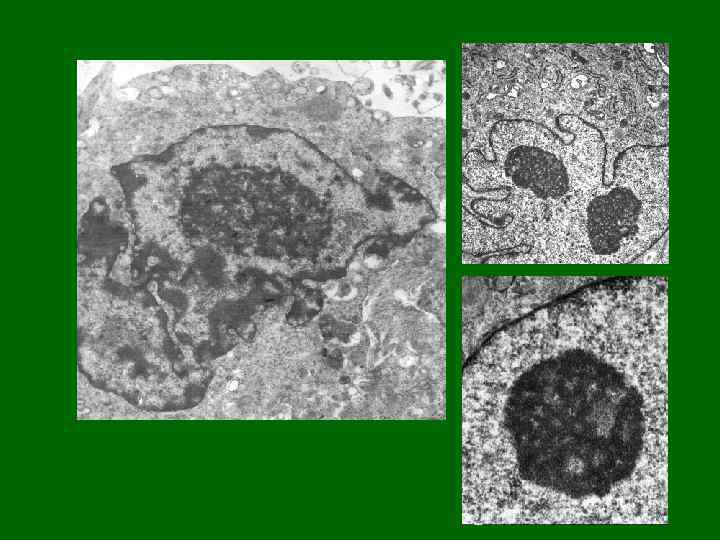
 Ядро с высоким содержанием эухроматина Ультратонкий срез Ядро с высоким содержанием гетерохроматина Ультратонкий срез
Ядро с высоким содержанием эухроматина Ультратонкий срез Ядро с высоким содержанием гетерохроматина Ультратонкий срез
 Ядра с различным содержанием гетерохроматина Клетки лимфоидной ткани
Ядра с различным содержанием гетерохроматина Клетки лимфоидной ткани
 Клеточный цикл 4 стадии: рост клеток, дупликация ДНК, разделение хромосом по дочерним клеткам, деление клетки М – фаза митоза G 1 -период – подготовка к дупликации ДНК, высокая метаболическая активность, рост клетки S-период – фаза синтеза - репликация ДНК интерфаза G 2 -период – рост клетки и подготовка к митозу М – фаза митоза Продолжительность фаз клеточного цикла разных клеток сильно варьирует
Клеточный цикл 4 стадии: рост клеток, дупликация ДНК, разделение хромосом по дочерним клеткам, деление клетки М – фаза митоза G 1 -период – подготовка к дупликации ДНК, высокая метаболическая активность, рост клетки S-период – фаза синтеза - репликация ДНК интерфаза G 2 -период – рост клетки и подготовка к митозу М – фаза митоза Продолжительность фаз клеточного цикла разных клеток сильно варьирует
 Митоз – деление клеток с предшествующим удвоением хромосом, в результате которого образуется две идентичных клетки с полноценным набором органоидов «Открытый» и «закрытый» митозы Митоз: нет синтеза, все структуры клетки «сосредоточены» на распределении между дочерними клетками и разделении клеток Должны разделиться все органоиды Перед митозом: в период G 2 завершены репликация ДНК и удвоение ядерных пор, ЭПР представлен цистернами и трубочками При переходе к митозу цистерны ЭПР исчезают, остаются трубочки Спирализация хромосом
Митоз – деление клеток с предшествующим удвоением хромосом, в результате которого образуется две идентичных клетки с полноценным набором органоидов «Открытый» и «закрытый» митозы Митоз: нет синтеза, все структуры клетки «сосредоточены» на распределении между дочерними клетками и разделении клеток Должны разделиться все органоиды Перед митозом: в период G 2 завершены репликация ДНК и удвоение ядерных пор, ЭПР представлен цистернами и трубочками При переходе к митозу цистерны ЭПР исчезают, остаются трубочки Спирализация хромосом
 Фазы митоза интерфаза прометафаза з цитокинез телофаза анафаза
Фазы митоза интерфаза прометафаза з цитокинез телофаза анафаза
 Дезинтеграция ядерной оболочки: - распадаются ядерные поры, - внешняя мембрана остается") Начало митоза (профаза) Дезинтеграция ядерной оболочки: - распадаются ядерные поры, - внешняя мембрана остается в составе ЭПР, - фрагменты внутренней мембраны присоединяются к трубочкам ЭПР - Диссоциация ядерных пор Компоненты пор диспергированы в цитоплазме, 3 нуклеопорина встраиваются в ЭПР - Деполимеризация ядерной пластинки - Спирализация и компактизация хромосом Метафаза: максимально компактизированные хромосомы выстраиваются по экватору клетки.
Начало митоза (профаза) Дезинтеграция ядерной оболочки: - распадаются ядерные поры, - внешняя мембрана остается в составе ЭПР, - фрагменты внутренней мембраны присоединяются к трубочкам ЭПР - Диссоциация ядерных пор Компоненты пор диспергированы в цитоплазме, 3 нуклеопорина встраиваются в ЭПР - Деполимеризация ядерной пластинки - Спирализация и компактизация хромосом Метафаза: максимально компактизированные хромосомы выстраиваются по экватору клетки.
 Анафаза: расхождение хромосом к полюсам клетки при помощи митотического веретена. Трубочки ЭПР связываются с хроматином и нуклеопоринами, ассоциированными с хроматином. Из ЭПР к хроматину рекрутируются мембранные трубочки и белки ядерной оболочки, формируется ядрышковый организатор. Большинство белков ядерной оболочки покидает ЭПР. Поздняя анафаза/ранняя телофаза: формируется замкнутая ядерная оболочка с порами. Сначала восстанавливается внутренний листок ядерной оболочки, затем собираются поры. Телофаза: завершение расхождения хромосом, оформление ядер дочерних клеток. Цитокинез
Анафаза: расхождение хромосом к полюсам клетки при помощи митотического веретена. Трубочки ЭПР связываются с хроматином и нуклеопоринами, ассоциированными с хроматином. Из ЭПР к хроматину рекрутируются мембранные трубочки и белки ядерной оболочки, формируется ядрышковый организатор. Большинство белков ядерной оболочки покидает ЭПР. Поздняя анафаза/ранняя телофаза: формируется замкнутая ядерная оболочка с порами. Сначала восстанавливается внутренний листок ядерной оболочки, затем собираются поры. Телофаза: завершение расхождения хромосом, оформление ядер дочерних клеток. Цитокинез
 Митоз Профаза Метафаза Прометафаза анафаза Телофаза
Митоз Профаза Метафаза Прометафаза анафаза Телофаза
 Период G 2: завершены репликация ДНК и удвоение ядерных пор, ЭПР представлен цистернами и трубочками. Митоз: ядерные поры «разобраны» , компоненты ядерной оболочки интегрированы в ЭПР, ЭПР представлен трубочками. Компоненты пор диспергированы в цитоплазме, три трансмембранных нуклеопорина вместе с белками оболочки ядра встраиваются в ЭПР. Центриоли локализуются у ядерной оболочки, микротрубочки участвуют в разрушении ядерной оболочки. Метафаза: часть компонентов ядерной пластинки связывается с кинетохором и формируется веретено. На этой стадии вокруг хромосом нет мембран.
Период G 2: завершены репликация ДНК и удвоение ядерных пор, ЭПР представлен цистернами и трубочками. Митоз: ядерные поры «разобраны» , компоненты ядерной оболочки интегрированы в ЭПР, ЭПР представлен трубочками. Компоненты пор диспергированы в цитоплазме, три трансмембранных нуклеопорина вместе с белками оболочки ядра встраиваются в ЭПР. Центриоли локализуются у ядерной оболочки, микротрубочки участвуют в разрушении ядерной оболочки. Метафаза: часть компонентов ядерной пластинки связывается с кинетохором и формируется веретено. На этой стадии вокруг хромосом нет мембран.
 Анафаза: трубочки ЭПР ассоциируют с хроматином и нуклеопоринами, связанными с хроматином. В эту стадию вовлечены мелкие ГТФазы Ran и фосфатазы. Из ЭПР рекрутируются мембранные трубочки и белки ядерной оболочки к хроматину, формируется ЯО. На этой стадии большинство белков ядерной оболочки покидает ЭПР. Поздняя анафаза/ранняя телофаза: формируется замкнутая ядерная оболочка с порами.
Анафаза: трубочки ЭПР ассоциируют с хроматином и нуклеопоринами, связанными с хроматином. В эту стадию вовлечены мелкие ГТФазы Ran и фосфатазы. Из ЭПР рекрутируются мембранные трубочки и белки ядерной оболочки к хроматину, формируется ЯО. На этой стадии большинство белков ядерной оболочки покидает ЭПР. Поздняя анафаза/ранняя телофаза: формируется замкнутая ядерная оболочка с порами.
 Цикл «размножения» есть у всех органоидов. Дупликация органелл - сложный процесс, он должен обеспечить удвоение митохондрий и пластид, окруженных двумя мембранами, и удвоение одномембранных органоидов, включая аппарат Гольджи и ЭПР. Митохондрии делятся перетяжкой, в них также происходит дупликация генома Аппарат Гольджи: разделение идет по центру от цис-стороны в сторону транс-стороны, образуется два дочерних аппарата Гольджи При «закрытом» митозе ЭПР делится вместе с ядром. Разделение ЭПР при «открытом» митозе пока не понято. Разделение органоидов между клетками происходит без «специальных» структур
Цикл «размножения» есть у всех органоидов. Дупликация органелл - сложный процесс, он должен обеспечить удвоение митохондрий и пластид, окруженных двумя мембранами, и удвоение одномембранных органоидов, включая аппарат Гольджи и ЭПР. Митохондрии делятся перетяжкой, в них также происходит дупликация генома Аппарат Гольджи: разделение идет по центру от цис-стороны в сторону транс-стороны, образуется два дочерних аппарата Гольджи При «закрытом» митозе ЭПР делится вместе с ядром. Разделение ЭПР при «открытом» митозе пока не понято. Разделение органоидов между клетками происходит без «специальных» структур
 Вольты - Vaults Вольты – красные Микротрубочки – зеленые
Вольты - Vaults Вольты – красные Микротрубочки – зеленые
 крио. ЭМ Вольты - Vault Размеры около 40 х 70 нм, толщина стенки – 20 А Самые крупные РНП-частицы Около 10 000 на клетку у млекопитающих
крио. ЭМ Вольты - Vault Размеры около 40 х 70 нм, толщина стенки – 20 А Самые крупные РНП-частицы Около 10 000 на клетку у млекопитающих
 маленькая нетранслируемая РНК Три белка: основной белок (MVP) 110 к.") Вольты: Одна (? ) маленькая нетранслируемая РНК Три белка: основной белок (MVP) 110 к. Да, два минорных белка: 193 к. Да поли(АДФ-рибоза) полимераза (v. PARP), и белок 240 к. Да (TEP-1), связанный с теломеразой, идентичен белку теломеразного комплекса ТЕР 1. Связывает РНК вольтов, необходим для их стабильности. Вольты локализуются в цитоплазме клеток, перемещаются путем диффузии. Показана связь вольтов с поровыми комплексами ядра. Полагают, что центральная ядерных пробка пор представлена именно вольтами, но это не доказано. Возможные функции вольтов: участие в транспортных механизмах, передаче сигналов и развитии иммунного ответа, формирование множественной лекарственной устойчивости. Уровень экспрессии основного белка вольтов – прогностический маркер многих видов рака и устойчивости к радиотерапии.
Вольты: Одна (? ) маленькая нетранслируемая РНК Три белка: основной белок (MVP) 110 к. Да, два минорных белка: 193 к. Да поли(АДФ-рибоза) полимераза (v. PARP), и белок 240 к. Да (TEP-1), связанный с теломеразой, идентичен белку теломеразного комплекса ТЕР 1. Связывает РНК вольтов, необходим для их стабильности. Вольты локализуются в цитоплазме клеток, перемещаются путем диффузии. Показана связь вольтов с поровыми комплексами ядра. Полагают, что центральная ядерных пробка пор представлена именно вольтами, но это не доказано. Возможные функции вольтов: участие в транспортных механизмах, передаче сигналов и развитии иммунного ответа, формирование множественной лекарственной устойчивости. Уровень экспрессии основного белка вольтов – прогностический маркер многих видов рака и устойчивости к радиотерапии.
") ПРОТЕАСОМА В клетках существует две системы деградации белков: лизосомы и протеасомы, обеспечивающие расщепление (протеолиз) белковых макромолекул. Поздние эндосомы и лизосомы – деградация белков, связанных с мембранами Протеасомы – деградация немембранных белков В клетке находится несколько тысяч протеасом, расположенных в нуклеоплазме и цитоплазме Протеасомы в клетках He. La
ПРОТЕАСОМА В клетках существует две системы деградации белков: лизосомы и протеасомы, обеспечивающие расщепление (протеолиз) белковых макромолекул. Поздние эндосомы и лизосомы – деградация белков, связанных с мембранами Протеасомы – деградация немембранных белков В клетке находится несколько тысяч протеасом, расположенных в нуклеоплазме и цитоплазме Протеасомы в клетках He. La
 Протеасома в электронном микроскопе
Протеасома в электронном микроскопе
 26 S – вся протеасома с каталитической активностью 20 S – сердцевина 1. Активация убиквитина в присутствии АТФ 2. Убиквитин связывается с молекулой, которую нужно уничтожить, но внутрь протеасомы не попадает
26 S – вся протеасома с каталитической активностью 20 S – сердцевина 1. Активация убиквитина в присутствии АТФ 2. Убиквитин связывается с молекулой, которую нужно уничтожить, но внутрь протеасомы не попадает
 Протеасома: Треониновые протеазы, каталитические центры – внутри Центральный кор, состоящий из 14 белковых димеров, по 7 в кольцевой структуре, всего 4 кольца, расположенных друг над другом; Две регуляторные части с двух сторон от кора, 14 белков в каждой, 6 из которых ATP-азы. Сборка протеасомы
Протеасома: Треониновые протеазы, каталитические центры – внутри Центральный кор, состоящий из 14 белковых димеров, по 7 в кольцевой структуре, всего 4 кольца, расположенных друг над другом; Две регуляторные части с двух сторон от кора, 14 белков в каждой, 6 из которых ATP-азы. Сборка протеасомы
 10 нм 20 S протеасома Thermoplasma
10 нм 20 S протеасома Thermoplasma