Лекция 10 Метаболизм липидов.ppt
- Количество слайдов: 53



Лекция 10. Метаболизм липидов.

Желчные кислоты

Вторичные желчные кислоты
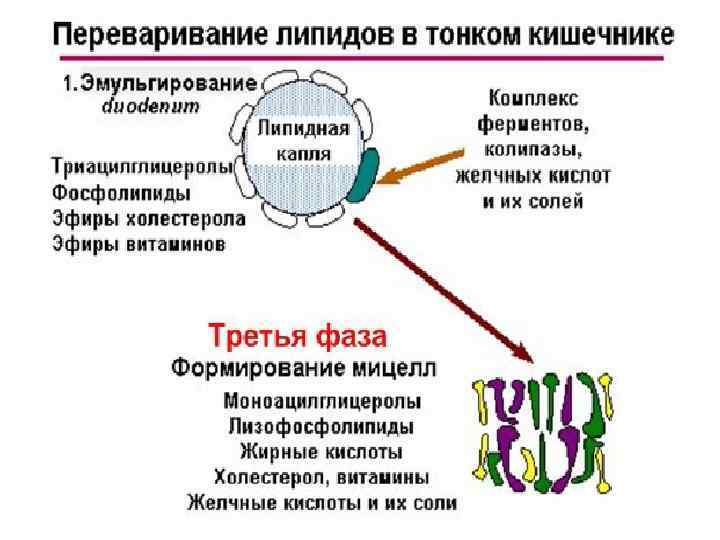
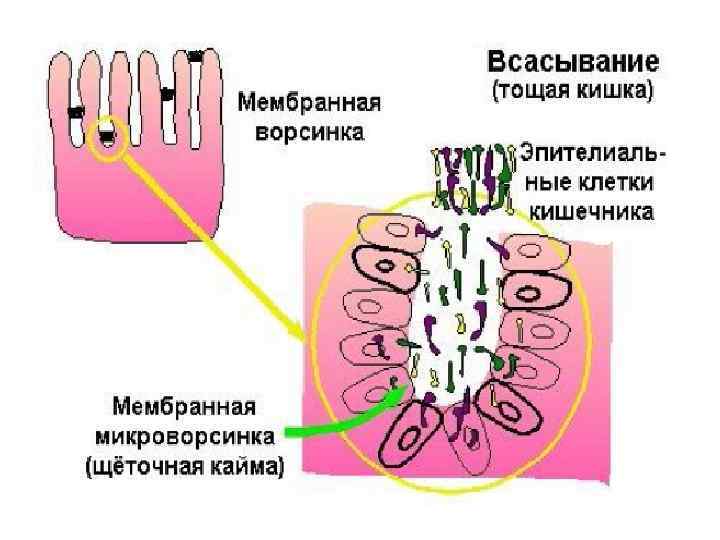

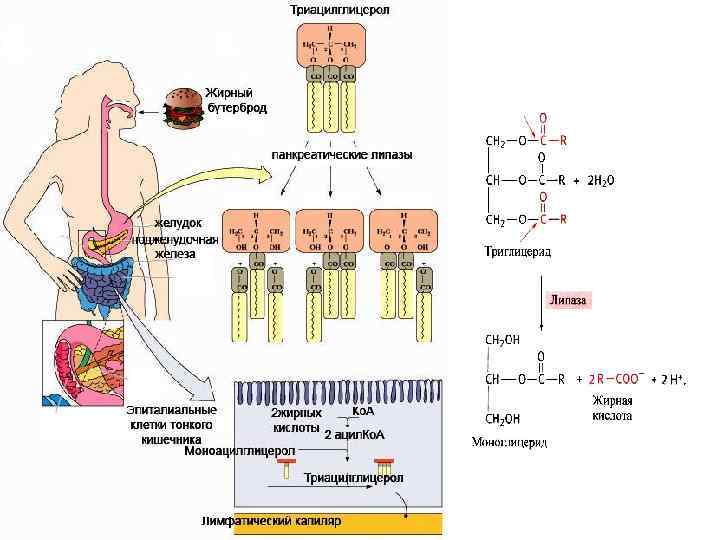
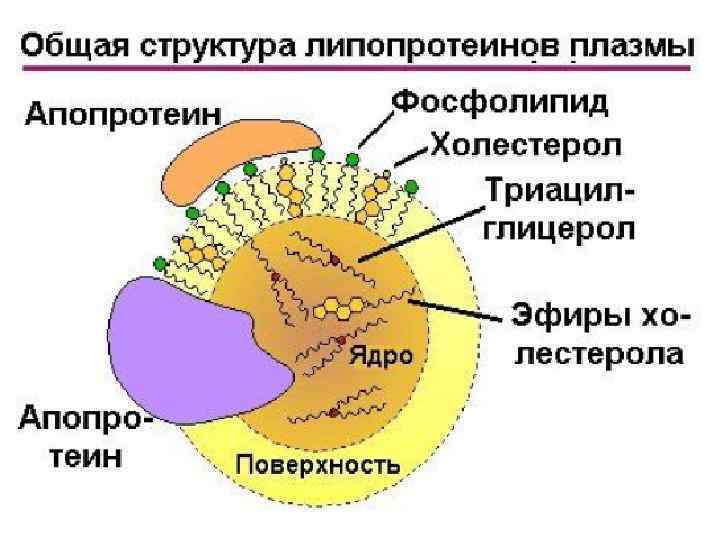
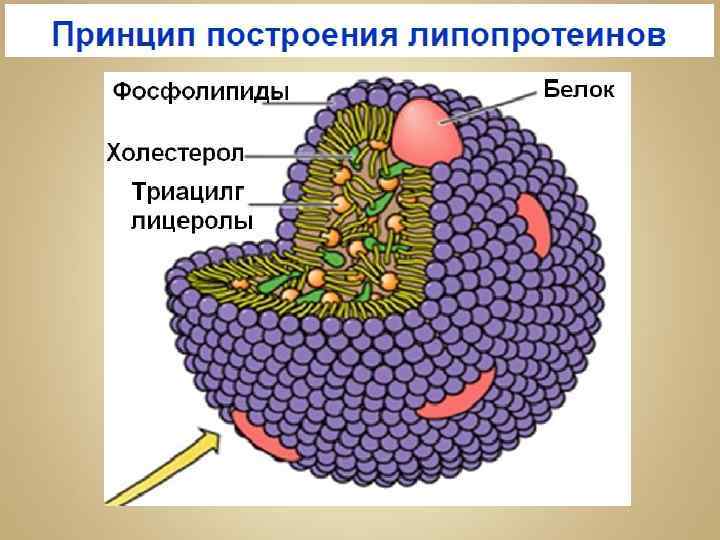
Хиломикроны Поверхность хиломикрона формируется молекулами фосфолипидов, холестерина и особыми белками - аполипопротеинами. Гидрофильная оболочка стабилизирует хиломикроны настолько, что они разносятся кровью и лимфой по организму как целые частицы.

Хиломикроны попадают из эпителиальных клеток не в кровь, а в лимфатические капилляры, также пронизывающие клеточные ворсинки эпителия кишечника. Что такое лимфа? Все ткани пронизаны сетью лимфатических капилляров, которые своими окончаниями собирают лимфу из межклеточного пространства. Система лимфатических капилляров и сосудов завершается грудным протоком, через который лимфа поступает в вены шеи. После приема жирной пищи, благодаря высокому содержанию хиломикронов, при смешивании лимфы с кровью, последняя мутнеет и начинает опалесцировать.

Функционирование хиломикронов

Разные контейнеры для транспорта липидов 1 – Хиломикроны 2 – ЛПОНП Липопротеины очень низкой плотности 2 1 3 3 – ЛПНП Липопротеины низкой плотности 4 4 – ЛПВП Липопротеины высокой плотности
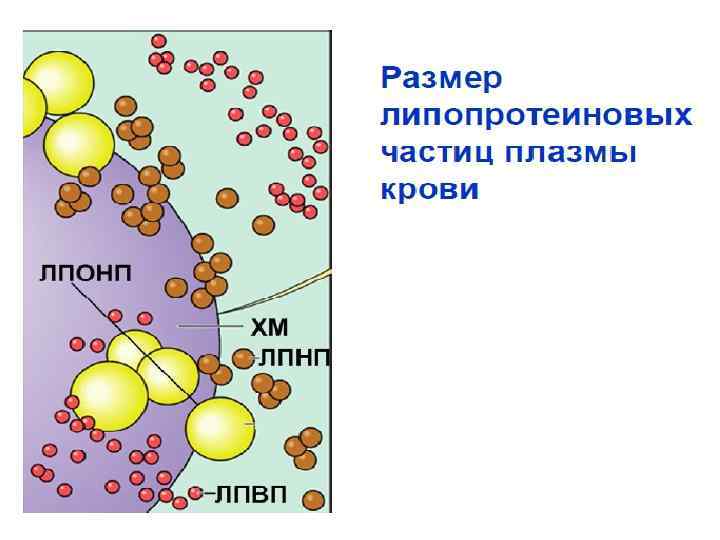

Сравнительные размеры липопротеинов


Липопротеины – ЛПОНП, ЛПВП синтезируются в печени. Печень не только собирает холестерин пищи, поступающий в нее в составе остатков хиломикронов, но является основным местом синтеза холестерина в организме. Из печени холестерин вместе с триглицеридами разносится по всему телу в составе ЛПОНП и ЛПНП. Вместе с тем существует и встречный поток холестерина из периферических тканей в печень. Это исходит благодаря ЛПВП. Они образуются в основном в печени и представляют собой дископодобную структру, состоящую из апобелков, фосфолипидов и относительно малого количества триглицеридов и холестерина. В периферических тканях ЛПВП обогащаются холестерином, благодаря диффузии последнего из плазматических мембран в прикрепляющийся к ним липопротеин.
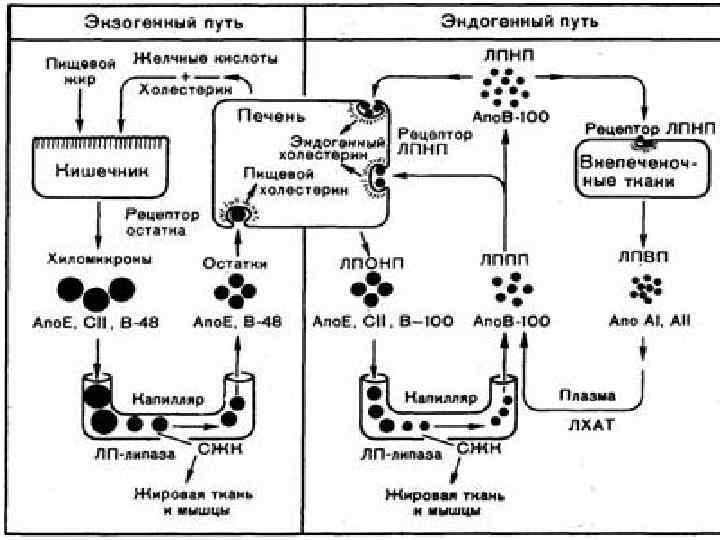

Транспорт холестерина и триацилглицеридов

Обмен липидов

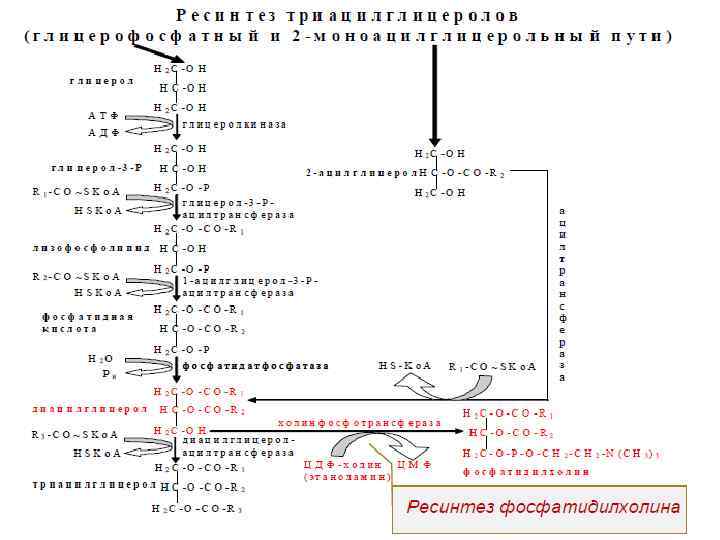
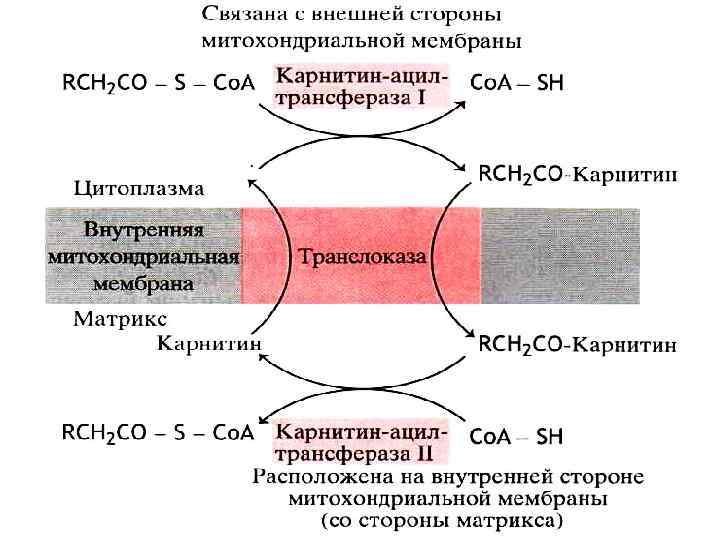
Трансформация и перемещение жирных кислот внутри клетки 1. В клетке жирные кислоты активируются, соединяясь Ко. А. Катализируют эту реакцию три родственных фермента – ацил. Ко. А-синтетазы жирных кислот. Эти ферменты специализируются в отношении коротких, средних и длинных жирных кислот. 2. Транспорт жирных кислот осуществляется с помощью карнитина
 1. Активация жирной кислоты 2.")
Окисление жирных кислот с четным числом атомов углерода (β-окисление) 1. Активация жирной кислоты 2. Транспорт жирной кислоты внутрь митохондрий

3. Внутримитохондриальное окисление жирных кислот 3. 1. Окисление ацил-Ко. А 3. 2. Гидратация

3. 3. Дегидрирование 3. 4. Тиолазная реакция

Окисление жирной кислоты – «Спираль Линена»

Энергетика окисления жирных кислот За один цикл β-окисления образуется 1 молекула ацетил-Со. А, окисление которого в цитратном цикле обеспечивает синтез 12 моль ATP. Кроме того, образуется 1 моль FADH 2 и 1 моль NADH+H, при окислении которых в дыхательной цепи синтезируется соответственно 2 и 3 моль ATP (в сумме 5). Таким образом, при окислении, например, пальмитиновой кислоты (С 16) происходит 7 циклов β -окисления, в результате которых образуется 8 моль ацетил-Со. А, 7 моль FADH 2 и 7 моль NADH+H. Следовательно, выход ATP составляет 35 молекул в результате β-окисления и 96 ATP в результате цитратного цикла, что соответствует в сумме 131 молекул АТФ.

Суммарное уравнение β-окисления пальмитиновой кислоты Пальмитоил-Ко. А + 7 ФАД + 7 НАД+ + 7 Н 2 O + 7 HS-Ko. A –> –> 8 Ацетил-Ко. А + 7 ФАДН 2 + 7 НАДН + 7 Н+. Изменение свободной энергии ΔF при полном сгорании 1 моля пальмитиновой кислоты составляет 2338 ккал, а богатая энергией фосфатная связь ATP характеризуется величиной 7, 6 ккал/моль. Нетрудно подсчитать, что примерно 990 ккал (7, 6 х 130), или 42% от всей потенциальной энергии пальмитиновой кислоты при ее окислении в организме, используется для ресинтеза АТР, а оставшаяся часть теряется в виде тепла.

Окисление жирных кислот с нечетным числом углеродных атомов

Синтез жирных кислот Образование ацетил-Со. А из лимонной кислоты Образование малонил-Со. А

Активные группы синтазы жирных кислот

Синтез жирных кислот

Регуляция синтеза жирных кислот

Синтез кетоновых тел в печени

Утилизация кетоновых тел в периферических тканях

Функции холестерина
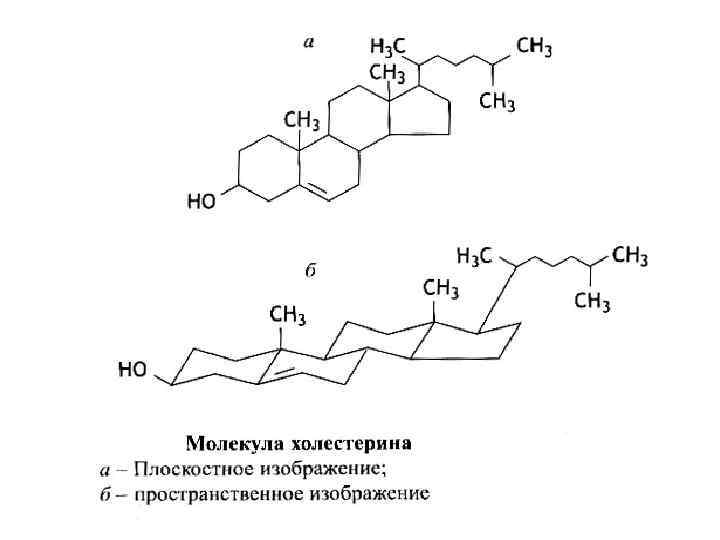

Холестерол еще раз

Биосинтез холестерола Начало 1 стадии: Три молекулы ацетил-Ко. А конденсируются с образованием ГОМГ-Ко. А (гаммаоксимасляная кислота)

Продолжение 1 стадии

Начало 2 стадии

Продолжение 2 стадии

Начало 3 стадии

Окончание 3 стадии

Транспорт холестерола

Эйкозаноиды, включающие в себя простагландины, тромбоксаны, лейкотриены - высокоактивные регуляторы клеточных функций. Они оказывают эффекты гормонов местного действия. Эйкозаноиды участвуют во многих процессах: на состояние бронхов, кишечника, матки. Эйкозаноиды регулируют секрецию воды и натрия почками, влияют на образование тромбов. Разные типы эйкозаноидов участвуют в развитии воспалительного процесса, происходящего после повреждения тканей или инфекции. Такие признаки воспаления, как боль, отёк, лихорадка, в значительной мере обусловлены действием эйкозаноидов. Избыточная секреция эйкозаноидов приводит к ряду заболеваний, например бронхиальной астме и аллергическим реакциям.

Синтез полиеновых жирных кислот с 20 углеродными атомами в организме человека

Семейства простагландинов

Структура тромбоксанов ТХ А 2 синтезируется из арахидоновои кислоты; ТХ А 3 синтезируется из эйкозапентаёновой кислоты.

Синтез эйкозаноидов из арахидоновой кислоты

Характеристика биологического действия эйкозаноидов Эйкозаноид Основное место синтеза Основное биологическое действие PG E 2 Большинство тканей, особенно почки Расслабляет гладкую мускулатуру, расширяет сосуды, инициирует родовую активность, подавляет миграцию лимфоцитов, пролиферацию Т-клеток. PG F 2α Большинство тканей Сокращает гладкую мускулатуру, суживает сосуды, бронхи, стимулирует сокращения матки. PG D 3 Клетки гладкой мускулатуры Вызывает расширение сосудов, снижает агрегацию тромбоцитов и лейкоцитов. PG I 2 Сердце, клетки эндотелия сосудов Уменьшает агрегацию тромбоцитов, расширяет сосуды. В клеткахмишенях увеличивает образование ц. АМФ. TX A 2 Тромбоциты Стимулирует агрегацию тромбоцитов, суживает сосуды и бронхи, в клетках уменьшает образование ц. АМФ. TX A 3 Тромбоциты Обладает функциями, одинаковыми с ТХ А 2, но значительно менее эффективен. LT B 4 Клетки белой крови, клетки эпителия Стимулирует хемотаксис и агрегацию лейкоцитов, освобождение лизосомальных ферментов лейкоцитов. Увеличивает проницаемость сосудов. Группа лейкотриенов Клетки белой крови, альвеолярные Стимулируют расширение сосудов, увеличивают их проницаемость. Вызывают сокращение бронхов. LTC 4→ макрофаги Основные компоненты «медленно реагирующей LT D 4→ субстанции» анафилаксии. LTE 4→ LXA 4 Лейкоциты Активирует хемотаксис и стимулирует образование супероксид аниона в лейкоцитах.

ПЕРЕКИСНОЕ ОКИСЛЕНИЕ ЛИПИДОВ ПОЛ - цепные реакции, обеспечивающие расширенное воспроизводство свободных радикалов, частиц, имеющих неспаренный электрон, которые инициируют дальнейшее распространение перекисного окисления. Инициация: образование свободного радикала (L • ) Развитие цепи L • + О 2 → LOO • + LH → LOOM + LR • Разрушение структуры липидов Обрыв цепи - взаимодействие радикалов между собой
Лекция 10 Метаболизм липидов.ppt