

Lektsia_1_Vvedenie_Protisty_Sarkodovye.ppt
- Количество слайдов: 78
 Лекция 1. Предмет и задачи зоологии
Лекция 1. Предмет и задачи зоологии
 Зачем Вам нужна зоология: 1. Будете знать точные названия животных, а также строение их тела и названия отдельных его частей (формальный повод и причина хорошо поработать). 2. Беспозвоночные животные имеют огромное значение в природе и в жизни человека (практическое значение): • они используются в пищу многими позвоночными животными, в том числе человеком; • многие беспозвоночные - сельскохозяйственные вредители; • многие беспозвоночные паразиты (споровики, нематоды, цестоды) и переносчики опасных заболеваний человека и животных (клещи, комары); 3. Познание основ биологии беспозвоночных привело к фундаментальным открытиям в биологии человека, • Илья Мечников, основавший иммунологию, в 1883 г. открыл явление фагоцитоза и показал его роль в формировании иммунитета провел простые наблюдения за ползающими амебоцитами из личинок морских звезд (за это он позднее получил Нобелевскую премию) • беспозвоночные, как модельные объекты (нематоды и плодовые мушки дрозофилы) помогли нам понять функции генов и их роли в возникновении заболеваний человека
Зачем Вам нужна зоология: 1. Будете знать точные названия животных, а также строение их тела и названия отдельных его частей (формальный повод и причина хорошо поработать). 2. Беспозвоночные животные имеют огромное значение в природе и в жизни человека (практическое значение): • они используются в пищу многими позвоночными животными, в том числе человеком; • многие беспозвоночные - сельскохозяйственные вредители; • многие беспозвоночные паразиты (споровики, нематоды, цестоды) и переносчики опасных заболеваний человека и животных (клещи, комары); 3. Познание основ биологии беспозвоночных привело к фундаментальным открытиям в биологии человека, • Илья Мечников, основавший иммунологию, в 1883 г. открыл явление фагоцитоза и показал его роль в формировании иммунитета провел простые наблюдения за ползающими амебоцитами из личинок морских звезд (за это он позднее получил Нобелевскую премию) • беспозвоночные, как модельные объекты (нематоды и плодовые мушки дрозофилы) помогли нам понять функции генов и их роли в возникновении заболеваний человека
 Зачем Вам нужна зоология: 3. Получите удовольствие от разнообразия строения и возможностей беспозвоночных (эстетическое значение) Многие беспозвоночные потрясающе красивы Впечатляюще большие размеры тела: • Крупнейшие из ныне живущих беспозвоночных достигают в длину 19 м, весят до 500 кг и могут плавать в воде со скоростью до 25 км/ч (гигантский кальмар)
Зачем Вам нужна зоология: 3. Получите удовольствие от разнообразия строения и возможностей беспозвоночных (эстетическое значение) Многие беспозвоночные потрясающе красивы Впечатляюще большие размеры тела: • Крупнейшие из ныне живущих беспозвоночных достигают в длину 19 м, весят до 500 кг и могут плавать в воде со скоростью до 25 км/ч (гигантский кальмар)
 Земляные черви") Размеры беспозвоночных Крабы могут иметь размах ног до 4 м! (Японский краб-паук) Земляные черви могут быть более 3 м в длину! (Гигантский австралийский земляной червь)
Размеры беспозвоночных Крабы могут иметь размах ног до 4 м! (Японский краб-паук) Земляные черви могут быть более 3 м в длину! (Гигантский австралийский земляной червь)
 – основатель зоологии.") Аристотель (384 до н. э. – 322 до н. э. ) – основатель зоологии. Он дал описание 454 видов животных. Он разделил животных на 2 большие группы - животных, снабженных кровью (позвоночные, по современным представлениям), и бескровных (беспозвоночные, в современном понимании). Животные с кровью в свою очередь были разделены им на группы, приблизительно отвечающие современным классам. В отношении беспозвоночных система Аристотеля была менее совершенна. Так, из современных типов он более или менее правильно выделял лишь членистоногих. Последующие 2 тыс. лет не внесли в систему животных ничего существенно нового. Аристотель
Аристотель (384 до н. э. – 322 до н. э. ) – основатель зоологии. Он дал описание 454 видов животных. Он разделил животных на 2 большие группы - животных, снабженных кровью (позвоночные, по современным представлениям), и бескровных (беспозвоночные, в современном понимании). Животные с кровью в свою очередь были разделены им на группы, приблизительно отвечающие современным классам. В отношении беспозвоночных система Аристотеля была менее совершенна. Так, из современных типов он более или менее правильно выделял лишь членистоногих. Последующие 2 тыс. лет не внесли в систему животных ничего существенно нового. Аристотель
 Дал первую биологическую концепцию вида.") Джон Рэй (1627 – 1705) Дал первую биологическую концепцию вида.
Джон Рэй (1627 – 1705) Дал первую биологическую концепцию вида.
 Линней заложил основы современной биноминальной (бинарной) номенклатуры, введя в") Карл Линней (1707 – 1778) Линней заложил основы современной биноминальной (бинарной) номенклатуры, введя в практику систематики так называемые nomina trivialia, которые позже стали использоваться в качестве видовых эпитетов в биноминальных названиях живых организмов. Введённый Линнеем метод формирования научного названия для каждого из видов используется до сих пор (применявшиеся ранее длинные названия, состоящие из большого количества слов, давали описание видов, но не были строго формализованы). Использование латинского названия из двух слов — название рода, затем специфичное имя — позволило отделить номенклатуру от таксономии. Он использовал четыре уровня ( «ранга» ): классы, отряды, роды и виды. Общее число видов в 10 -м издании его «Системы природы» (т. 1 -2, 175859) превышало 4200 (в т. ч. 1222 вида позвоночных и 1936 видов насекомых).
Карл Линней (1707 – 1778) Линней заложил основы современной биноминальной (бинарной) номенклатуры, введя в практику систематики так называемые nomina trivialia, которые позже стали использоваться в качестве видовых эпитетов в биноминальных названиях живых организмов. Введённый Линнеем метод формирования научного названия для каждого из видов используется до сих пор (применявшиеся ранее длинные названия, состоящие из большого количества слов, давали описание видов, но не были строго формализованы). Использование латинского названия из двух слов — название рода, затем специфичное имя — позволило отделить номенклатуру от таксономии. Он использовал четыре уровня ( «ранга» ): классы, отряды, роды и виды. Общее число видов в 10 -м издании его «Системы природы» (т. 1 -2, 175859) превышало 4200 (в т. ч. 1222 вида позвоночных и 1936 видов насекомых).
 Две основные работы «Система беспозвоночных» (1801) и в «Философия") Жан-Батист Ламарк (1744 – 1829) Две основные работы «Система беспозвоночных» (1801) и в «Философия зоологии» (1809). Ламарк разделил всех животных на беспозвоночных и позвоночных. Впервые предложил термин беспозвоночные. Он различал классы инфузорий, полипов, лучистых (кишечнополостные и иглокожие), червей, насекомых, паукообразных, ракообразных, кольчецов, усоногих и моллюсков.
Жан-Батист Ламарк (1744 – 1829) Две основные работы «Система беспозвоночных» (1801) и в «Философия зоологии» (1809). Ламарк разделил всех животных на беспозвоночных и позвоночных. Впервые предложил термин беспозвоночные. Он различал классы инфузорий, полипов, лучистых (кишечнополостные и иглокожие), червей, насекомых, паукообразных, ракообразных, кольчецов, усоногих и моллюсков.
 Английский натуралист и путешественник, одним из первых осознал и") Чарльз Дарвин (1809 – 1802) Английский натуралист и путешественник, одним из первых осознал и наглядно продемонстрировал, что все виды живых организмов эволюционируют во времени от общих предков. В своей теории, первое развёрнутое изложение которой было опубликовано в 1859 году в книге «Происхождение видов» (полное название: «Происхождение видов путём естественного отбора, или сохранение благоприятствуемых пород в борьбе за жизнь» ), основной движущей силой эволюции Дарвин назвал естественный отбор и неопределённую изменчивость.
Чарльз Дарвин (1809 – 1802) Английский натуралист и путешественник, одним из первых осознал и наглядно продемонстрировал, что все виды живых организмов эволюционируют во времени от общих предков. В своей теории, первое развёрнутое изложение которой было опубликовано в 1859 году в книге «Происхождение видов» (полное название: «Происхождение видов путём естественного отбора, или сохранение благоприятствуемых пород в борьбе за жизнь» ), основной движущей силой эволюции Дарвин назвал естественный отбор и неопределённую изменчивость.
 Сравнительная характеристика клеток прокариот и эукариот
Сравнительная характеристика клеток прокариот и эукариот
 Происхождение эукариотической клетки Эукариоты возникли -2 млрд. лет т. н. Эндосимбиотическая теория – Линн Маргулис (1981) Эукариоты возникали полифилитически (неоднократно и независимо в разное время и во многих местах планеты) Lynn Margulis
Происхождение эукариотической клетки Эукариоты возникли -2 млрд. лет т. н. Эндосимбиотическая теория – Линн Маргулис (1981) Эукариоты возникали полифилитически (неоднократно и независимо в разное время и во многих местах планеты) Lynn Margulis
") Филогенетические взаимоотношения между основными группами живых организмов (по: Whittaker, 1969)
Филогенетические взаимоотношения между основными группами живых организмов (по: Whittaker, 1969)
") Происхождение эукариотической клетки Константин Мережковский Предложил термин «симбиогенез» «Растения, рассматриваемые как симбиотический комплекс» (1920) Lynn Margulis Sagan Lynn. 1967. On the origin of mitosing cells // J. Theor. Bio. 14 (3): 255– 274 Margulis, Lynn, 1970, Origin of Eukaryotic Cells, Yale University Press
Происхождение эукариотической клетки Константин Мережковский Предложил термин «симбиогенез» «Растения, рассматриваемые как симбиотический комплекс» (1920) Lynn Margulis Sagan Lynn. 1967. On the origin of mitosing cells // J. Theor. Bio. 14 (3): 255– 274 Margulis, Lynn, 1970, Origin of Eukaryotic Cells, Yale University Press
 Основные признаки царств эукариотических организмов Животные – многоклеточные гетеротрофы с голозойным питанием; кристы в митохондриях пластинчатые; обычно имеют сложное эмбриональное развитие; мейоз гаметический Растения – преобладают многоклеточные автотрофы с хлорофиллом а и b; в основном наземные; клеточная стенка из целлюлозы, запасы крахмала в хлоропластах; кристы митохондрий пластинчатые; преобладает спорический тип мейоза. Грибы – мицелиальные гетеротрофы с сапрофитным питанием; клеточная стенка из хитина; кристы митохондрий пластинчатые; зиготический мейоз Протисты – в основном одноклеточные, микроскопические, имеются также крупные синцитиальные формы, ценобии и многоклеточные; движение при помощи жгутиков, ресничек, псевдоподий; питание автотрофное, гетеротрофное, миксотрофное; формы мейоза – во всех вариантах; кристы митохондрий различной формы.
Основные признаки царств эукариотических организмов Животные – многоклеточные гетеротрофы с голозойным питанием; кристы в митохондриях пластинчатые; обычно имеют сложное эмбриональное развитие; мейоз гаметический Растения – преобладают многоклеточные автотрофы с хлорофиллом а и b; в основном наземные; клеточная стенка из целлюлозы, запасы крахмала в хлоропластах; кристы митохондрий пластинчатые; преобладает спорический тип мейоза. Грибы – мицелиальные гетеротрофы с сапрофитным питанием; клеточная стенка из хитина; кристы митохондрий пластинчатые; зиготический мейоз Протисты – в основном одноклеточные, микроскопические, имеются также крупные синцитиальные формы, ценобии и многоклеточные; движение при помощи жгутиков, ресничек, псевдоподий; питание автотрофное, гетеротрофное, миксотрофное; формы мейоза – во всех вариантах; кристы митохондрий различной формы.
 Левенгук и его микроскоп Антони ван Левенгук В 1675 г. описал первых простейших, назвав их «анималькули»
Левенгук и его микроскоп Антони ван Левенгук В 1675 г. описал первых простейших, назвав их «анималькули»
 Современные методы микроскопии
Современные методы микроскопии
 Общая характеристика протистов • В настоящее время насчитывается от 120 до 200 тыс. видов протистов, причем это меньшая часть их многообразия в биосфере. Они включают большое число разнообразных таксонов, которые не формируют монофилетических групп. • Все они – одноклеточные эукариоты. Лишены клеточной стенки и имеют по крайне мере одну подвижную стадию в жизненном цикле и большинство питаются анимально (заглатывая пищу). Клетки простейших более крупные и более сложные, чем у прокариот, они содержат различные органеллы (Аппарат Гольджи, митохондрии, рибосомы и т. д. ). • Протисты обитают в природе во всех средах – океаны, моря, пресные и солоноватые воды, почвы, паразитируют во всех группах эукариот. • Занимают одно из ключевых мест в продукции и деструкции органических веществ, участвуют в цепях питания животных в морских и пресных водоемах. • Могут быть использованы в индикации органического загрязнения воды, применяются в биологической очистке сточных вод и в самоочищении водоемов.
Общая характеристика протистов • В настоящее время насчитывается от 120 до 200 тыс. видов протистов, причем это меньшая часть их многообразия в биосфере. Они включают большое число разнообразных таксонов, которые не формируют монофилетических групп. • Все они – одноклеточные эукариоты. Лишены клеточной стенки и имеют по крайне мере одну подвижную стадию в жизненном цикле и большинство питаются анимально (заглатывая пищу). Клетки простейших более крупные и более сложные, чем у прокариот, они содержат различные органеллы (Аппарат Гольджи, митохондрии, рибосомы и т. д. ). • Протисты обитают в природе во всех средах – океаны, моря, пресные и солоноватые воды, почвы, паразитируют во всех группах эукариот. • Занимают одно из ключевых мест в продукции и деструкции органических веществ, участвуют в цепях питания животных в морских и пресных водоемах. • Могут быть использованы в индикации органического загрязнения воды, применяются в биологической очистке сточных вод и в самоочищении водоемов.
 Форма тела и размеры üНаиболее разнообразна форма активно подвижных свободно живущих простистов. Это связано с характером движений, питания, особенностями среды обитания. üУ сидячих протистов появляются элементы радиальной симметрии. üУ многих протистов есть колониальные формы (линейные, пластинчатые, шаровидные, древовидные). üВстречаются также сетчатые плазмодии (огромные многоядерные особи без клеточных перегородок между ядрами). üРазмеры тела варьируют от 1 мкм у споровиков до десятков см у фораменифер. üПротисты бывают не только одноклеточные. üПри описании морфологии клеток протистов мы рассмотрим 3 системы: поверхностные структуры, цитоплазму и ядро.
Форма тела и размеры üНаиболее разнообразна форма активно подвижных свободно живущих простистов. Это связано с характером движений, питания, особенностями среды обитания. üУ сидячих протистов появляются элементы радиальной симметрии. üУ многих протистов есть колониальные формы (линейные, пластинчатые, шаровидные, древовидные). üВстречаются также сетчатые плазмодии (огромные многоядерные особи без клеточных перегородок между ядрами). üРазмеры тела варьируют от 1 мкм у споровиков до десятков см у фораменифер. üПротисты бывают не только одноклеточные. üПри описании морфологии клеток протистов мы рассмотрим 3 системы: поверхностные структуры, цитоплазму и ядро.
 Основной набор ультраструктурных признаков протистов • Структура покровов • Особенности цитоскелета (в основном жгутикового аппарата) • Структура псевдоподий • Строение хлоропластов • Форма крист митохондрий • Наличие или отсутствие диктиосом • Наличие уникальных органелл
Основной набор ультраструктурных признаков протистов • Структура покровов • Особенности цитоскелета (в основном жгутикового аппарата) • Структура псевдоподий • Строение хлоропластов • Форма крист митохондрий • Наличие или отсутствие диктиосом • Наличие уникальных органелл
 Поверхностные структуры В основе поверхностных структур протистов – трехслойная поверхностная мембрана с прилегающим к ней слоем гликокаликса. Однако покровы могут быть существенно усложнены: 1. За счет надмембранных слоев; 2. За счет изменения прилегающей к мембране цитоплазмы.
Поверхностные структуры В основе поверхностных структур протистов – трехслойная поверхностная мембрана с прилегающим к ней слоем гликокаликса. Однако покровы могут быть существенно усложнены: 1. За счет надмембранных слоев; 2. За счет изменения прилегающей к мембране цитоплазмы.
 Цитоскелет большинства эукариотических клеток состоит из трех филаментных систем Промежуточные филаменты 25 нм d=9 -12 нм Специфичные для разных типов клеток субъединицы белков. Эластичность и сопротивляемость растяжению Микрофиламенты 25 нм d=6 -7 нм Микротрубочки 25 nm d=25 нм Актин (миозин и дополнительные белки) Тубулины (и молекулярные моторы) Сокращение, транспорт, движение и деление Транспорт везикул, клеточная полярность и деление
Цитоскелет большинства эукариотических клеток состоит из трех филаментных систем Промежуточные филаменты 25 нм d=9 -12 нм Специфичные для разных типов клеток субъединицы белков. Эластичность и сопротивляемость растяжению Микрофиламенты 25 нм d=6 -7 нм Микротрубочки 25 nm d=25 нм Актин (миозин и дополнительные белки) Тубулины (и молекулярные моторы) Сокращение, транспорт, движение и деление Транспорт везикул, клеточная полярность и деление
 занимают в основном") Строение цитоскелета по данным флюоресцентной микроскопии Актиновые микрофиламенты (в ультрафиолете красные) занимают в основном периферическое положение в клетке, а микротрубочки (зеленые) – в основном радиальное, соединяют ядро клетки с периферией клетки.
Строение цитоскелета по данным флюоресцентной микроскопии Актиновые микрофиламенты (в ультрафиолете красные) занимают в основном периферическое положение в клетке, а микротрубочки (зеленые) – в основном радиальное, соединяют ядро клетки с периферией клетки.
 Строение и функции микрофиламентов • Ультратонкие филаменты, похожи на скрученное жемчужное ожерелье • Функционирование псевдоподий в амебоидных клетках • Плотная сеть под плазмалеммой поддерживает форму клетки • Разделение материнской клетки на две при митозе • Поддерживает структуру микровилей • Контролирует внутриклеточные движения: перемещения веществ • Важный компонент мускульного сокращения (вместе с миозином) внутри клетки и потоки цитоплазмы © 2010 Nature Education
Строение и функции микрофиламентов • Ультратонкие филаменты, похожи на скрученное жемчужное ожерелье • Функционирование псевдоподий в амебоидных клетках • Плотная сеть под плазмалеммой поддерживает форму клетки • Разделение материнской клетки на две при митозе • Поддерживает структуру микровилей • Контролирует внутриклеточные движения: перемещения веществ • Важный компонент мускульного сокращения (вместе с миозином) внутри клетки и потоки цитоплазмы © 2010 Nature Education
 Строение микрофиламентов
Строение микрофиламентов
 Строение и функции микротрубочек Полый цилиндр из двух белков a и b тубулинов Спонтанное объединение молекул a и b тубулина образуют структуры, называемые димерами Димеры собираются в спираль по 13 по димеров по кругу Под контролем микротрубочек образуется клеточный центр Взаимодействуюя с белками кинезином и динеином, могут перемещать органеллы
Строение и функции микротрубочек Полый цилиндр из двух белков a и b тубулинов Спонтанное объединение молекул a и b тубулина образуют структуры, называемые димерами Димеры собираются в спираль по 13 по димеров по кругу Под контролем микротрубочек образуется клеточный центр Взаимодействуюя с белками кинезином и динеином, могут перемещать органеллы
 Промежуточные филаменты
Промежуточные филаменты
 Цитоскелет
Цитоскелет
 Экструсомы. Трихоцисты Цитоплазматические органелы простистов, способных к «выстреливанию» при механическом или химическом раздражении. Веретенообразные Т. — органеллы защиты, характерные для инфузорий; имеют вид удлинённых (2— 6 мкм) белковых телец паракристаллические структуры. Обычно снабжены плотным остриём, располагаются в цитоплазме перпендикулярно поверхности тела (например, у парамеций; при выстреливании вытягиваются в нить длиной 20 -60 мкм с остриём на конце.
Экструсомы. Трихоцисты Цитоплазматические органелы простистов, способных к «выстреливанию» при механическом или химическом раздражении. Веретенообразные Т. — органеллы защиты, характерные для инфузорий; имеют вид удлинённых (2— 6 мкм) белковых телец паракристаллические структуры. Обычно снабжены плотным остриём, располагаются в цитоплазме перпендикулярно поверхности тела (например, у парамеций; при выстреливании вытягиваются в нить длиной 20 -60 мкм с остриём на конце.
 Экструсомы жгутиконосцев
Экструсомы жгутиконосцев
 Ядро
Ядро
 При открытом митозе в отличие от закрытого") Типы митоза у протистов (по: Райков, 1994) При открытом митозе в отличие от закрытого разрушается ядерная оболочка, ядрышки и хромосомы оказываются свободно лежащими в цитоплазме. При закрытом митозе ядерная оболочка сохраняется до завершения процесса деления ядра. При ортомитозе в метафазе хромосомы образуют метафазную экваториальную пластинку. При плевромитозе ее нет, а ахроматиновое веретено представлено двумя полуверетенами, лежащими под углом друг к другу, поэтому фигура деления асимметрична
Типы митоза у протистов (по: Райков, 1994) При открытом митозе в отличие от закрытого разрушается ядерная оболочка, ядрышки и хромосомы оказываются свободно лежащими в цитоплазме. При закрытом митозе ядерная оболочка сохраняется до завершения процесса деления ядра. При ортомитозе в метафазе хромосомы образуют метафазную экваториальную пластинку. При плевромитозе ее нет, а ахроматиновое веретено представлено двумя полуверетенами, лежащими под углом друг к другу, поэтому фигура деления асимметрична
 1 2 Строение крист митохондрий у протист 3 4 1, 4 Пластинчатые и дисковидные кристы жгутиконосца Euglena spirogyra 2 Форма крист митохондрий может быть использована как основа для построения системы протист. У протист выделяют три формы – дискокристаты, тубулокристаты, платикристаты. 3 Ветвистые кристы амебы Sappinia diploidea Трубчатые кристы инфузории Paramecium putrinum 4 В криптонах (производные митохондрий) дизентерийной амебы Entamoeba histolytica крипты отсутствуют (Ghoch et al. , 2000) Пластинчатые кристы флагелляты Stephanoeca diplocostata.
1 2 Строение крист митохондрий у протист 3 4 1, 4 Пластинчатые и дисковидные кристы жгутиконосца Euglena spirogyra 2 Форма крист митохондрий может быть использована как основа для построения системы протист. У протист выделяют три формы – дискокристаты, тубулокристаты, платикристаты. 3 Ветвистые кристы амебы Sappinia diploidea Трубчатые кристы инфузории Paramecium putrinum 4 В криптонах (производные митохондрий) дизентерийной амебы Entamoeba histolytica крипты отсутствуют (Ghoch et al. , 2000) Пластинчатые кристы флагелляты Stephanoeca diplocostata.
 Гидрогеносомы жгутиконосца Tritrichomonas foetus, связанные с элементами цитоскелета, – пример конвергентной эволюции с митохондриями эукариот. На нижнем фото видны двойные мембраны (о – наружная, i – внутренняя) a, d. Гидрогеносомы (H) Nyctotherus ovalis инфузории из кишечника Blaberus). Вокруг видны эндосимбиотические метано-продуцирующие археи (черные точки); N, макронуклеус; n, микронуклеус; V, вакуоль. b, c, e, N. ovalis из кишечника Periplaneta americana; m, метаногенные Archaea (эндосимбионты). На d ясно видны кристы. Стрелками показаны рибосомы. Масштаб на рис. b–d – 1 мкм, на а – 10 мкм. По: Akhmanova A. et al. A hydrogenosome with a genome // Nature. 1998. Vol. 396. P. 527– 528.
Гидрогеносомы жгутиконосца Tritrichomonas foetus, связанные с элементами цитоскелета, – пример конвергентной эволюции с митохондриями эукариот. На нижнем фото видны двойные мембраны (о – наружная, i – внутренняя) a, d. Гидрогеносомы (H) Nyctotherus ovalis инфузории из кишечника Blaberus). Вокруг видны эндосимбиотические метано-продуцирующие археи (черные точки); N, макронуклеус; n, микронуклеус; V, вакуоль. b, c, e, N. ovalis из кишечника Periplaneta americana; m, метаногенные Archaea (эндосимбионты). На d ясно видны кристы. Стрелками показаны рибосомы. Масштаб на рис. b–d – 1 мкм, на а – 10 мкм. По: Akhmanova A. et al. A hydrogenosome with a genome // Nature. 1998. Vol. 396. P. 527– 528.
 Хлоропласты
Хлоропласты
 клеточный органоид") Лизосомы Лизосома — (от греч. λύσις — растворяю и sōma — тело) клеточный органоид размером 0, 2 — 0, 4 мкм, один из видов везикул. Эти одномембранные органоиды — часть вакуома (эндомембранной системы клетки). Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты.
Лизосомы Лизосома — (от греч. λύσις — растворяю и sōma — тело) клеточный органоид размером 0, 2 — 0, 4 мкм, один из видов везикул. Эти одномембранные органоиды — часть вакуома (эндомембранной системы клетки). Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты.
 Пероксисомы Сходны с лизосомами Ограниченные мембраной пузырьки Содержат ферменты Однако Ферменты синтеризрованы свободными рибосомами (вне эндоплазматического ретикулюма) Активны в липидном метаболизме Катализируют реакции с образованием пероксида водорода H 2 O 2 О н токсичен Р азрушается до воды и O 2 каталазой
Пероксисомы Сходны с лизосомами Ограниченные мембраной пузырьки Содержат ферменты Однако Ферменты синтеризрованы свободными рибосомами (вне эндоплазматического ретикулюма) Активны в липидном метаболизме Катализируют реакции с образованием пероксида водорода H 2 O 2 О н токсичен Р азрушается до воды и O 2 каталазой
 Диффузия Гидрофильные молекулы и мелкие молекулы могут проникать через мембрану путем диффузии. Краситель диффундирует по градиенту концентрации из области большей концентрации в область меньшей. Со временем достигается динамическое равновесие: растворенные молекулы проникают через мембрану, но в равном количестве в обоих направлениях
Диффузия Гидрофильные молекулы и мелкие молекулы могут проникать через мембрану путем диффузии. Краситель диффундирует по градиенту концентрации из области большей концентрации в область меньшей. Со временем достигается динамическое равновесие: растворенные молекулы проникают через мембрану, но в равном количестве в обоих направлениях
 Формы эндоцитоза Фагоцитоз. Клетка заглатывает частицу, окружая ее псевдоподией и заключая внутри замкнутого мембранного пузырька, который называется пищеварительной вакуолью. Пищевая частица переваривается после проникновения в вакуоль содержимого лизосом. Пиноцитоз. Клетка «глотает» капли внеклеточной жидкости вместе с растворенными в ней молекулами в виде маленького пузырька. Поскольку при пиноцитозе все растворенные вещества попадают в клетку – это неспецифический транспорт вещества. Рецепторный эндоцитоз. Рецепторные белки включенные в мембрану могут связывать специфические внеклеточные вещества (лиганды). Связанный комплекс образует углубление, выстланное слоем мембранных белков, которые замыкают образующуюся вакуоль.
Формы эндоцитоза Фагоцитоз. Клетка заглатывает частицу, окружая ее псевдоподией и заключая внутри замкнутого мембранного пузырька, который называется пищеварительной вакуолью. Пищевая частица переваривается после проникновения в вакуоль содержимого лизосом. Пиноцитоз. Клетка «глотает» капли внеклеточной жидкости вместе с растворенными в ней молекулами в виде маленького пузырька. Поскольку при пиноцитозе все растворенные вещества попадают в клетку – это неспецифический транспорт вещества. Рецепторный эндоцитоз. Рецепторные белки включенные в мембрану могут связывать специфические внеклеточные вещества (лиганды). Связанный комплекс образует углубление, выстланное слоем мембранных белков, которые замыкают образующуюся вакуоль.
 вещества, попадающие в клетку и перерабатывают внеклеточные вещества. Молекулы могут") Фагоцитоз Лизосомы переваривают (гидролизуют) вещества, попадающие в клетку и перерабатывают внеклеточные вещества. Молекулы могут быть поглощены включением в цитоплазматическую мембрану или в пищеварительную вакуоль. Для переваривания пищи при фагоцитозе пищеварительная вакуоль сливается с лизосомами.
Фагоцитоз Лизосомы переваривают (гидролизуют) вещества, попадающие в клетку и перерабатывают внеклеточные вещества. Молекулы могут быть поглощены включением в цитоплазматическую мембрану или в пищеварительную вакуоль. Для переваривания пищи при фагоцитозе пищеварительная вакуоль сливается с лизосомами.
 Сократительная вакуоль и осморегуляция
Сократительная вакуоль и осморегуляция
 Сократительная вакуоль и осморегуляция
Сократительная вакуоль и осморегуляция
") Обобщенная схема формирования таксонов эукариот в эволюционном процессе (по: Patterson, 1994 из Протисты, 2001)
Обобщенная схема формирования таксонов эукариот в эволюционном процессе (по: Patterson, 1994 из Протисты, 2001)
 Три формы движение протистов
Три формы движение протистов
 Формы псевдоподий Слева направо: полиподиальные лобозные, моноподиальные лобозные, филозные, конические, гранулоретикулозные, конические актиноподии, неконические актиноподии.
Формы псевдоподий Слева направо: полиподиальные лобозные, моноподиальные лобозные, филозные, конические, гранулоретикулозные, конические актиноподии, неконические актиноподии.
 Лобоподии и филоподии Лобоподии – широкие, с закругленным концом выпячивания клеточной мембраны. На конце область прозрачной эктоплазмы, за которой находится менее прозрачная эндоплазма с клеточными органеллами. Филоподии – очень гибкие, тонкие (около 0, 2 мкм в диаметре) псевдоподии, под клеточной мембраной – ось из микрофиламентов. Они очень подвижны; быстро выпускаются клеткой и так же быстро втягиваются обратно. Возможно, действуют как чувствительные щупальца, с помощью которых клетка исследует консистенцию находящейся перед ней поверхности, прежде чем наползет на нее
Лобоподии и филоподии Лобоподии – широкие, с закругленным концом выпячивания клеточной мембраны. На конце область прозрачной эктоплазмы, за которой находится менее прозрачная эндоплазма с клеточными органеллами. Филоподии – очень гибкие, тонкие (около 0, 2 мкм в диаметре) псевдоподии, под клеточной мембраной – ось из микрофиламентов. Они очень подвижны; быстро выпускаются клеткой и так же быстро втягиваются обратно. Возможно, действуют как чувствительные щупальца, с помощью которых клетка исследует консистенцию находящейся перед ней поверхности, прежде чем наползет на нее
 Ретикулоподии Называют также ризоподии или сетчатые псевдоподии, чаще встречаются у фораменифер. Они тонкие, сильно разветвляющиеся нитевидные, образуют между собой множество анастомозов, что создает сложную трехмерную сеть вокруг клетки. Внутри ризоподии находится ось из микрофиламентов, в окружающей эту ось цитоплазме часто обнаруживается много гранул (митохондрии и цитоплазматические включения). Вся сеть может сокращаться, но редко используется клеткой для передвижения.
Ретикулоподии Называют также ризоподии или сетчатые псевдоподии, чаще встречаются у фораменифер. Они тонкие, сильно разветвляющиеся нитевидные, образуют между собой множество анастомозов, что создает сложную трехмерную сеть вокруг клетки. Внутри ризоподии находится ось из микрофиламентов, в окружающей эту ось цитоплазме часто обнаруживается много гранул (митохондрии и цитоплазматические включения). Вся сеть может сокращаться, но редко используется клеткой для передвижения.
 Аксоподии Характерен двунаправленный ток цитоплазмы. Есть осевая опора из микротрубочек. Используются для движения и питания.
Аксоподии Характерен двунаправленный ток цитоплазмы. Есть осевая опора из микротрубочек. Используются для движения и питания.

 Строение клетки амебоидного организма
Строение клетки амебоидного организма
 Амебоидное движение Псевдоподия формируется на переднем конце амебы. Суть этого процесса в изменении вязкости цитоплазмы. Внутренняя текучая цитоплазма называется эндоплазмой и находится в состоянии золя. Эндоплазма течет вперед, заполняя псевдоподию. Спереди эндоплазма превращается в похожую на плотный гель эктоплазму, которая располагается непосредственно под цитоплазматической мембраной. Сзади амебы эктоплазма переходит обратно в эндоплазму и течет к переднему концу клетки, где вновь превращается в эктоплазму. Этот циклический процесс продолжается в течение всего движения амебы.
Амебоидное движение Псевдоподия формируется на переднем конце амебы. Суть этого процесса в изменении вязкости цитоплазмы. Внутренняя текучая цитоплазма называется эндоплазмой и находится в состоянии золя. Эндоплазма течет вперед, заполняя псевдоподию. Спереди эндоплазма превращается в похожую на плотный гель эктоплазму, которая располагается непосредственно под цитоплазматической мембраной. Сзади амебы эктоплазма переходит обратно в эндоплазму и течет к переднему концу клетки, где вновь превращается в эктоплазму. Этот циклический процесс продолжается в течение всего движения амебы.
 Механизм формирования псевдоподии Разрушение актиновых филаментов увеличивает локальную осмотическую концентрацию. Жидкость изнутри клетки движется вдоль градиента концентрации, образуя псевдоподию. Актин реполимеризуется и стабилизирует эту часть клетки, формируя систему филаментов.
Механизм формирования псевдоподии Разрушение актиновых филаментов увеличивает локальную осмотическую концентрацию. Жидкость изнутри клетки движется вдоль градиента концентрации, образуя псевдоподию. Актин реполимеризуется и стабилизирует эту часть клетки, формируя систему филаментов.
 Способы передвижения амеб с использованием различных типов псевдоподий Ползание Качение Перетекание с лобоподиями Ходьба Ползание с филоподиями Бипедальное шагание
Способы передвижения амеб с использованием различных типов псевдоподий Ползание Качение Перетекание с лобоподиями Ходьба Ползание с филоподиями Бипедальное шагание
 Перемещение клетки с помощью филоподий a | Движение начинается с образования актин зависимых выростов по переднему краю, который состоит из ламеллиподии и филоподий. Эти выросты содержат актиновые филаменты с концами, ориентированными по направлению к мембране. b | В течение клеточного вытягивания образуются новые точки приклеивания к субстрату по переднему краю. c | Затем ядро и тело клетки перемещаются вперед силой актин-миозиновых контактов. d | Сокращение волокон толкает клетку вперед, что сопровождается откреплением ее от участков сзади.
Перемещение клетки с помощью филоподий a | Движение начинается с образования актин зависимых выростов по переднему краю, который состоит из ламеллиподии и филоподий. Эти выросты содержат актиновые филаменты с концами, ориентированными по направлению к мембране. b | В течение клеточного вытягивания образуются новые точки приклеивания к субстрату по переднему краю. c | Затем ядро и тело клетки перемещаются вперед силой актин-миозиновых контактов. d | Сокращение волокон толкает клетку вперед, что сопровождается откреплением ее от участков сзади.
 Формирование и движение филоподий По: Filopodia: Steps in filopodia formation and movement. Manual of Cellular and Molecular Function, The National University of Singapore, Republic of Singapore. 2009 Стадии формирования филоподий. Динамическое поведение филоподий.
Формирование и движение филоподий По: Filopodia: Steps in filopodia formation and movement. Manual of Cellular and Molecular Function, The National University of Singapore, Republic of Singapore. 2009 Стадии формирования филоподий. Динамическое поведение филоподий.
 Полимеризация актина в") Латеральные движения филоподий Направление движения перемещающейся клетки контролируется актиновыми комплексами. (1) Полимеризация актина в основании филоподии (желтая стрелка толкает мембрану вперед и влево. (2 -3) Силы полимеризации актина в филоподиях приводят к латеральному движению филоподии. (4 -6) Филоподии могут пресекаться, терять самостоятельность и образовывать трехмерную сеть. (7 -8) Когда латерально двигающаяся филоподия встречает другую филоподию они могут сливаться, объединяясь в одну. При этом образуется филоподия гораздо более широкая и длинная.
Латеральные движения филоподий Направление движения перемещающейся клетки контролируется актиновыми комплексами. (1) Полимеризация актина в основании филоподии (желтая стрелка толкает мембрану вперед и влево. (2 -3) Силы полимеризации актина в филоподиях приводят к латеральному движению филоподии. (4 -6) Филоподии могут пресекаться, терять самостоятельность и образовывать трехмерную сеть. (7 -8) Когда латерально двигающаяся филоподия встречает другую филоподию они могут сливаться, объединяясь в одну. При этом образуется филоподия гораздо более широкая и длинная.
 Строение жгутика
Строение жгутика
 Строение жгутика
Строение жгутика
 Движение жгутиков, токи воды и направление движения клеток различных протистов
Движение жгутиков, токи воды и направление движения клеток различных протистов
 Модель поступательного движения T. brucei в водной среде Rodríguez J A et al. PNAS 2009; 106: 19322 -19327
Модель поступательного движения T. brucei в водной среде Rodríguez J A et al. PNAS 2009; 106: 19322 -19327
 Механизм работы реснички
Механизм работы реснички
 Метаболия или эвгленоидное движение A. Три различные стадии похожего на перистальтику движения неокрашенной первично осмотрофной эвглениды Astasia klebsii. B–D. SEM, метаболия различных эвгленид: (B) Distigma, (C) Eutreptia и (D) Euglena. © 2004 A: W. Bourland; B-D: B. Leander. Способность к метаболии увеличивается при возрастании числа пластинок пелликулы до 20 и более. Появляется возможность заглатывания крупных пищевых частиц, в т. ч. эукариотических клеток
Метаболия или эвгленоидное движение A. Три различные стадии похожего на перистальтику движения неокрашенной первично осмотрофной эвглениды Astasia klebsii. B–D. SEM, метаболия различных эвгленид: (B) Distigma, (C) Eutreptia и (D) Euglena. © 2004 A: W. Bourland; B-D: B. Leander. Способность к метаболии увеличивается при возрастании числа пластинок пелликулы до 20 и более. Появляется возможность заглатывания крупных пищевых частиц, в т. ч. эукариотических клеток
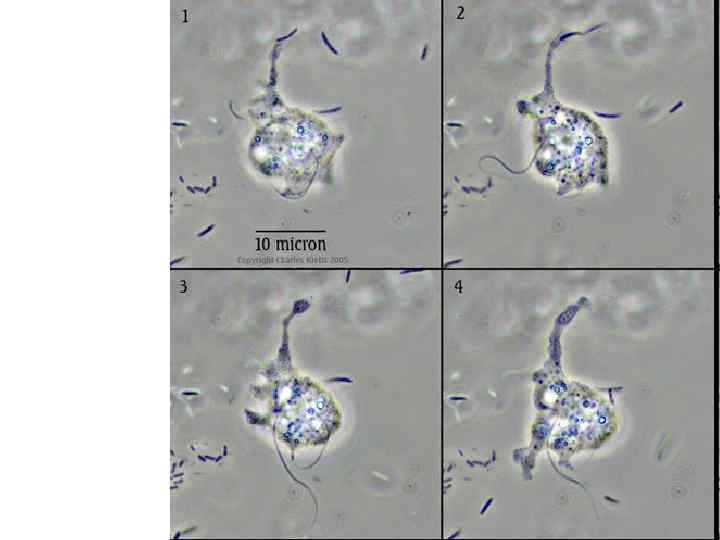
 Типы деления клеток простейших В основе процессов размножения лежит деление клетки: деление ядра, затем деление цитоплазмы. Выделяют три основных типа деления клеток: монотомия, палинтомия и синтомия. Монотомия (простое деление надвое) – вслед за делением ядра происходит деление цитоплазмы и далее следует период роста. При неравномерном делении материнской клетки носит название почкования.
Типы деления клеток простейших В основе процессов размножения лежит деление клетки: деление ядра, затем деление цитоплазмы. Выделяют три основных типа деления клеток: монотомия, палинтомия и синтомия. Монотомия (простое деление надвое) – вслед за делением ядра происходит деление цитоплазмы и далее следует период роста. При неравномерном делении материнской клетки носит название почкования.
 – серия быстрых делений без периодов") Типы деления клеток простейших Палинтомия (множественное последовательное деление) – серия быстрых делений без периодов роста между ними при этом у разных видов образуется от 4 до 256 (или больше) дочерних клеток. Синтомия (шизогония) – деления ядер без деления цитоплазмы, затем деление цитоплазмы и распад материнской клетки
Типы деления клеток простейших Палинтомия (множественное последовательное деление) – серия быстрых делений без периодов роста между ними при этом у разных видов образуется от 4 до 256 (или больше) дочерних клеток. Синтомия (шизогония) – деления ядер без деления цитоплазмы, затем деление цитоплазмы и распад материнской клетки
 Amoeba leningradensis (Amoebidae), (B)") Представители главных филогенетических линий голых амеб A D. Tubulinea. (A) Amoeba leningradensis (Amoebidae), (B) Saccamoeba limax (Hartmannellidae) (C, D) две альтернативные формы Rhizamoeba saxonica (Leptomyxidae). E, F. Flabellinea, Vannellida. (E) Vannella simplex, (F) Vannella devonica. G J. Flabellinea, Dactylopodida. (G) Neoparamoeba pemaquidensis (H) Korotnevella stella, (I) Vexillifera bacillipedes (J) Vexillifera minutissima. Дактилоподии отмечены стрелками. По статье: Smirnov Масштаб: 25 мкм. A. et al. 2005. Molecular Phylogeny and Classification of the Lobose Amoebae // Protist, Vol. 156, P. 129 -142
Представители главных филогенетических линий голых амеб A D. Tubulinea. (A) Amoeba leningradensis (Amoebidae), (B) Saccamoeba limax (Hartmannellidae) (C, D) две альтернативные формы Rhizamoeba saxonica (Leptomyxidae). E, F. Flabellinea, Vannellida. (E) Vannella simplex, (F) Vannella devonica. G J. Flabellinea, Dactylopodida. (G) Neoparamoeba pemaquidensis (H) Korotnevella stella, (I) Vexillifera bacillipedes (J) Vexillifera minutissima. Дактилоподии отмечены стрелками. По статье: Smirnov Масштаб: 25 мкм. A. et al. 2005. Molecular Phylogeny and Classification of the Lobose Amoebae // Protist, Vol. 156, P. 129 -142
, питающихся клетками") Трофозоиты Entamoeba histolytica На мазках показаны несколько трофозоидов дизентерийной амебы (тканевая форма), питающихся клетками крови человека
Трофозоиты Entamoeba histolytica На мазках показаны несколько трофозоидов дизентерийной амебы (тканевая форма), питающихся клетками крови человека
 Циста E. histolytica (4 ядра) Циста E. coli") Жизненный цикл дизентерийной амебы (Entamoeba histolytica) Циста E. histolytica (4 ядра) Циста E. coli (8 ядер)
Жизненный цикл дизентерийной амебы (Entamoeba histolytica) Циста E. histolytica (4 ядра) Циста E. coli (8 ядер)
 Стадии развития Naegleria fowleri циста трофозоит жгутиковая форма N. fowleri – свободно живущая амёба, интенсивно размножается в тёплых (25– 30 °С) проточных и непроточных пресных водоёмах (реках, озерах), в геотермальных источниках, промышленных стоках, почве, а также в плохо очищаемых бассейнах. Попадает в организм человека во время купания. Может вторгаться и разрушать нервную систему человека, что почти всегда заканчивается смертью зараженного через 2– 7 дней после первых симптомов болезни (инкубационный период 3– 7 дней). Первые симптомы заболевания – головная боль, изменение вкусовых и обонятельных ощущений. Одна из мер профилактики - избегать купания в водоемах в жаркую погоду. N. fowleri была открыта в Австралии в 1960 -е гг. Она теплолюбива, причина роста заболеваемости – необычно жаркая погода летом, а также общая тенденция глобального потепления.
Стадии развития Naegleria fowleri циста трофозоит жгутиковая форма N. fowleri – свободно живущая амёба, интенсивно размножается в тёплых (25– 30 °С) проточных и непроточных пресных водоёмах (реках, озерах), в геотермальных источниках, промышленных стоках, почве, а также в плохо очищаемых бассейнах. Попадает в организм человека во время купания. Может вторгаться и разрушать нервную систему человека, что почти всегда заканчивается смертью зараженного через 2– 7 дней после первых симптомов болезни (инкубационный период 3– 7 дней). Первые симптомы заболевания – головная боль, изменение вкусовых и обонятельных ощущений. Одна из мер профилактики - избегать купания в водоемах в жаркую погоду. N. fowleri была открыта в Австралии в 1960 -е гг. Она теплолюбива, причина роста заболеваемости – необычно жаркая погода летом, а также общая тенденция глобального потепления.
 Жизненный цикл Naegleria fowleri имеет три стадии жизненного цикла: циста, трофозоит и жгутиковая форма. Трофозоиты делятся промитозом (ядерная мембрана остается нетронутой. Трофозоиты могут превращаться во временно не питающуюся жгутиковую форму, которая обычно возвращается на стадию трофозоита. Трофозоиты инфицируют человека и животных проникая через слизистую носа и мигрируют в мозг по обонятельным нервам, вызывают первичный амебный менингоэнцефалит. N. fowleri обнаруживается в цереброспинальной жидкости (жгутиковая форма) и тканях (трофозоиты). Цисты не обнаруживаются в тканях мозга.
Жизненный цикл Naegleria fowleri имеет три стадии жизненного цикла: циста, трофозоит и жгутиковая форма. Трофозоиты делятся промитозом (ядерная мембрана остается нетронутой. Трофозоиты могут превращаться во временно не питающуюся жгутиковую форму, которая обычно возвращается на стадию трофозоита. Трофозоиты инфицируют человека и животных проникая через слизистую носа и мигрируют в мозг по обонятельным нервам, вызывают первичный амебный менингоэнцефалит. N. fowleri обнаруживается в цереброспинальной жидкости (жгутиковая форма) и тканях (трофозоиты). Цисты не обнаруживаются в тканях мозга.
 Патология при инфицировании мозга неглерией
Патология при инфицировании мозга неглерией
 Строение раковин фораминифер Ископаемые фузулиниды в известняке Nonionina depressula
Строение раковин фораминифер Ископаемые фузулиниды в известняке Nonionina depressula
 Гигантская фораминифера Gromia sphaerica Диаметр клетки составляет до 3 см. Двигаясь по дну она оставляет билатерально симметричный след. Такие же следы находили в докембрии в породах возрастом 1. 8 млрд. лет. Слева отмытая от наилка особь. Справа вверху следы, оставляемые громией при ползании. Внизу особь на дне, покрытая наилком.
Гигантская фораминифера Gromia sphaerica Диаметр клетки составляет до 3 см. Двигаясь по дну она оставляет билатерально симметричный след. Такие же следы находили в докембрии в породах возрастом 1. 8 млрд. лет. Слева отмытая от наилка особь. Справа вверху следы, оставляемые громией при ползании. Внизу особь на дне, покрытая наилком.
 Планктонные и бентосные фораминиферы По: Brasier, 1980
Планктонные и бентосные фораминиферы По: Brasier, 1980
 Строение фораминифер
Строение фораминифер
 Структура и состав стенки раковины фораминифер Хитинизированная стенка: представлена у пресноводных бентосных фораменифер. Агглютинированная стенка: специфична для бентосных фораминифер, строящих раковинки, цементирую экзогенные частицы (песчинки, оолиты, мелкие частицы кальцита или спикул губок) при помощи карбонатной минерализации. Стенка состоит из тонкого слоя частиц по периметру клетки. Карбонатная стенка (Ca. CO 3): присутствует как у бентосных, так и у планктонных фораменифер. Стекловидная стенка: прозрачная, перфорированная, кристаллы радиальные, с пластинками, разделенынми органическими слоями. Фарфоровая стенка: блестящая, гладкая, кристаллы расположены случайно
Структура и состав стенки раковины фораминифер Хитинизированная стенка: представлена у пресноводных бентосных фораменифер. Агглютинированная стенка: специфична для бентосных фораминифер, строящих раковинки, цементирую экзогенные частицы (песчинки, оолиты, мелкие частицы кальцита или спикул губок) при помощи карбонатной минерализации. Стенка состоит из тонкого слоя частиц по периметру клетки. Карбонатная стенка (Ca. CO 3): присутствует как у бентосных, так и у планктонных фораменифер. Стекловидная стенка: прозрачная, перфорированная, кристаллы радиальные, с пластинками, разделенынми органическими слоями. Фарфоровая стенка: блестящая, гладкая, кристаллы расположены случайно
. Микросферическая форма") Жизненный цикл фораминифер Фораминиферы имеют две стадии развития: 1. Бесполое размножение (шизогония). Микросферическая форма с большим количеством камер. - производит молодого гамотта с гаплоидным набором хромосом (n) - встречается в зимние месяцы 2. Половое размножение (гамогония). Мегалосферическая форма - производит молодого шизонта с диплоидным набором хромосом (2 n) летние месяцы У планктонных фораминифер обнаружено только половое размножение - встречается в
Жизненный цикл фораминифер Фораминиферы имеют две стадии развития: 1. Бесполое размножение (шизогония). Микросферическая форма с большим количеством камер. - производит молодого гамотта с гаплоидным набором хромосом (n) - встречается в зимние месяцы 2. Половое размножение (гамогония). Мегалосферическая форма - производит молодого шизонта с диплоидным набором хромосом (2 n) летние месяцы У планктонных фораминифер обнаружено только половое размножение - встречается в
 Живые радиолярии (20 X)") Радиолярии Скелет ископаемой радиолярии (250 X) Живые радиолярии (20 X)
Радиолярии Скелет ископаемой радиолярии (250 X) Живые радиолярии (20 X)
 Raphidiophrys elegans
Raphidiophrys elegans